
Siti Nur Aisyah
Laboratory of Biotechnology, Faculty of Agriculture, Andalas University, 2nd Floor, Campus Limau Manis, 25163 Padang, West Sumatera, Indonesia
Sulastri Sulastri
Laboratory of Biotechnology, Faculty of Agriculture, Andalas University, 2nd Floor, Campus Limau Manis, 25163 Padang, West Sumatera, Indonesia
Retmi Retmi
Laboratory of Biotechnology, Faculty of Agriculture, Andalas University, 2nd Floor, Campus Limau Manis, 25163 Padang, West Sumatera, Indonesia
Rahmi Henda Yani
Laboratory of Biotechnology, Faculty of Agriculture, Andalas University, 2nd Floor, Campus Limau Manis, 25163 Padang, West Sumatera, Indonesia
Elly Syafriani
Laboratory of Biotechnology, Faculty of Agriculture, Andalas University, 2nd Floor, Campus Limau Manis, 25163 Padang, West Sumatera, Indonesia
Lily Syukriani
Laboratory of Biotechnology, Faculty of Agriculture, Andalas University, 2nd Floor, Campus Limau Manis, 25163 Padang, West Sumatera, Indonesia
Fatchiyah Fatchiyah
Department of Biology, Faculty of Mathematics and Natural Sciences, Brawijaya University, Malang, East Java, Indonesia
Amri Bakhtiar
Faculty of Pharmacy, Andalas University, Padang, West Sumatera, Indonesia
Jamsari Jamsari
Laboratory of Biotechnology, Faculty of Agriculture, Andalas University, 2nd Floor, Campus Limau Manis, 25163 Padang, West Sumatera, Indonesia
LiveDNA: 62.12109
Asian Journal of Plant Pathology
Year: 2017 | Volume: 11 | Issue: 3 | Page No.: 139-147







ABSTRACT
Background and Objective: Plant-associated bacteria, such as Phyllobacterium play a significant role in protecting plant from pathogenic fungal infection. These Phyllobacterium are known to be existed in such a microbial community with various microbes thus leading to elevated disease suppression. The aim of this study was to assess the fungal suppression activity of indigenous Phyllobacterium isolates and its strain/species compatibility with rhizobacteria. Materials and Methods: Two indigenous Phyllobacterium isolates were identified using 16S rRNA gene sequence and its antifungal activities were tested against several phytopathogenic fungus. Further antagonistic assay was performed to compare the efficacy of cell culture and cell-free supernatants. Its compatibility was assayed by performing the antifungal assay using the combination of these Phyllobacterium isolates with rhizobacteria ones. Data were statistically analyzed using one-way analysis of variance and the significance was further processed using Duncan’s new multiple range test with a p<0.05. Results: Both isolates (UBCF_01 and UBCF_13), identified as Serratia plymuthica, exhibited higher suppression activity against Colletotrichum gloeosporioides compared to Fusarium oxysporum and Sclerotium rolfsii. Both isolates revealed opposite trend in their activities resulted from cultured cells and cell-free supernatants. Furthermore, the better suppression efficacy of the culture supernatants was resulted from single cultured cells, instead of co-culture. However, both isolates displayed quite poor compatibility with rhizobacteria isolates. Conclusion: These indigenous Phyllobacterium showed promising ability to be used as biocontrol agents for anthracnose. The application of its culture supernatants offered the less hazardous option of biological control implementation. However, their poor compatibility, even with the same species (rhizospheric UBCR_12) might be occurred due to habitat differences.
PDF Abstract XML References Citation
Received: April 16, 2017;
Accepted: June 10, 2017;
Published: June 15, 2017
How to cite this article
Siti Nur Aisyah, Sulastri Sulastri, Retmi Retmi, Rahmi Henda Yani, Elly Syafriani, Lily Syukriani, Fatchiyah Fatchiyah, Amri Bakhtiar and Jamsari Jamsari, 2017. Suppression of Colletotrichum gloeosporioides by Indigenous Phyllobacterium and its Compatibility with Rhizobacteria. Asian Journal of Plant Pathology, 11: 139-147.
URL: https://scialert.net/abstract/?doi=ajppaj.2017.139.147
URL: https://scialert.net/abstract/?doi=ajppaj.2017.139.147
INTRODUCTION
Phyllosphere located in aerial parts of plants is known to be one of habitat predominantly occupied by bacteria (with a density up to 108 cm–2 leaf)1,2. This habitat is considered as one of vital and ubiquitous environment, not only for bacteria but also for diverse microbial community1,3. However, unlike other plant parts, phyllosphere is a very harsh environment since it is exposed to rapid and fluctuating environmental stresses4. This hostility contributes to great microbial heterogeneity in the phyllosphere due to various mechanisms used by the colonizers to avoid or tolerate with this kind of habitat4,5. It also provides a broad array of plant-microbe interactions comprising of mutualism, commensalism, parasitism as well as pathogenicity2,3.
