
Khaled Arafat
Department of Plant Pathology, Faculty of Agriculture, New Valley University, Egypt
LiveDNA: 20.7732
Asian Journal of Plant Pathology
Year: 2018 | Volume: 12 | Issue: 1 | Page No.: 27-37







ABSTRACT
Background and Objective: Guava black spot (GBS) disease is a quiescent infection, that infect immature fruit prior to harvest. Visible symptoms of the disease on guava fruit showed sunken lesions with concentric development, variation in color ranging from greenish black to black and spread in severity affected fruit. An unrecorded disease of guava fruit (Psidium guajava L.) cv. White Balady, was observed in this study during postharvest disease survey in Egypt. Materials and Methods: Tissues of guava fruit spot used to isolate the pathogenic fungal. To perform the phylogenetic analysis, the internal transcribed spacer (ITS) region amplified by Polymerase chain reaction (PCR). To amplify the ITS, the primer ITS-1 and reverse primer ITS-4 used to amplify rDNA-ITS regions of the fungus. The fungal identification was done by molecular analysis as Phyllosticta capitalensis novel isolate ARAFAT-GF5 according to the GenBank (Accession number–LC269950.1; GI: 119461242) with the synonym: Guignardia mangiferae. Results: The isolate ARAFAT-GF5 (626 bootstrap) used and the Basic Local Alignment Search Tool (BLAST) program used to search for nucleotide sequence homology in GenBank. The computational analysis of the synonymous DNA sequence was useful for predicting the codon profiling. Pathogenicity test performed to complete Koch’s postulates. Typical black spot symptoms developed and the pathogen recovered from the inoculated fruit after 10 days and found as P. capitalensis. Conclusion: This is the first report of black spot disease on guava fruits in Egypt, caused by a novel isolate of P. capitalensis. The results presented here may enable enhancements in the program of integrated disease management.
PDF Abstract XML References Citation
Received: September 24, 2018;
Accepted: November 07, 2018;
Published: December 01, 2018
How to cite this article
Khaled Arafat, 2018. A Novel Isolate of Phyllosticta capitalensis Causes Black Spot Disease on Guava Fruit in Egypt. Asian Journal of Plant Pathology, 12: 27-37.
URL: https://scialert.net/abstract/?doi=ajppaj.2018.27.37
URL: https://scialert.net/abstract/?doi=ajppaj.2018.27.37
INTRODUCTION
Guava fruit (Psidium guajava L.) cultivated and widespread in many tropical and subtropical regions. Egypt is a subtropical country which existing between 22° and 32° north latitude. In recent years, several studies have focused on the incidence of postharvest diseases that decrease the value of the fruits and alter their physical and chemical properties and contribute to their reduced shelf life1-3. The major postharvest diseases of guava are including anthracnose, black spot and astylar end rot3,4. The GBS disease is a quiescent infection, that infect immature fruit prior to harvest. Fruits with quiescent infections stay asymptomatic until maturity, when structural and physiological changes trigger the onset of the disease5. However, there have been few studies in the literature reporting GBS disease. The G. psidii reported in the first study, which causing black spot on guava fruits, during in the field and transportation in India6. While G. pisidii infection occurs in young fruits and stays quiescent until maturity in Brazil7. Similarly, P. psidiicola, reported as a potentialcauseof GBS disease in Taiwan8 and in Venezuela9. The results to date, has been extensive experiments on Guignrdia species, where 10 isolated from asymptomatic tissues on different hosts, that were G. mangiferae classification by rDNA ITS1-5.8S-ITS2 sequence methods, caused symptoms in guava fruits10. Visible symptoms of the disease showed sunken lesions with concentric development, variation in color ranging from greenish black to black and spread on severity affected fruit9. The genus Phyllosticta Pers. Ex Desm. confirmed11. It includes endophytes, plant pathogens and saprobes12-17. Species in the genus Phyllosticta are mostly plant pathogens of a wide range of hosts and responsible for diseases, including black spots on leaf and fruits13,18-24. Phyllosticta species are also potential biocontrol agents25 and has reported to produce novel mycotoxin viz. phyllostine and phyllostoxin26. Recently, the name Phyllosticta Pers. Ex. Desm. (asexual state) and Guignardia Viala and Ravaz (sexual state) have used separately following the dual classification system used by mycologist over several decades27-30. ITS rDNA sequences often used to infer phylogeny relationships in many groups of fungi, including Phyllosticta10,14,15,31. Nevertheless, researchers using molecular methods suggested that the fungi isolates found as G. psidii could be in fact G. mangiferae or also could be conspecific to this cosmopolitan species32. The objective of this study was to identify and characterization of P. capitalensis novel isolate ARAFAT-GF5 associated with GBS a new disease on guava fruit in Egypt. Identification of the isolate performed using DNA sequence data of the rDNA ITS1-5.8S-ITS2-28S.
