
Donatha Damian Tibuhwa
Department of Molecular Biology and Biotechnology, College of Natural and Applied Sciences, University of Dar es Salaam, Tanzania
Shamte Shomari
Naliendele Agriculture Research Institute, Mtwara, Tanzania
Asian Journal of Plant Pathology
Year: 2016 | Volume: 10 | Issue: 4 | Page No.: 36-48







ABSTRACT
Background and Objective: The emergence of unknown pathogens that cause massive wilting of the cashew tree in Magawa, Coast region, Tanzania has caused big turmoil to famers. Many families are frustrated and have cut down the wilting cashews leaving their farm bear, while others shifted to alternative food crops which has caused unsustainable life due to decrease in their household income and generally increased poverty. This study presents the scientific investigations on the causes of the diseases; gives scientific explanations to related farmers field observations and provides some way forward in mitigating the chaos. Materials and Methods: Field investigations were carried out including face-to-face interview to the farmers. Laboratory investigation involved isolation and characterisation of the pathogen using both morphological and molecular markers as well as pathogenicity test to prove the disease causative agent. Results: Field investigation showed that the wilting disease becomes epidemic within few weeks of its first appearance, leading to complete cashew tree loss especially after rain season. The laboratory investigations and pathogenicity test established the pathogen-causing cashew wilting disease to be Fusarium oxysporum. Conclusion: This is the first study of Fusarium species being associated with cashew wilt disease. The study thus, draws an attention to other cashew growers in the world of the emerging threat of Fusarium wilting diseases to cashew crop.
PDF Abstract XML References Citation
Received: July 29, 2016;
Accepted: August 20, 2016;
Published: September 15, 2016
How to cite this article
Donatha Damian Tibuhwa and Shamte Shomari, 2016. Fusarium Wilt Disease: An Emerging Threat to Cashew Nut Crop Production in Tanzania. Asian Journal of Plant Pathology, 10: 36-48.
URL: https://scialert.net/abstract/?doi=ajppaj.2016.36.48
URL: https://scialert.net/abstract/?doi=ajppaj.2016.36.48
INTRODUCTION
Cashew (Anacardium occidentale Linn) is an unusual nut grown on the outside of the cashew apple. Cashews are well known for their health benefits such as rich in unsaturated fatty acids, plant protein, dietary fiber, antioxidant vitamins (e.g., vitamin E and tocopherols), minerals (e.g., magnesium and potassium) and phytochemicals such as flavonoids1,2. In East Africa, cashew production was introduced in the 16th century by Portuguese. The good characteristics of cashew trees including its tolerance of drought, ability to grow on poor soils and to be intercropped with food crops makes it an ideal product for small farmers.
In Tanzania, cashew nut is the most important export crop in terms of foreign exchange earnings replacing coffee that dominated since independence3 and an important source of income for small farmers in the Southern coastal region. Cashew production in the country is facing various problems including powdery mildew disease4, which may cause crop losses ranging between 70-100%5 and other effects augmented by effects of climate changes6. The emergence of the unknown cashew wilt diseases in Magawa area located in Mkuranga district coast region Tanzania, which is among the cashew grower has added up another challenge to the growers and subjected them to increasing poverty, malnutrition and general frustrations that has caused some of them to abandon the crop. This outbreak poses devastating consequences since it attacks and causes wilting of cashew trees massively, rendering a cashew farm into dying cashew tree logs within a short period of time. Farmers noted the wilting of the cashew trees in not less than three years ago and many cashew trees have been affected and completely wilted out. Wilting diseases in various vascular plants has been associated with pathogens that produces high and low molecular weight phytotoxins during host colonization which cause wilting symptoms7,8. The wilting caused by then unknown reason to Magawa cashew growers becomes epidemic within few weeks, leading to complete crop loss especially the period after the rain season. This has led to some farmers abandoning cultivating cashew trees. The abandoning of cashew cultivation by many families has caused unsustainable life due to decrease in their household income and general poor living conditions. The high frequency of death of cashew nut tree in Mkuranga area has attracted this study to identify the causes of cashew plant wilting and total death. The objectives of the present study were to investigate and establish the causes of the wilting of the cashew trees in Magawa village whereby both conventional and molecular characters were used in identifying the pathogen while pathogenicity test was carried out to authenticate the causative agent of wilting.