The presence of Phyllobacterium displayed symbiotic effects to their host by influencing plant growth and functions, such as production of growth promoting compounds and protection against pathogen infections6-10. As a biocontrol agent, these Phyllobacterium utilize its outstanding colonization ability to create preemptive exclusion of the pathogen11. Numerous species of Phyllobacterium, such as Paenibacillus sp., Pseudomonas sp., Serratia sp., Pantoea sp. and Myroides spp., have been reported for their capability in suppressing several phytopathogens11-15.
Colletotrichum is genus of pathogenic fungus which has been reported to infect numerous crops16. One of its species, C. gloeosporioides has become one of major pathogens worldwide by causing fruit rots (anthracnose) to at least 1000 plant species17. This species is also known as the most frequent species causing anthracnose in chili pepper, especially in tropical regions18-22. Due to this disease infection, significant yield loss and unreliable product quality become unavoidable23. Various biocontrol agents have been reported for its ability in controlling this fungal attack, i.e., Bacillus subtilis, Pseudomonas fluorescens, Streptomyces hygroscopicus and Oudemansiella mucida24-27.
The utilization of Phyllobacterium to overcome anthracnose infection is still far less documented. Regarding to its high tolerance against harsh environment, the competitive trait of Phyllobacterium could be utilized to managing this disease. Furthermore, this disease is difficult to be early managed due to its long-term incubation period known as quiescence phase until the symptom appeared visually during fruit ripening stage28. Outstanding colonization ability of this Phyllobacterium might block the symptom development through nutrient and space competition. Additionally, this study also examined the efficacy of cell-free supernatant application instead of living cell to develop more eco-friendly plant disease management. This study was aimed to investigate in vitro antifungal activity of two West Sumatera indigenous Phyllobacterium isolates against the anthracnose causing fungus C. gloeosporioides and its compatibility with rhizobacteria isolates.
MATERIALS AND METHODS
Isolation of Phyllobacterium isolates: In total of 120 isolates of indigenous Phyllobacterium were collected from phyllosphere of Brassica juncea L. in 2012 from District of Solok, West Sumatera, Indonesia. About 5 g of leaves were surface sterilized 3.5% sodium hypochlorite for 5 min and rinsed thoroughly three times using sterile distilled water. Leaves were subsequently grounded and cultured on a NaCl solution for 24 h under shaking condition. The bacteria were then isolated through serial dilutions and grown on Nutrient Agar (NA) medium for 2 days. Further selection was performed according to the morphological characteristics before subjected to antagonist assay. Bacterial isolates showing the highest antifungal activity would be used for further analysis.
Culture of fungi and bacterial isolates: Fungi used in this study consisted of three species of pathogenic fungi, i.e., Colletotrichum gloeosporioides, Sclerotium rolfsii and Fusarium oxysporum. Colletotrichum gloeosporioides was obtained from internal collection of Biotechnology Laboratory (Andalas University), while S. rolfsii and F. oxysporum were obtained from Phytopathology Laboratory (Andalas University) collection. Fungal mycelial disk (5×5 mm) was cultured onto PDA (Potato dextrose agar) medium (pH 7.0) and incubated at room temperature for 7 days in darkness.
In addition, two rhizobacteria isolates used in this study were Pseudomonas lurida strain UBCR_36 and Serratia plymuthica strain UBCR_12 collected from Biotechnology Laboratory (Andalas University). Both phyllo and rhizobacteria were grown on NA medium (pH 7.0) for 18 h at room temperature in darkness as well.
Species identification using 16S rRNA gene sequences: Two Phyllobacterium isolates showing the best suppression were molecularly identified using 16S rRNA gene sequences. This gene from each isolate was amplified using 27F (5’-AGAGTTTGATCTGGCTCAG-3’) and 1525R (5’-AAGGAGGTGWTCCARCC-3’) primers. This analysis was conducted using the same protocol as described by Syafriani et al.29.