MATERIALS AND METHODS
Samples collection and isolation of the pathogenic fungal: P. capitalensis obtained from naturally guava fruit (Psidium guajava L.) cv. White Balady at an immature and mature stage. Fruits collected for each three seasons (2015-2017) from different local markets in El-Kharga city (25.4390 N, 30.5586 E), New Valley Governorate, Egypt. The collected samples kept in sterilized polyethylene bags and brought to the laboratory of the Plant Pathology Department, Faculty of Agriculture, New Valley University, Egypt. Fruit samples cut into (5 mm) and immersed in NaOCl (0.5%) for 5 min rinsed in sterile distilled water, then transfer to blotted dry in sterile paper towels for drying. Samples transferred into 9 cm Petri dishes containing potato dextrose agar (PDA) and incubation at 25±3°C under a 12 h photo period for 10-15 days33. The fungus characterized by initially gray and turned black with overripe.
Identification of pathogenic fungus: Identification of pathogenic fungus based on morphological methods, using characters of the phenotype of the fungus culture, i.e., colony or hyphae, the characters of the spore or reproductive structure if these features were discernible34, 35.
Molecular characterization of pathogenic fungus: The fungus was grown in the cultivation media and incubated at 25°C for 15 days, then the growth of fungal was scraped and suspended in 100 μL of distilled water and boiled at 100°C for 15 min and stored at -80°C. DNA was extracted from fungal cultures using the genomic DNA Prep kit (SolGent, Daejeon, Korea) according to SDS/CTAB lysis and phenol/chloroform extraction method15. The ITS region, including ITS1, 5.8S and ITS4, 28S rRNA amplified via PCR using primer pair ITS1 (5'–CTTGGTCATTTAGAGGAAGTAA-3') and ITS4 (5' TCCTCCGCTTATTGATATGC-3')36. The ITS sequence obtained through the commercial service offered by Macrogen (Macrogen Comp., South Korea). The sequence compared with known homologous sequences of Phyllosticta and Guignardia in databanks (National Center for Biotechnology Information (NCBI)-(http: //www.ncbi.nlm.nih.gov/Genbank) and The European Molecular Biology Laboratory (EMBL) (https://www.ebi.ac.uk/) by the BLSAT program. DNA sequences deposited in the GenBank database (GenBank Accession No. LC269950).
Sequence analysis: The BLAST search program at NCBI (http://blast.ncbi.nlm.nih.gov/BLAST.cgi) used to analyze the obtained sequence. CLUSTALW program (http://clustalw.ddbj.nig.ac.jp/top-ehtml) applied to achieve the sequence alignment and phylogeny. Phylogenetic analysis performed by a neighbor joining method to infer the relationships between the fungus isolate and sequences available for Phyllosticta and Guignardia in the NCBI and The EMBL nucleotide databases using Kimura 2-parameter distances37. For analysis, 100 and 33 bootstrap replicates performed to assess the statistical support for the tree.
Nucleotide distributions and frequencies: The DNA sequence analysis used free software (http://www. bioinformatics.org/sms2/index.html) usefully to attain the coding usage of DNA stats. DNA stats returns the number of occurrences of each residue in the sequence entered. Codon usage accepts a DNA sequence and returns the number and frequency of each codon type. Since the program also compares the frequencies of codons that code for the same amino acid (synonymous codons). So, it used to assess whether a sequence shows a preference for certain synonymous codons38.
Pathogenicity test: Guava fruits obtained from a local supermarket and immediately transferred into mycological laboratory. Fruits in similar shape and size certain and treated with 96% ethanol, soaked with sterilized distilled water and drained at room temperature 25+2°C. Two wounds (5 mm diameter and 3 mm deep) made through at different equatorial lines of each fruit using the tip of a sterile cork-borer. Each one of guava fruits inoculated with a mycelial plug (5 mm in diameter) of the fungus culture into each wound. Other guava fruits having artificial wounds of only plugs of PDA culture used as control. Twenty-five fruits used for each treatment and then the fruits air dried and placed in the plastic boxes (with wetted sterilized cotton pieces to maintain high-level of humidity). The experiment frequent twice. The virulence of the tested fungus identified by observing the development of GBS disease, after 10 days on infested guava fruits39.