MATERIALS AND METHODS
Plant material and soil sampling: Field surveys which involved physical observation of the cashew wilting effects to farmers (Fig. 1), collecting plant material as well as soil samples, involved complete uprooting of both already wilted trees and healthy trees that were not showing any symptoms. Both symptomatic and asymptomatic trees were collected in duplicate each, in two field trips done in two consecutive years.
Trees which were not showing any symptoms were collected in two different sites, one farm with some affected trees showing all the symptoms and another from a quite distant of about 2 km apart, showing completely no symptom on external observation (Fig. 2a, b).
 | |
| Fig. 1(a-b): | Photographs showing (a) Wilting cashew due to pathogen infection and (b) Cashew farm abandoned and turned into alternative food crops. Photo taken by the researchers in Magawa-coast region Tanzania |
 | |
| Fig. 2(a-c): | Cashew tree showing different stages, (a) Healthy, (b) Severe symptoms but few roots supplying nutrients for the survived branches and (c) Complete wilted tree. Photo taken by the researchers in Magawa-coast region Tanzania |
 | |
| Fig. 3(a-b): | Cashew tree (a) Branches and stem cut using chain saw and (b) Cross sections made in the field |
The collected parts of the trees involved leaves, stem, branches, adventitious roots and cross section of different parts of the plant roots and collar. Affected trees and cross sections were photographed in situ (Fig. 3). Cross sections were properly made using chain saw. The cut sections and roots were then packed into plastic bags, which were correctly labelled with collection number, date, from infected or non infected tree as well as proper locality where it was collected. They were then brought to the Department of Molecular Biology and Biotechnology Laboratory of the University of Dar es Salaam for further studies and pathogen isolation.
For the soils, samples were collected along the soil profile after soil excavations to expose the different horizons. The samples were collected using soil auger at different depths made under the excavated tree trunk of both affected and unaffected trees at 0-10, 10-40, 40-80, 80-120 and 120-140 cm, respectively. The soils were immediately packed into air-tight plastic bags, labelled correctly with the date, depth, sampling locality including whether from affected or unaffected tree trunk. Soils were sampled from three pits dug within 3 m and were used to assess the possible effect on root damage by excess salt, heavy metals and pathogen.
Laboratory work
Isolation of biotic disease causal agents (pathogen): In order to determine the cause of an infection in a diseased plant, isolation of possible causing pathogen was done from infected tissues. In the laboratory, the first step was to place a sample of the diseased tissue under condition that will allow an infectious agent to grow and possibly induce sporulation. To facilitate this, pieces of infected cashew plant tissue were first surface sterilized with 0.5% of sodium hypochlorite for 3 min and rinsed twice in sterile water. They were then placed on three different nutrient media Malt Extract Agar (MEA), Potato Dextrose Agar (PDA) and MacConkey agar (MAC) for 3 days at 25°C (Fig. 4). Growth was so evident with PDA, thus the experiment was continued with PDA media.
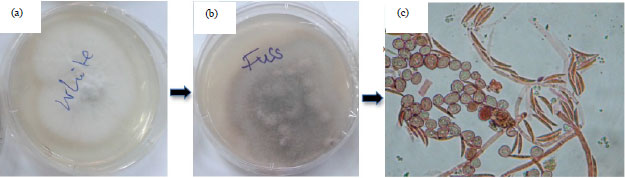 | |
| Fig. 4(a-c): | Isolated Fusarium oxysporum (a) Young stage, (b) Old stage and (c) Microscopic features |
Isolation of the pathogen was performed following the protocol of Perez-Vicente et al.9 where by about 0.8 cm of infested cashew tissues from vascular vessels were put in a petri dish containg Potato Dextrose Agar (PDA) and antibiotic streptomycin 1.2 mL/240 of PDA. This was followed by growing organisms into pure culture by plate dilution method and streaking plates as detailed in Perez-Vicente et al.9. Sampling portions were both from the margins of the diseased tissue where the pathogen is more numerous which help ed to avoid taking saprophytes that quickly colonize the recently killed tissue and also from sampled soil up to 40 cm deep.