Evaluation of bacterial inhibition range against pathogenic fungi: Inhibition range of these two isolates against three pathogenic fungi was tested through dual culture assay. The fungal mycelial disk was grown onto PDA (pH 7.0) for 48 h. Bacterial isolates were cultured in Luria Broth (LB) (pH 7.0) at 160 rpm room temperature for 12 h (OD600 nm = 1.0). Paper disks (diameter 5 mm) were impregnated with 20 μL of cell culture and applied at 4 different positions located 3 cm away from the center of fungal growth. The plate was subsequently incubated at room temperature for 7 days. Inhibition was measured daily using this following formula30:
| (1) |
where, DC is the diameter of untreated fungus spot and DT is the diameter of fungus spot treated with bacteria30. Each treatment was performed in five replicates.
Co-cultivation of bacterial isolates and chili pepper isolated C. gloeosporioides: The protocol used for this co-cultivation was adapted from Zhang et al.31 with several modifications. The culture of C. gloeosporioides was performed using a disk (5×5 mm) in 100 mL potato dextrose broth (pH 7.0) medium for 72 h under shaking condition at 100 rpm and room temperature. Bacteria cells were then cultured in LB medium (pH 7.0) until it reached OD600 nm = 1.0. Bacterial pellet was collected through centrifugation (20,000×g at 4°C for 15 min) and washed three times with dH2O. About 5 mL of cells suspension (3×108 CFU mL–1) was then inoculated into the fungal culture and grown for 24 h at room temperature under agitated condition (100 rpm). Fungal mycelia were discarded and the bacteria was pelleted by centrifugation (20,000×g at 4°C for 15 min). The supernatants were then used for inhibition assay.
Antifungal assay of bacterial culture supernatants against C. gloeosporioides: The CSN of UBCF_01 and UBCF_13 were subsequently tested for its antifungal activity against chili pepper isolated C. gloeosporioides. The CSN used for this assay were divided into two categories, single culture and co-culture supernatants. This assay was performed using agar well plate method and each treatment was performed in five replicates. CSN, either from single culture or co-culture were harvested by centrifugation (20000×g at 4°C for 15 min) and subsequently filtered using a membrane with a pore size of 0.22 μm. An aliquot (50 μL) of the cell-free filtrate was then applied on 3 cm away from 2 days-aged C. gloeosporioides at four different positions.
Evaluation of CSN compatibility between phyllo and rhizobacteria isolates: The bacterial CSN compatibility for consortium type application was evaluated by combining the culture supernatants of phyllo and rhizobacteria isolates obtained from co-culture. Cell-free culture supernatants from each isolate were applied in the form of dual, triple and quadruple strain consortium with the mixing ratio 1:1 (v/v). Mixed culture supernatants (50 μL) was then subjected to dual culture medium containing 2 days-aged C. gloeosporioides and grown for 7 days at room temperature. The resulting inhibition was routinely measured using the formula as described previously.
Statistical analysis: Data were statistically analyzed using SPSS version 23.0 and presented with mean and standard deviation. Data from inhibition range assay and strains compatibility was processed by one-way analysis of variance (ANOVA). Significant differences from each treatment group were further processed using Duncan’s New Multiple Range Test (DNMRT) with a p<0.0532.
RESULTS
Species identity of Phyllobacterium isolates: Of 120 bacterial isolates collected from B. juncea leaves, 23 isolates showed antifungal activity against C. gloeosporioides and the highest suppression was resulted by two isolates, named UBCF_01 and UBCF_13. Based on the sequence of 16S rRNA genes, these two isolates were identified as Serratia plymuthica. Gene sequence from both isolates which covered 1534 bp has been deposited in the NCBI database with accession number KX394778 (UBCF_01) and KX394779 (UBCF_13). From a BLAST homology search, high homology (99%) of the gene sequence was shown between S. plymuthica UBCF_01 (KX394778) with other S. plymuthica species, such as S. plymuthica UBCF_13 (KX394779), S. plymuthica UBCR_12 (KU299959) and S. plymuthica AS9 (CP002773). Based on the phylogenetic tree, S. plymuthica UBCF_01 displayed closer genetic relationship with S. plymuthica UBCR_12 (KU299959) than with S. plymuthica UBCF_13 (Fig. 1).