RESULTS
Samples collection and isolation of the pathogenic fungal: Samples of guava fruit at immature and mature stage, collected according to GBS disease symptoms, from the different local market in El-Kharga city, New Valley Governorate, Egypt. The first visible symptoms of the infected guava fruit were small, slightly sunken on mature fruits. Symptom developer showed in Fig. 1, sunken lesions with concentric development, variation in color ranging from greenish black to black and spread in severity affected fruit. As shown before, these symptoms showed that, a strong relationship between symptoms and P. capitalensis as the pathogenic fungal of GBS disease of guava fruit.
Identification of the pathogenic fungus: Morphological characters of the isolated fungus from guava fruit showed that closed to P. capitalensis in colony appearance, although the hyphal growth of PDA culture, produced gray mycelium at the early stage of growth followed by black colored conidia (Fig. 2). Conidia is hyaline, unicellular, obovate, ranged 6-11 X 5-7 μm (Fig. 3). The pathogen found associated with the GBS disease based on morphological features identified as Phyllosticta capitalensis.
Molecular characterization of pathogenic fungus: The fungus found by molecular analysis as P. capitalensis according to the GenBank (Accession number-LC269950.1; GI: 119461242) with synonym: Guignardia mangiferae. Analysis of ITS1, 5.8S rRNA, ITS2, 28S rRNA, partial and complete sequence. The DNA from P. capitalensis from guava fruit amplified when the PCR region performed using primers ITS1 and ITS4. The corresponding PCR region amplified the ITS rDNA sequence of region 1 and 4, which also include 5.8S rRNA and 28S rRNA gene. The PCR produce was 626 bp.
Sequence analysis: ITS sequence of P. capitalensis isolate (ARAFAT-GF5) aligned with different Phyllosticta isolates available from the GenBank nucleotide database. The aligned sequences visually inspected and minor adjustments made to improve alignment. Phylogenetic analysis performed by a neighbor joining method to infer the relationships between the Phyllosticta isolate (ARAFAT-GF5) and sequences available for Phyllosticta and Guignardia in the NCBI (https://www.ncbi.nlm.nih.gov/nuccore/?cmd=historysearch&querykey=1) and EMBL-EBI (https://www.ebi.ac.uk/ena/ data/view/LC269950) nucleotide databases (Table 1) using Kimura 2-parameter distances. For analysis, 100 bootstrap replicates performed to assess the statistical support for the tree. Phyllosticta capitalensis (LC 269950.1) sequence No. 1 homologous with all sequences, ranged score between (98.08-100%). Furthermore, the isolate LC269950 compared with the other sequences (33 bootstrap) published for Phyllosticta and Guignardia isolates obtained in the NCBI. Alignment of the available sequences of Phyllosticta and Guignardia exposed both similarity in the ITS sequences (Table 2).