Pathogen identification and morphological characterization: Morphological characters of the pathogen were described from the obtained cultures on PDA. For macromorphological characteristics of the pathogen we directly used them for the observation of the colony morphology as well as pigmentation and observation of the growth rates. However, for microscopic characterization, PDA pure cultures were transferred on Carnation Leaf Agar (CLA) and after 10 days of growth, the micromorphological characteristics were observed and evaluated according to Burgess et al.10 and Leslie and Summerell11 using the compound light microscope at 40X magnification of a bright field compound microscope (Olympus BX50 Phase Pol Darkfield Microscope, Japan). The observations and measurements were taken on a digital microscope whereby the microscopic features were directly observed on the computer screen connected to the microscope, equipped with micrometer scale which was adjusted for reading accurately the measurement of the feature and wherever necessary the snap shot of the feature taken directly and saved on digital format on the attached computer. The pathogen asexual spores (microconidia, macroconidia and chlamydospores) and mycelia specifically their structures and shapes were clearly observed and compared with already published study of Nelson et al.12 Burgess et al.10, Leslie and Summerell11, Thangavelu and Mustaffa13 and Pataky14 internet facility, which helped in establishing the pathogen identity conventionally.
Cashew anatomy study for observing affected parts: In order to determine the parts or tissues of the plant that were affected by the pathogen, observations were done on a microscope for the conducting tissues systems of roots and shoot of both affected and healthy plant. The plant cross-sections of both healthy and wilted plants were obtained by using both hand sections and microtome in the laboratory. After cross sectioning, the sections were stained for anatomical study using both safranin and light green (a common stain for vascular tissue anatomical study). The staining procedures involved placing the sections on petri dishes containing safranin for 10 min followed by washing in commercial alcohol and acid alcohol for differentiating. Washing with alcohol were repeated to remove all traces of acid. After washing the sections were mounted on slides and covered by cover slips. The tissue was examined for the presence of pathogen using compound microscope Olympus cx42 at 40X magnification. Sections from unaffected plant cultivars were used as control.
Pathogenicity test: It is well known that fungi use several mechanisms to invade plants. These mechanisms can generally be observed during pre-entry, entry and colonization. This study conducted pathogenicity test following Koch’s postulate theory that involved inoculation of healthy plants (Anacardium occidentale L.) with the isolated pathogen. The cashew seedlings grown in controlled conditions in unaffected zone were bought and inoculated with a suspected pathogenic organism, i.e., the pathogen pure culture was obtained. The spores suspensions were prepared by adding water to the cultures and mixing in a blender. The suspensions were strained through two layers of cheesecloth, diluted with distilled water to a concentration of 1×105 conidia mL–1 as detailed in Al-Juboory and Juber15. Inoculum was used within 2 h of preparation.
Experiments: Experiment was conducted in vitro in the green house of Botany Department of the University of Dar es Salaam. Ten milliliters of spore suspensions at 1×105 spores mL–1 of isolate was injected separately near the rooting system of both damaged with small cuts introduced purposely using surgical blade (sterilized by GAMA radiation 25 KGY) and undamaged without any cuts together with two control damaged and none damaged injected with distilled water for control. The cashew plants were in duplicates and were kept in the growing bags placed in a bucket with 2 m spacing to avoid the chances of spreading the pathogen. Sterile water was gradually added on the cultivar in order to maintain the humidity. Observations of the symptoms in the set experiment were carried out consistently for every week for three months to allow the completion of Koch’s postulate theory. The pathogen was then re isolated following the same procedure on the infested inoculated seedlings with cuts after showing similar symptoms.
Soil analysis: Soil pH and Electric Conductivity (EC) were measured in water suspension (1:2). For determination of concentrations of soluble microelements and heavy metals (Mn, Fe, Ni, Cu, Zn, Cd and Pb) were extracted using DTPA and HCl. The soils were digested in a mixture of HNO3 and HClO4 (S.1) (9:1). Their respective amounts in the digests then measured by atomic absorption spectrometry AA-6800 of the make Shimadzu, Kyoto Japan as detailed by Tijjani et al.16.