Antifungal activity range of Phyllobacterium isolates: Both Phyllobacterium isolates, UBCF_01 and UBCF_13 displayed antifungal activity against C. gloeosporioides but no inhibition of F. oxysporum and S. rolfsii was observed (Fig. 2).
 | |
| Fig. 1: | Genetic relationship of both Phyllobacterium isolates with other Serratia species based on the sequence of 16S rRNA gene |
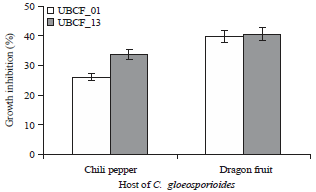 | |
| Fig. 2: | Inhibition spectrum of Phyllobacterium isolates against C. gloeosporioides from different host |
| Values are Means±SD (n = 5) | |
Of the two C. gloeosporioides phytovars used, the suppression by both Phyllobacterium was stronger against the one isolated from dragon fruit compared to the chili pepper one. However, each isolate showed similar inhibitory effect against dragon fruit isolate of C. gloeosporioides. Unlike the dragon fruit phytovar, UBCF_13 showed significantly higher suppression (p<0.05) than UBCF_01 at 7 days post application (DPA), respectively 34 and 26% (Fig. 2). However, the best suppression was still obtained from UBCF_13 about 41% against dragon fruit phytovar of C. gloeosporioides (Fig. 2). This result suggested that the antifungal activities of UBCF_01 and UBCF_13 are very specific for isolates of C. gloeosporioides; while they have no measurable inhibitory effect on F. oxysporum and S. rolfsii.
Effect of culture supernatants (CSN) on fungal suppression: Application of CSN from cultured Phyllobacterium showed similar inhibitory effects as the complete cell culture (Fig. 2) on C. gloeosporioides. Figure 3 shows that the single-cultured CSN from each isolate exhibited similar suppression trend. Of these two isolates, CSN from UBCF_01 exhibited higher inhibition at 7 DPA (30%) compared to the one from UBCF_13 CSN (26%) (Fig. 3). Despite having similar overall suppressions, the effect of UBCF_01 CSN was observed early (2 DPA) (Fig. 3), while inhibition by UBCF_13 was only detected at 4 DPA. The results imply that the two isolates suppress fungal growth by different substances or mechanisms. The UBCF_01 had a rapid effect but this was not very sustainable and it appeared that the fungus adapted. In contrast, UBCF_13 showed a delayed effect but this was better maintained.
In contrast, CSN effect from single-cultured UBCF_13 started to retard the fungal growth and displayed a measurable suppression at 4 DPA (Fig. 3). According to the suppression pattern, it showed that there might be a sudden effect at 4 DPA that was successfully maintained until 7 DPA, so that the suppression seemed to increase continuously. However, since no inhibition recorded at previous days, it is assumed that the distance between the CSN application site and the fungal growth might be one of the barriers. Thus, the fungal enabled to grow freely but then started to be suppressed when its growth was more closer to the CSN application site.
Unlike the single-cultured CSN, fungal growth was less affected by the CSN from bacteria co-cultured with fungus (Fig. 3). These results showed that the antifungal activity of both bacterial strains is not stimulated by the presence of the pathogen. Even though the suppression was clearly occurred during the co-culture and marked by the remarkable mycelial mass difference but it still could not be correlated with the CSN efficacy used in the antagonistic assay. Due to various unspecified substances contained in the CSN, the measurement to determine the concentration is seemingly unable to be performed.
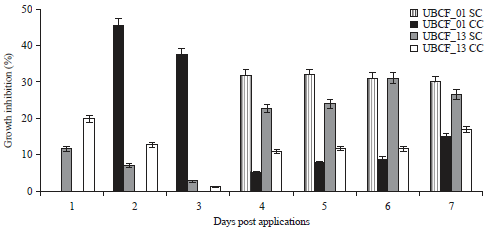 | |
| Fig. 3: | Difference in fungal suppression efficacy of each Phyllobacterium culture supernatant resulted from different types of culture |
| SC: Single culture and CC: Co-culture, values are Mean±SD (n = 5) | |
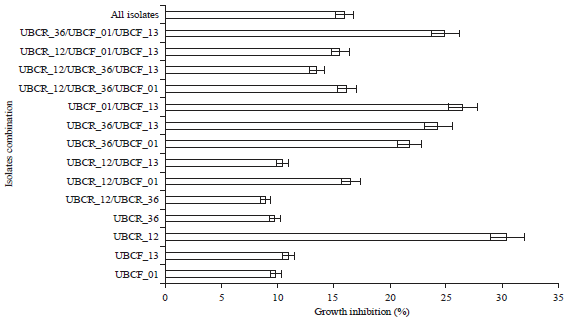 | |
| Fig. 4: | Effect of strain/species compatibility towards the resulting suppression against C. gloeosporioides at 7 days post application |
| Values are Mean±SD (n = 5) | |
CSN compatibility between phyllo and rhizobacteria isolates: To improve the inhibition efficacy of the Phyllobacterium isolate, the CSN were applied in a mixture with rhizobacteria isolates. Figure 4 shows that both single and mixed applications resulted in different effects regarding the fungal suppression activity against chili pepper isolated C. gloeosporioides. This may indicate various compatibilities or mutual inhibition of antifungal effects. It showed clearly that combination of Phyllobacterium with rhizospheric UBCR_36 resulted higher inhibitory effect rather than the single isolate one. In contrast, mixed CSN application involving rhizospheric UBCR_12 showed relatively lower suppression than other combinations.