| Table 1: | Nucleotide length of 100 isolates compared with P. capitalensis and GenBank accession numbers of their characters data |
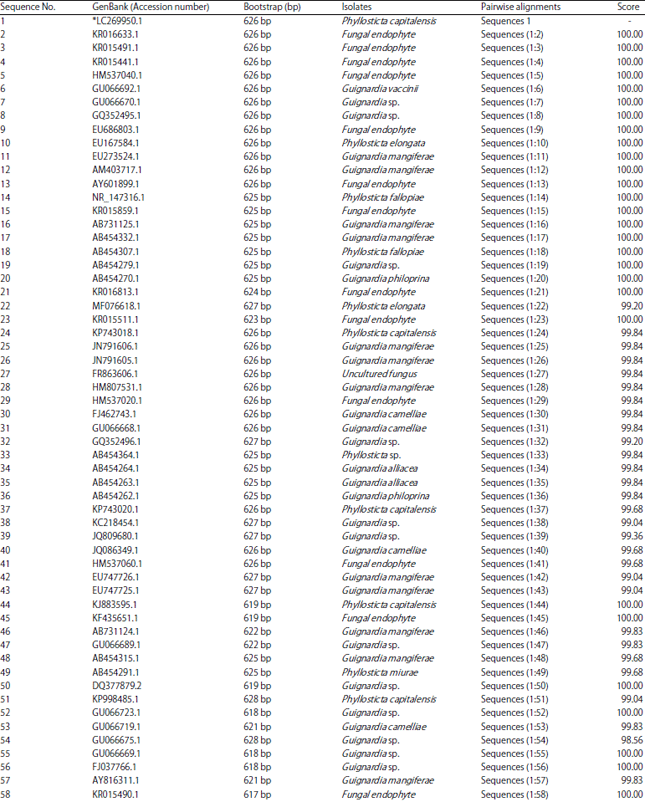 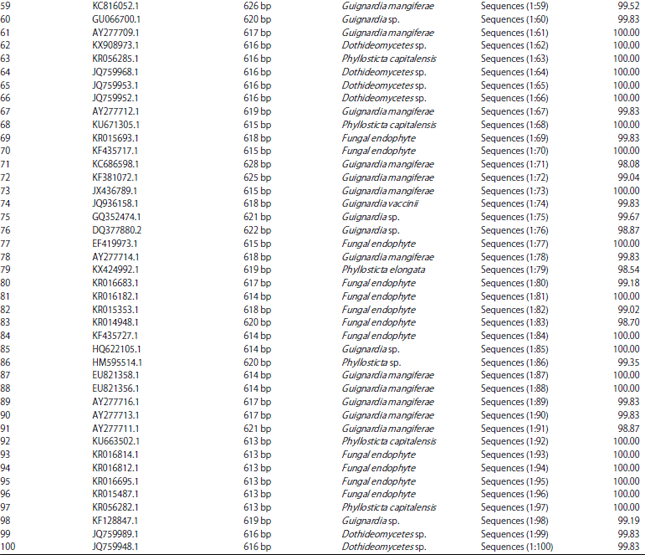 | |
 | |
| Fig. 1: | Natural GBS disease symptoms of guava fruits |
 | |
| Fig. 2: | Hyphal growth of P. capitalensis on PDA medium |
| Table 2: | Nucleotide length of 33 isolates of Phyllosticta or Guignardia compared with P. capitalensis and GenBank accession numbers of their characters data |
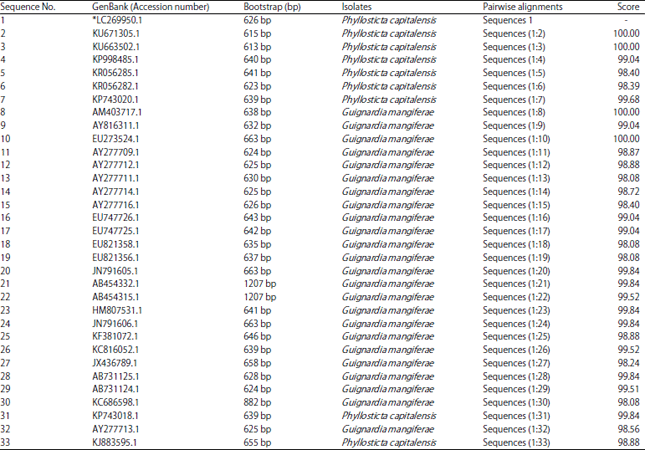 | |
 | |
| Fig. 3: | Conidia spores of P. capitalensis (X400) |
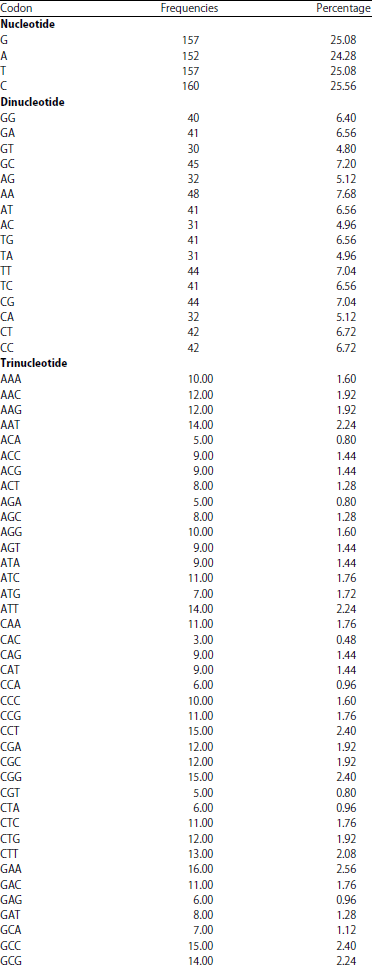
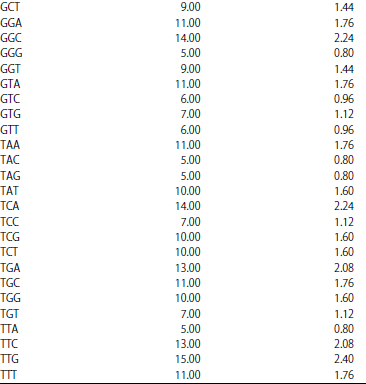
Nucleotide distributions and frequencies: The results of P. capitalensis (LC 269950.1) showed that sequences length, consisting of 626 bases. Distributions and frequencies of bases found (C) base repeated 160 times with 25.56%, followed by (G, T, A) bases repeated 157, 157, 152 times with 25.08, 25.08, 24.28%, respectively. Dinucleotide frequencies found the highest bases were (AA and GC) repeated 48 and 45 times with 7.68 and 7.20%, respectively. Moreover, G, C and A, T dinucleotide bases found repeated 317 and 309 times with 50.64 and 49.36%, respectively. Trinucleotide frequencies found the highest bases (GAA) repeated 16 times with 2.56% and the latest trinucleotide was (CAC) repeated 3 times with 0.48% (Table 3).
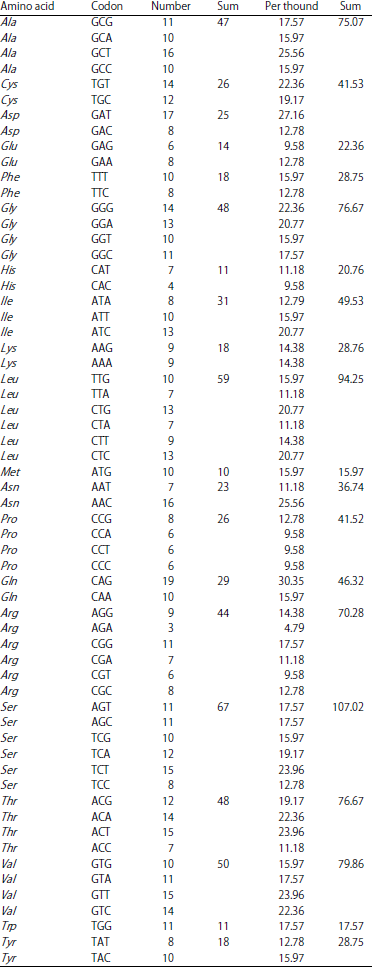