Culture sensitivity test: In order to establish the best mitigation method in combating the pathogen, this study carried out culture sensitivity test to come up with the best chemical that kills the pathogen. Isolated pathogen was tested for its sensitivity to five broad-spectrum antifungal chemicals namely: Benomyl (Fundazol WP, 500 g L–1), carbendazim (Kolfugo super SC, 200 g L–1), azoxystrobin (Quadris SC, 250 g L–1), fludioxonil (Maxim SC, 25 g L–1) and bromuconazole (Bectra SC, 100 g L–1) at four different concentration (0.01, 0.1, 1 and 10 μg mL–1). They were all tested individually by aseptically transfer the pathogen culture to the center of the solidified PDA medium in plates (90 mm diameter) with different concentrations of fungicides and a control plates contained only pathogen. The plates were incubated for five days at 25±2°C. The pathogen growth inhibition was observed, measured and compared with the growth of the pathogen in control.
Molecular study: In order to ascertain the identity of the conventionally identified pathogen, we carried out molecular characterization whereby we generated the sequences from PDA cultures and compared with available sequences in the GenBank.
DNA extraction, amplification and sequencing: Total DNA was extracted from the isolated pure culture of the pathogen following the protocol of the plant genomic DNA extraction kit (VIOGEN). Diluted (10–1-10–3) were used for PCR amplifications. The ef1 and ef2 primers very useful in a wide variety of filamentous ascomycetes as in Geiser et al.17 were used. The primer targeted to amplify an ~700 bp region of TEF which is over half of the amplicon’s length, in all known fusaria. The gene was chosen because it consistently appears as a single-copy in Fusarium, which shows a high, level of sequence polymorphism among its closely related species18. A standard Polymerase Chain Reaction (PCR) protocol was used to amplify the TEF gene region ef1 (forward primer: 5’-ATGGGTAAGGA(A/G)GACAAGAC-3) and ef2 (reverse primer: 5’-GGA (G/A)GTACCAGT(G/C)ATCATGTT-3’) designed by O'Donnell et al.19. Amplifying conditions were as detailed in Tibuhwa et al.20 with little modification. Primers were used in a PCR reaction with an annealing temperature of 53°C. Amplicons were visualized on 0.5% agarose gels, the DNA stained with 2.5 μL gel red while the marker used was lambda DNA ladder (20 ng μL–1) and the PCR product were purified using Millipore plates (MultiScreen™ PCR, Danvers, Massachusetts, USA). Sequencing, automated reaction clean up and visualization was carried out at Incaba in South Africa.
Alignments and phylogenetic analysis: The obtained Fusarium sequences were aligned using MAFFT version 6.847b21 on the online server (version 6), which were used to create alignments that utilized the L-INS-I MAFFT algorithm22. Two alignments were generated using the default settings (gap opening penalty = 1.53 and offset value = 0.00). For the analysis of the partial TEF region parsimonious tree (Fig. 5) was constructed using MEGA version 6 by Tamura et al.22. The sequences were analysed together with those found in the GenBank for comparison and ascertain the identity of the Fusarium isolates. After editing, the TEF sequence from sequenced isolates were copied and pasted into a web browser connected to the BLAST server and used as a query for comparison to the database. The nucleotide BLAST retrieved the closest matches to the query sequence and presented the matches as a series of DNA alignments with corresponding percentage match information. Taxa included in the phylogenetic analysis are as in Fig. 5 represented by names and their corresponding we continued GenBank accession number.
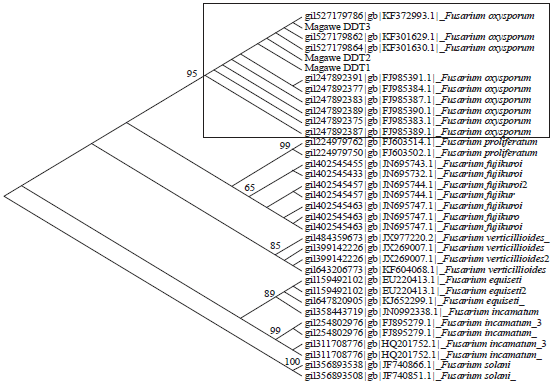 | |
| Fig. 5: | Phylogenetic tree using partial TEF region by neighbour joining method showing Fusarium oxysporum forming a monophyletic clade highlighted (Bootstrap support value above 50 is shown in each node) |
Moreover, the three Fusarium sequences generated in this study assigned collection number Tibuhwa D 01.2014, Tibuhwa D 02. 2014 and Tibuhwa D 03. 2014 were also included in the analysis.