Compared to the single application, both Phyllobacterium isolates exhibited higher inhibition with consortium application. The combination of both Phyllobacterium strains resulted in the highest suppression (about 26%) (Fig. 4). Both isolates were also well compatible with P. lurida UBCR_36 although the suppression was not significantly enhanced. Double consortium of UBCR_36 with each Phyllobacterium isolate displayed quite similar suppression activity as the consortium consisted of these three isolates (Fig. 4).
Unlike P. lurida UBCR_36, rhizobacteria isolate S. plymuthica UBCR_12 exhibited weak synergistic effect with other isolates, including the same S. plymuthica species. Furthermore, CSN mixtures of both rhizobacteria isolates repressed its inhibitory activity. Compared to UBCF_13 and UBCR_36, UBCR_12 showed better compatibility with UBCF_01. From the consortium of UBCR_12-UBCF_01, the highest suppression was observed compared to other UBCR_12 combinations. The presence of UBCF_01 in the consortium of triple isolates might improve the weak compatibility between UBCR_12 and UBCR_36. Nonetheless, the resulting inhibitions under consortium did not surpass its suppression when applied singly (30.54%) (Fig. 4). Combining all four isolates showed a weak effect on fungal inhibition indicating that there was no additive or synergistic mechanism. In the contrary, it rather appeared that the antifungal effects negatively interfered with each other.
DISCUSSION
Result of this study had proven that phyllospheric S. plymuthica could be developed as one of choice for anthracnose biocontrol agents (Fig. 2). This species is rarely reported for managing C. gloeosporioides. Recent studies reported that the rhizospheric S. plymuthica UBCR_12 displayed high suppression against C. gloeosporioides29,33. Compared to the rhizosperic S. plymuthica from previous work33, these phyllospheric species exhibited different characteristic in suppressing C. gloeosporioides. It might be associated with lifestyle difference as the consequence of habitat difference, even the genetic relationship between both strains were closely related (Fig. 1). Antifungal activity of both Phyllobacterium isolates seemed to be more independent and specific only against C. gloesporioides (Fig. 2). Such characteristic had been previously reported by Zachow et al.34 where the phyllospheric Pseudomonas sp. only showed remarkable suppression against R. solani of four pathogens tested.
Phyllobacterium is also known as highly competitive colonizer. This trait affects their ability to protect the host plant by suppressing the pathogen growth. Their prominent plant-protective effect is occurred due to their high abundance. Although this abundance related to the total cell number is not the key factor determining their protection but this high density population may be associated to the resources competition9,11. This kind of mechanism is termed as pre-emptive colonization which is mainly stimulated by niche occupation35. Several studies reported that nutritional similarity could bottle up the pathogen’s secondary colonization and lead to better antagonistic effect9,36,37.
This study was also proved that cell-free culture supernatants (CSN) of both Phyllobacterium isolates were able to suppress the growth of C. gloeosporioides (Fig. 3). In addition, both isolates showed that higher antifungal effect was resulted only from single culture. This result suggested that these two isolates did not require to be induced by fungal cells to obtain higher suppression efficacy. Regarding to this characteristic, these Phyllobacterium isolates displayed better option for the ease of mass production compared to rhizospheric UBCR_12 from previous study33. According to this result, the utilization of biocontrol agents for large-scale field application could be applied in more eco-friendly way through this CSN, instead of using living cell. Since it is cell free, it could reduce the risk of uncontrolled bacterial spreading and effects on ecosystem balance when applied in high volume. The CSN is mostly composed of secreted secondary metabolites that play a crucial role in defense mechanisms against biotic and abiotic stresses. Its production is usually occurred during the late growth phase of bacteria and is greatly stimulated by several factors, such as nutrient compositions38.