Codon usage: The analysis and simulations of P. capitalensis results for 626 sequences indicated that prediction of amino acids was the highest (leucine) frequency 26 times with 124.99 times/1000, followed by (serine, arginine, alanine, glycine, cysteine, isoleucine, asparagine, Valine, phenylalanine, glutamine, throine, proline, Tyrosine, glutamic acid, lysine, aspartic acid, tryptophane, histidine and methionine) frequencies “26, 18, 16, 15, 14, 11, 11, 11, 11, 9, 9, 9, 8, 7, 6, 6, 5, 5, 4 and 0" with “107.02, 76.93, 72.12, 67.32, 52.88, 52.88, 52.89, 79.86, 43.27, 28.85, 76.67, 38.48, 28.75, 28.85, 28..84, 24.04, 17.57 and 19.23/1000", respectively (Table 4).
| Table 3: | Frequencies and percentage nucleotides of P. capitalensis |
  | |
 | |
| Fig. 4: | Artificial GBS disease symptoms of guava fruit inoculated with P. capitalensis |
Taxonomy
Phyllosticta capitalensis (Guignardia mangiferae): Eukaryota, Fungi, Dikarya, Ascomycota, Pezizomycotina, Dothideomycetes, Dothideomycetes incertae sedis, Botryosphaeriales, Phyllostictaceae, Phyllosticta.
Pathogenicity tests: Pathogenicity of the characteristic P. capitalensis novel isolate ARAFAT-FG5 confirmed by inoculating guava fruits at mature stage. The artificially inoculated guava fruits developed black spot symptoms after 10 days of the inoculation. These symptoms similar to those of the naturally infected guava fruits (Fig. 4).
| Table 4: | Frequencies amino acids probability of P. capitalensis |
 | |
All control fruits persisted healthy. The pathogen of the inoculated fruit was re-isolated, cultivated and confirmed as ARAFAT-FG5 isolate based on fungal morphology. Pathogenicity tests revealed the presence of P. capitalensis as the pathogen for GBS in El-Kharga city, New Valley Governorate, Egypt. The morphological characteristic identification of the pathogen confirmed with a molecular and phylogenetic approach.