Farmers observations: Interviews were conducted to the Magawa cashew farmers on the symptoms and the general observation of their cashew wilts. Farmers explained the disease symptom to appear over 3 years ago. We also monitored the symptoms for the 2 years 2014-2015 to ascertain some of the famer’s observations. The face-to-face interview as detailed by Tibuhwa23 was conducted among 25 farmers and the observations were for over 4 periods of rain seasons. The interviewees were stratified in the village to cover farms owners in three areas, which were severely affected, less affected and asymptomatic farms. The questionnaire also asked for any observation on the effect of the disease as related to other plants.
RESULTS AND DISCUSSION
The study shows that the cashew wilt in Magawa village is caused by a fungal pathogen. The study established that the pathogen was soil-borne and entered the cashew trees (host) through wounds that are caused naturally by the growth of young rootlets through the soil or cracks that appear at the sites of lateral root formation. It may also get access to the plant via wounds in older roots that are made during transplanting and cultivating; by root-feeding organisms, such as insects or nematodes and by man-made injuries during transplanting, cultivation, hand hoe weeding. This mechanism of Fusarium inversion to the cashew is similar to that observed in banana wilt disease caused by Fusarium species7 and in other vascular plants as noted by Genin24. Both macro-micromorphological characters and molecular analysis results obtained in this study show that this pathogen belongs to a genus Fusarium. The obtained pathogen sequences analysed with different other Fusarium sequences, largely confirm this as Fusarium oxysporum by forming a monophyletic clade highlighted in Fig. 5.
Pathogen characterization and identification
Morphological: Macroscopically the pathogen Fusarium oxysporum grows rapidly on PDA at 25°C±2 and produce spreading colonies which are flat cottony like. The colony colours are initially white with slightly pale yellowish margin observed from front while they look slightly dark when observed on reverse. With age, the culture turn purple-pinkish (Fig. 4a, b). Microscopically it possess hyaline septate hyphae, conidiophores which are relatively short with lateral branched and unbranched monophialides on which both micro and macroconidia are produced.
 | |
| Fig. 6(a-c): | (a) Cashew seedlings, (b) Inoculated seedlings and (c) Dying cashew seedling following inoculation. Symptoms and death occurred within 52-68 days, therefore, pathogenicity of tested isolate on cashew cultivars was clearly distinct and the same fungus species was isolated from the discoloured vascular tissue of diseased plants stem |
Macroconidia are typical fusiform, spindle shapedor slightly curved, pointed at the tip and mostly three septate. Chlamydospores appeared globose with relatively thick walls and they are either formed from hyphae or by the modification of hyphal cells (Fig. 4c).
Molecular study: Despite the fact that the exact sequence match with a known isolate in the database has been recently contended that sometime represent unambiguous species identification17, this study used both macro-micromorphological characters and approximately exact sequence match in the database to ascertain the identity of the studied pathogen. The pathogen isolates TEF sequences analysis reveals that it belongs to Fusarium genus as established conventionally. In Fig. 5 a phylogenetic tree places the pathogen together with Fusarium oxysporum clade (highlighted). These results together with the macro-micro-morphological supporting characters of culture colours, spindle shaped spores, places this pathogen together with Fusarium species. It thus taxonomically belongs to: Kingdom: Fungi, division: Ascomycota, class: Sordariomycetes, order: Hypocreales, family: Nectriaceae, genus: Fusarium, species: Fusarium oxysporum Schlecht. emend. Snyder and Hansen.
Pathogenicity test: Pathogenicity test of the isolate was confirmed on cashew seedlings with cuts by the diseases symptoms on infected plants. The seedling exhibited typical symptoms of Fusarium wilt as that noted in banana by Thangavelu and Mustaffa13. First indication of the disease was yellowing and drying of lower leaves which lead into general stunting of the plant, dark brown vascular discoloration and death unlike the control and undamaged seedling as shown in Fig. 6.