To develop an efficient CSN application, CSN effect should be measured by a proper method that could accurately explain the CSN efficacy on pathogenic fungus. Figure 3 shows that the inhibitory effect of CSN from both isolates was appeared quite slowly. This slow occurrence could be one of limitations that affect the measurement, so that, the exact efficacy of this CSN could be precisely determined. It was also predicted that this slow effect could be associated to diffussion problem. The selection of agar diffusion method to be used in this study was considered due to its simplicity, low cost, low amount of sample required and easy to interpret39. However, this method showed some limitations, such medium incompatibility and substance instability problems occurred during diffusion process40.
Application of microbial consortium had been widely studied to observe the possibility of obtaining higher suppression efficacy through the combination of diverse mechanisms from each microbial component11,41,42. However, application of microbial consortium using bacterial CSN as conducted in this study, was still less documented. Figure 4 shows that CSN combination composed of both Phyllobacterium isolates resulted in elevated suppression compared to single isolate application. Moreover, both isolates exhibited remarkable compatibility with rhizospheric P. lurida UBCR_36. However, this synergistic effect was quite unseen from the CSN combination of both phyllo and rhizospheric S. plymuthica. Successful consortium demands the high compatibility among the involving strain or species to cooperate synergistically or at least additive on the disease suppression. Incompatible combinations might result in the decrease or even loss of inhibitory effects42-44.
CONCLUSION AND FUTURE RECOMMENDATIONS
It is concluded that the Phyllobacterium isolates UBCF_01 and UBCF_13 are promising alternative biocontrol agents against C. gloeosporioides. Supernatants from cultures of these bacteria were also effective. Field application of bacterial CSN instead of living cells would provide a less hazardous approach to plant disease management. However, these supernatants were only tested under controlled conditions in the absence of plants. The supernatants should be further tested directly with infected plants both in the greenhouse and then in the field.
This present study presented new option of biocontrol agent against C. gloeosporioides in chili pepper by utilizing the indigenous Phyllobacterium. It also offered safer way in biocontrol application through bacterial cell-free CSN to minimize the risk of uncontrolled bacterial spreading when applied in large-scale. Elevated suppression resulted from strain/species combinations suggested that presence of other antagonist bacteria could maximize the resulting inhibitory effect.
SIGNIFICANCE STATEMENTS
This study discovers an easily accessible yet eco-friendly choice for biological agents to manage the anthracnose infection in chili pepper. It will help the researchers to develop better plant diseases management system for field scale with lower risk of biological contaminations due to direct application of biocontrol agents that was not fully explored, yet. Thus, a new idea on these applications of biocontrol agents and possibly other improvement, may be arrived at.
ACKNOWLEDGMENTS
This research is funded by Directorate General of Higher Education-Ministry of Research and Technology via PMDSU research grant fiscal year 2015-2016, contract No. 124/SP2H/LT/DRPM/III/2016, on March 6th, 2016. Several data are also supported by the Competency Grant, contract No. 020/SP2H/LT/DRPM/II/2016, on February 17th, 2016.
REFERENCES
- Whipps, J.M., P. Hand, D. Pink and G.D. Bending, 2008. Phyllosphere microbiology with special reference to diversity and plant genotype. J. Applied Microbiol., 105: 1744-1755.
CrossRefDirect Link - Vorholt, J.A., 2012. Microbial life in the phyllosphere. Nat. Rev. Microbiol., 10: 828-840.
CrossRefDirect Link - Baker, C.M., R. Chitrakar, N. Obulareddy, S. Panchal, P. Williams and M. Melotto, 2010. Molecular battles between plant and pathogenic bacteria in the phyllosphere. Braz. J. Med. Biol. Res., 43: 698-704.
CrossRefDirect Link - Meyer, K.M. and J.H. Leveau, 2012. Microbiology of the phyllosphere: A playground for testing ecological concepts. Oecologia, 168: 621-629.
CrossRefDirect Link - Fedorov, D.N., N.V. Doronina and Y.A. Trotsenko, 2011. Phytosymbiosis of aerobic methylobacteria: New facts and views. Microbiology, 80: 443-454.
CrossRefDirect Link - Partida-Martinez, L.P.P. and M. Heil, 2011. The microbe-free plant: Fact or artifact? Frontiers Plant Sci., Vol. 2.
CrossRef - Reed, S.C., A.R. Townsend, C.C. Cleveland and D.R. Nemergut, 2010. Microbial community shifts influence patterns in tropical forest nitrogen fixation. Oecologia, 164: 521-531.
CrossRefDirect Link - Innerebner, G., C. Knief and J.A. Vorholt, 2011. Protection of Arabidopsis thaliana against leaf-pathogenic Pseudomonas syringae by Sphingomonas strains in a controlled model system. Applied Environ. Microbiol., 77: 3202-3210.