DISCUSSION
This was the first study of the GBS disease in Egypt, with P. capitalensis novel isolate ARAFAT-GF5 of guava fruit. The postharvest diseases caused by fungi handle biodeterioration of tropical fresh fruits pulp40,41. Postharvest fungal pathogens cause severe losses on guava during postharvest storage and marketing. The most aggressive pathogen is P. capitalensis on guava fruits under environmental conditions in El-Kharga city, New Valley Governorate-Egypt. The presence GBS disease had received little attention and not well documented in Egypt, hence, this study focused more attention to this disease. During investigation of postharvest fungal diseases, GBS disease of novel symptoms seen, comprehension of disease symptoms on plant hosts is important for field identification by taxonomists as well as a plant pathologist interested in disease incidence, management and distribution42. After infection by P. capitalensis the guava mature fruit may become sunken lesions with concentric development, variation in color ranging from greenish black to black and spread on severity affected fruit and pycnidia on fruits is usually black. The fungus isolated and found using a combination of morphological and molecular (ITS region sequences) methods. The morphological characteristics of the fungus P. capitalensis isolated from guava fruit, helped to show the fungus on PDA medium16,42,43. In the recent decade, results of molecular biology have progressed the systematic classifications of different multiplex groups of plant pathogenic fungi, including Phyllosticta species that have helped to facilitate the identification of species and resolution of species complex’s44-46. The ITS phylogram supported the identify of P. capitalensis (or Guignardia mangiferae) as a common foliar endophyte and pathogen with wide range of hosts15,47. Most of endophytic fungi belong to the ascomycetes and asexual fungi48. P. capitalensis was recorded 1543 times in GenBank to 31August2018 (https://www.ncbi.nlm.nih.gov/ Taxonomy/Browser/wwwtax.cgi?id=121624). However, there are powerful proofs that G. psidii and G. mangiferae are either the same species10. Moreover, no data available about the biology and ecology of the G. endophytes except for the production of chemically novel and pharmaceutically useful secondary metabolism of some isolates in Brazil49. Codon usage plays a significant role in the efficiency of the gene expression system. Studies have shown that the presence of rare codons influences gene expression levels and the solubility and amount of the expressed protein50,51. Therefore, synonymous codons not only specify protein sequences and translation dynamics, but also help determine gene expression levels52. However, this review emphasizes the significant role of determining codon usage to gene expression levels.
CONCLUSION
The novel isolate of P. capitalensis which isolated from guava fruit, found caused GBS as a new disease in Egypt. This is the first report of P. capitalensis causing GBS in Egypt. The results presented here may enable enhancements in the program of integrated disease management.
SIGNIFICANCE STATEMENT
This study discover the Phyllosticta capitalensis that can be beneficial for identification the causal agent of guava black spot disease This study will help the researcher to uncover the critical areas of postharvest diseases that many researchers were not able to explore. Thus a new theory on fungal taxonomy may be arrived at.
REFERENCES
- Martins, M.C., L. Amorim, S.A. Lourenco, A.S.S. Gutierrez and H.S. Watanabe, 2007. Incidence of post harvest damages in guavas at the wholesale market of Sao Paulo and its relationship to pre harvest bagging. Rev. Brasil. Fruticult., 29: 245-248.
CrossRefDirect Link - Fischer, I.H., A.M.D. Almeida, M.C.D. Arruda, R.M.D.A. Bertani, M.J.D.M. Garcia and L. Amorim, 2011. Postharvest damages in guavas from the Midwest region of the state of Sao Paulo. Bragantia, 70: 570-576.
CrossRefDirect Link - Gonzalez, M.S. and A. Rondon, 2005. First report of Guignardia psidii, an ascigerous state of Phyllosticta psidiicola, causing fruit rot on guava in Venezuela. Plant Dis., 89: 773-773.
CrossRefDirect Link - Wickert, E., A. de Souza, R.M. Pereira, L.T. Kishi, E.G. de Macedo Lemos and A. de Goes, 2014. Molecular and pathogenic study of Guignardia spp. isolates associated to different hosts. Adv. Microbiol., Vol. 4.
CrossRefDirect Link - Baayen, R.P., P.J.M. Bonants, G. Verkley, G.C. Carroll and H.A. van Der Aa et al., 2002. Nonpathogenic isolates of the citrus black spot fungus, Guignardia citricarpa, identified as a cosmopolitan endophyte of woody plants, G. mangiferae (Phyllosticta capitalensis). Phytopathology, 92: 464-477.
CrossRefDirect Link - Okane, I., A. Nakagiri, T. Ito and S. Lumyong, 2003. Extensive host range of an endophytic fungus, Guignardia endophyllicola (anamorph: Phyllosticta capitalensis). Mycoscience, 44: 353-363.
CrossRefDirect Link - Wulandari, N.F., C. To-Anun, K.D. Hyde, L.M. Duong and J. De Gruyter et al., 2009. Phyllosticta citriasiana sp. nov., the cause of Citrus tan spot of Citrus maxima in Asia. Fungal Divers., 34: 23-39.
Direct Link - Glienke, C., O.L. Pereira, D. Stringari, J. Fabris and V. Kava-Cordeiro et al., 2011. Endophytic and pathogenic Phyllosticta species, with reference to those associated with Citrus black spot. Persoonia, 26: 47-56.
Direct Link - Paul, A.P. and M.D. Blackburn, 1986. Phyllosticta beaumarisii sp. nov.: A cause of leafspot on Muehlenbeckia adpressa. Aust. Plant Pathol., 15: 40-41.
CrossRefDirect Link - McManus, P.S., 1998. First report of early rot of cranberry caused by Phyllosticta vaccinii in Wisconsin. Plant Dis., 82: 350-350.
CrossRefDirect Link - Olatinwo, R.O., E.J. Hanson and A.M.C. Schilder, 2003. A first assessment of the cranberry fruit rot complex in Michigan. Plant Dis., 87: 550-556.
CrossRefDirect Link - Paul, I., A.S. van Jaarsveld, L. Korsten and V. Hattingh, 2005. The potential global geographical distribution of citrus black spot caused by Guignardia citricarpa (Kiely): Likelihood of disease establishment in the European Union. Crop Prot., 24: 297-308.
CrossRefDirect Link - Liu, K., X. Ding, B. Deng, W. Chen, 2009. Isolation and characterization of endophytic taxol-producing fungi from taxus chinensis. J. Ind. Microbiol. Biotechnol., Vol. 36.
CrossRefDirect Link - Wang, X., G. Chen, F. Huang, J. Zhang, K.D. Hyde and H. Li, 2012. Phyllosticta species associated with citrus diseases in China. Fungal Divers., 52: 209-224.
CrossRefDirect Link - Yan, X.N., R.A. Sikora and J.W. Zheng, 2011. Potential use of cucumber (Cucumis sativus L.) endophytic fungi as seed treatment agents against root-knot nematode Meloidogyne incognita. J. Zhejiang Univ. Sci. B, 12: 219-225.
CrossRefDirect Link - Evidente, A., A. Cimmino, A. Andolfi, M. Vurro, M.C. Zonno and A. Motta, 2008. Phyllostoxin and phyllostin, bioactive metabolites produced by Phyllosticta cirsii, a potential mycoherbicide for Cirsium arvense biocontrol. J. Agric. Food Chem., 56: 884-888.
CrossRefDirect Link - Hawksworth, D.L., 2004. Fungal diversity and its implications for genetic resource collections. Stud. Mycol., 50: 9-18.
Direct Link - Shenoy, B.D., R. Jeewon and K.D. Hyde, 2007. Impact of DNA sequence-data on the taxonomy of anamorphic fungi. Fungal Divers., 26: 1-54.
Direct Link - Shenoy, B.D., R. Jeewon, H. Wang, K. Amandeep and W.H. Ho et al., 2010. Sequence data reveals phylogenetic affinities of fungal anamorphs Bahusutrabeeja, Diplococcium, Natarajania, Paliphora, Polyschema, Rattania and Spadicoides. Fungal Divers., 44: 161-169.
CrossRefDirect Link - Motohashi, K., S. Inaba, K. Anzai, S. Takamatsu and C. Nakashima, 2009. Phylogenetic analyses of Japanese species of Phyllosticta sensu stricto. Mycoscience, 50: 291-302.