CrossRefDirect Link - Balint-Kurti, P., S.J. Simmons, J.E. Blum, C.L. Ballare and A.E. Stapleton, 2010. Maize leaf epiphytic bacteria diversity patterns are genetically correlated with resistance to fungal pathogen infection. Mol. Plant-Microbe Interact., 23: 473-484.
CrossRefDirect Link - Stockwell, V.O., K.B. Johnson, D. Sugar and J.E. Loper, 2011. Mechanistically compatible mixtures of bacterial antagonists improve biological control of fire blight of pear. Phytopathology, 101: 113-123.
CrossRefDirect Link - Marimuthu, S., V. Ramamoorthy, R. Samiyappan and P. Subbian, 2013. Intercropping system with combined application of Azospirillum and Pseudomonas fluorescens reduces root rot incidence caused by Rhizoctonia bataticola and increases seed cotton yield. J. Phytopathol., 161: 405-411.
CrossRefDirect Link - Muller, T., U. Behrendt, S. Ruppel, G. von der Waydbrink and M.E. Muller, 2016. Fluorescent pseudomonads in the phyllosphere of wheat: Potential antagonists against fungal phytopathogens. Curr. Microbiol., 72: 383-389.
CrossRefDirect Link - Erlacher, A., M. Cardinale, R. Grosch, M. Grube and G. Berg, 2014. The impact of the pathogen Rhizoctonia solani and its beneficial counterpart Bacillus amyloliquefaciens on the indigenous lettuce microbiome. Front. Microbiol., 5: 1-8.
CrossRefDirect Link - Sato, I., S. Yoshida, Y. Iwamoto, M. Aino and M. Hyakumachi et al., 2014. Suppressive potential of Paenibacillus strains isolated from the tomato phyllosphere against Fusarium crown and root rot of tomato. Microbes Environ., 29: 168-177.
CrossRefDirect Link - Hyde, K.D., L. Cai, P.F. Cannon, J.A. Crouch and P.W. Crous et al., 2009. Colletotrichum-names in current use. Fungal Divers., 39: 147-182.
Direct Link - Phoulivong, S., L. Cai, H. Chen, E.H.C. McKenzie, K. Abdelsalam, E. Chukeatirote and K.D. Hyde, 2010. Colletotrichum gloeosporioides is not a common pathogen on tropical fruits. Fungal Divers., 44: 33-43.
CrossRefDirect Link - Than, P.P., R. Jeewon, K.D. Hyde, S. Pongsupasamit, O. Mongkolporn and P.W.J. Taylor, 2008. Characterization and pathogenicity of Colletotrichum species associated with anthracnose on chilli (Capsicum spp.) in Thailand. Plant Pathol., 57: 562-572.
CrossRefDirect Link - Than, P.P., H. Prihastuti, S. Phoulivong, P.W.J. Taylor and K.D. Hyde, 2008. Chilli anthracnose disease caused by Colletotrichum species. J. Zhejiang Univ. Sci. B, Vol. 9.
CrossRefDirect Link - Mongkolporn, O., P. Montri, T. Supakaew and P.W.J. Taylor, 2010. Differential reactions on mature green and ripe chili fruit infected by three Colletotrichum spp. Plant Dis., 94: 306-310.
CrossRefDirect Link - Ratanacherdchai, K., H.K. Wang, F.C. Lin and K. Soytong, 2007. RAPD analysis of Colletotrichum species causing chilli anthracnose disease in Thailand. J. Agric. Technol., 3: 211-219.
Direct Link - Suwan, N., W. Boonying and S. Nalumpang, 2012. Antifungal activity of soil actinomycetes to control chilli anthracnose caused by Colletotrichum gloeosporioides. J. Agric. Technol., 8: 725-737.
Direct Link - Kim, S.H., J.B. Yoon, J.W. Do and H.G. Park, 2008. A major recessive gene associated with anthracnose resistance to Colletotrichum capsici in chili pepper (Capsicum annuum L.). Breed. Sci., 58: 137-141.
CrossRefDirect Link - Zhao, X., X. Zhao, Y. Wei, Q. Shang and Z. Liu, 2013. Isolation and identification of a novel antifungal protein from a rhizobacterium Bacillus subtilis strain F3. J. Phytopathol., 161: 43-48.
CrossRefDirect Link - Fitsum, S., M. Amin, T. Selvaraj and A. Alemayehu, 2014. In vitro evaluation of some fungicides and bioagents against common bean anthracnose (Colletotrichum lindemuthianum Sacc. and Magnus) Briosi and Cavara. Afr. J. Microbiol. Res., 8: 2000-2005.
Direct Link - Prapagdee, B., C. Kuekulvong and S. Mongkolsuk, 2008. Antifungal potential of extracellular metabolites produced by Streptomyces hygroscopicus against phytopathogenic fungi. Int. J. Biol. Sci., 4: 330-337.
CrossRefPubMedDirect Link - Syafriani, E., F. Riwany, R. Kamelia, I. Ferita, F. Fatchiyah and J. Jamsari, 2016. A promising novel rhizobacteria isolate UBCR_12 as antifungal for Colletotrichum gloeosporioides. Res. J. Pharm. Biol. Chem. Sci., 7: 2202-2209.
Direct Link - Islam, M.R., Y.T. Jeong, Y.S. Lee and C.H. Song, 2012. Isolation and identification of antifungal compounds from Bacillus subtilis C9 inhibiting the growth of plant pathogenic fungi. Mycobiology, 40: 59-66.
CrossRefDirect Link - Zhang, C., X. Zhang and S. Shen, 2014. Proteome analysis for antifungal effects of Bacillus subtilis KB-1122 on Magnaporthe grisea P131. World J. Microbiol. Biotechnol., 30: 1763-1774.
CrossRefDirect Link - Aisyah, S.N., H. Harnas, S. Sulastri, R. Retmi and H. Fuaddi et al., 2016. Enhancement of a novel isolate of Serratia plymuthica as potential candidate for an antianthracnose. Pak. J. Biol. Sci., 19: 250-258.
CrossRefDirect Link - Zachow, C., R. Tilcher and G. Berg, 2008. Sugar beet-associated bacterial and fungal communities show a high indigenous antagonistic potential against plant pathogens. Microbial Ecol., 55: 119-129.
CrossRefDirect Link - Remus‐Emsermann, M.N.P., G.A. Kowalchuk and J.H.J. Leveau, 2013. Single-cell versus population-level reproductive success of bacterial immigrants to pre-colonized leaf surfaces. Environ. Microbiol. Rep., 5: 387-392.
CrossRefDirect Link - Smits, T.H., F. Rezzonico, T. Kamber, A. Goesmann and C.A. Ishimaru et al., 2010. Genome sequence of the biocontrol agent Pantoea vagans strain C9-1. J. Bacteriol., 192: 6486-6487.
CrossRefDirect Link - Stockwell, V.O., K.B. Johnson, D. Sugar and J.E. Loper, 2010. Control of fire blight by Pseudomonas fluorescensA506 and Pantoea vagans C9-1 applied as single strains and mixed inocula. Phytopathology, 100: 1330-1339.
CrossRefDirect Link - Ruiz, B., A. Chavez, A. Forero, Y. Garcia-Huante and A. Romero et al., 2010. Production of microbial secondary metabolites: Regulation by the carbon source. Crit. Rev. Microbiol., 36: 146-167.
CrossRefPubMedDirect Link - Balouiri, M., M. Sadiki and S.K. Ibnsouda, 2016. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal., 6: 71-79.
CrossRefDirect Link - Bonev, B., J. Hooper and J. Parisot, 2008. Principles of assessing bacterial susceptibility to antibiotics using the agar diffusion method. J. Antimicrob. Chemother., 61: 1295-1301.
CrossRefDirect Link - Vacheron, J., G. Desbrosses, M.L. Bouffaud, B. Touraine and Y. Moenne-Loccoz et al., 2013. Plant growth-promoting rhizobacteria and root system functioning. Frontiers Plant Sci., Vol. 4.
CrossRef - Sarma, B.K., S.K. Yadav, S. Singh and H.B. Singh, 2015. Microbial consortium-mediated plant defense against phytopathogens: Readdressing for enhancing efficacy. Soil Biol. Biochem., 87: 25-33.
CrossRefDirect Link - Castillo, B.M., M.F. Dunn, K.G. Navarro, F.H. Melendez, M.H. Ortiz, S.E. Guevara and G.H. Palacios, 2016. Antifungal performance of extracellular chitinases and culture supernatants of Streptomyces galilaeus CFFSUR-B12 against Mycosphaerella fijiensis Morelet. World J. Microbiol. Biotechnol., Vol. 32.
CrossRef - Walker, V., O. Couillerot, A. von Felten, F. Bellvert and J. Jansa et al., 2012. Variation of secondary metabolite levels in maize seedling roots induced by inoculation with Azospirillum, Pseudomonas and Glomus consortium under field conditions. Plant Soil, 356: 151-163.
CrossRefDirect Link