CrossRefDirect Link - Wickert, E., A.D. Goes, E.G.D.M. Lemos, A.D. Souza, E.L.D. Silveira, F.D. Pereira and D. Rinaldo, 2009. Phylogenetic relationships and diversity of Guignardia spp isolated from different hosts on ITS1-5, 8S-ITS2 region. Rev. Brasil. Fruticult., 31: 360-380.
CrossRefDirect Link - Escanferla, M.E., S.R.G. Moraes, R.B. Salaroli and N.S. Massola, Jr., 2009. Prepenetration stages of Guignardia psidii in guava: Effects of temperature, wetness duration and fruit age. J. Phytopathol., 157: 611-624.
CrossRefDirect Link - White, T.J., T.D. Bruns, S.B. Lee and J.W. Taylor, 1990. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In: PCR Protocols: A Guide to Methods and Applications, Innis, M.A., D.H. Gelfand, J.J. Sninsky and T.J. White (Eds.), Academic Press, San Diego, CA, USA, ISBN-13: 9780123721808, pp: 315-322.
CrossRefDirect Link - Kimura, M., 1980. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol., 16: 111-120.
CrossRefDirect Link - Stothard, P., 2000. The sequence manipulation suite: JavaScript programs for analyzing and formatting protein and DNA sequences. Biotechniques, 28: 1102-1104.
PubMedDirect Link - Mohamed, H. and A. Saad, 2009. The biocontrol of postharvest disease (Botryodiplodia theobromae) of guava (Psidium guajava L.) by the application of yeast strains. Post Harvest Biol. Technol., 53: 123-130.
CrossRefDirect Link - Kusumaningrum, D., S.H.Lee, W.H. Lee, C. Mo and B.K. Cho, 2015. A review of technologies to prolong the shelf life of fresh tropical fruits in Southeast Asia. J. Biosyst. Eng., 40: 345-358.
CrossRefDirect Link - Wikee, S., D. Udayanga, P.W. Crous, E. Chukeatirote and E.H. McKenzie et al., 2011. Phyllosticta: An overview of current status of species recognition. Fungal Divers., 51: 43-61.
CrossRefDirect Link - Cai, L., D. Udayanga, D.S. Manamgoda, S.S. Maharachchikumbura and E.H. McKenzie et al., 2011. The need to carry out re-inventory of plant pathogenic fungi. Trop. Plant Pathol., 36: 205-213.
CrossRefDirect Link - Udayanga, D., X. Liu, E.H. McKenzie, E. Chukeatirote, A.H. Bahkali and K.D. Hyde, 2011. The genus Phomopsis: Biology, applications, species concepts and names of common phytopathogens. Fungal Divers., Vol. 50.
CrossRefDirect Link - Zhou, N., Q. Chen, G. Carroll, N. Zhang, R.G. Shivas and L. Cai, 2015. Polyphasic characterization of four new plant pathogenic Phyllosticta species from China, Japan and the United States. Fungal Biol., 119: 433-446.
CrossRefDirect Link - Huang, Y., J. Wang, G. Li, Z. Zheng and W. Su, 2001. Antitumor and antifungal activities in endophytic fungi isolated from pharmaceutical plants Taxus mairei, Cephalataxus fortunei and Torreya grandis. FEMS Immunol. Med. Microbiol., 31: 163-167.
CrossRefPubMedDirect Link - Rodrigues‐Heerklotz, K.F., K. Drandarov, J. Heerklotz, M. Hesse and C. Werner, 2001. Guignardic acid, a novel type of secondary metabolite produced by the endophytic fungus Guignardia sp.: Isolation, structure elucidation and asymmetric synthesis. Helvetica Chimica Acta, 84: 3766-3772.
CrossRefDirect Link - Rosano, G.L. and E.A. Ceccarelli, 2009. Rare codon content affects the solubility of recombinant proteins in a codon bias-adjusted Escherichia coli strain. Microbial Cell Fact., Vol. 8.
CrossRefDirect Link - Josse, L., T. Singh and T. von der Haar, 2018. Experimental determination of codon usage-dependent selective pressure on high copy-number genes in Saccharomyces cerevisiae. BioRxiv.
CrossRefDirect Link - Daniel, E., G.U. Onwukwe, R.K. Wierenga, S.E. Quaggin, S.J. Vainio and M. Krause, 2015. ATGme: Open-source web application for rare codon identification and custom DNA sequence optimization. BMC Bioinfor., Vol. 16. 10.1186/s12859-015-0743-5.
Direct Link