Anatomic study
Healthy plant (plant showing no symptoms): In healthy plants, we studied that the conducting tissues systems of both roots and shoots. The study on shoots showed that the conducting tissues (xylems and phloem) were normal (Fig. 7) and the pattern of conduction between the roots and leaves after injection of dyes and observing their movement indicated that in cashew tree a given root is directly connected to a particular set of branches. This explains the uneven chlorosis symptoms on different branches on mild affected tree (Fig. 2b). Both roots and stem were porous with large diameter xylem vessels (Fig. 7).
Diseased plant (plant showing symptoms): Anatomical study of the infested plants collected direct from the field showed that the effects of the pathogen are entirely on vascular tissues (conduction vessels). Cross section examination of both infested roots and stem tissues beneath the collar revealed pathogen mycelium and macroconidia in xylem vessels. The xylem vessels had bladder like growth (tyloses) protruding into lumen of tracheary elements. In most cases the xylems were completely blocked and cells growing in the vessels were typical parenchymatous which in severe cases made the whole system invisible and had brown appearance (Fig. 7a-c). The severely affected xylem in the infected cashew support the finding by Tjamos and Beckman25, Klosterman et al.26, Agrios27 and Genin24 who established that regardless of the mechanism used by vascular wilt pathogens to enter their hosts, they subsequently colonize the xylem vessels where they proliferate. From the anatomy of the diseased plant it shows that, the normal flow of liquids and nutrients from the roots to the foliage is greatly reduced or stopped because the conducting tissue becomes partially plugged or killed by excessive growth of pathogen mycelium, fungal spores or by the overgrowth of neighbouring cells.
 | |
| Fig. 7(a-c): | Cashew cross section (a and b) Stem and root cross section showing xylems and phloem tracheary elements in healthy cashew nut tree and (c) Cross section of root showing xylem blockage by parenchymatous cells in affected cashew nut roots |
These evidently interfere the translocation of nutrients by blocking their flow in phloem and xylem vascular bundles. This situation cause plant stress, leading them chlorosis, wilting and death, which at the same time their anatomy studies revealed the mycelia and spore blockage to the vascular bundle tissues (Fig. 7c). This thus, shows that they are the main causative agent of wilting of the trees. This study observation is in line with another finding by Moore et al.28 and Hermanto et al.29 who also noted wilting of tomato and banana by different Fusarium species through same mechanism of blocking the conducting tissues. It is well known that toxic substances are secreted by interaction of the pathogen and the host plant7,8. These materials apparently cause the wilting and eventual death of the plant. In line with this facts, this study observed plant exudes which were gums like thence sealed the vascular tissues in affected plants. Probably these exudes contained some chemical and toxins which might also contribute to wilting of the affected plant. It is well known that the growth of a plant is an integrated phenomenon that depends on a proper balance and functioning of all parts. If a large portion of the roots system is destroyed, a corresponding portion of the leaves and branches will die. Proper functioning of roots is as essential to the processes of photosynthesis as are the leaves and other chlorophyll-bearing parts of the plant30. In cashew nut plant, the fine feeder roots of the trees are connected to the leaves by an elaborate plumbing system consisting of larger transport roots, trunk, branch and twigs. Despite the fact that pattern of conduction between the roots and leaves of the trees varies between and within species, for ring-porous plant with large diameter xylem vessels like cashew nut, a given root is directly connected to a particular set of branches usually on the same side of the tree as the root30. This thus justify the obtained result in this study where by death or damage to the roots of the infested cashew (which are one-sided plumbing system) were observed to results in the death of the corresponding branches as opposed to those in which roots of the tree serve all of the branches and leaves.
Besides, during field study the physical observations identified that most of the cashew trees were undergoing chlorosis, dieback, wilting and death of the whole plant within few weeks of symptom appearances (Fig. 2a, b, 8a, c, d). Magawa area is located along the coast of Indian Ocean thus experience a very high temperature. Furthermore, the analyzed soil samples showed that it is sandy soil characterized with low chroma, which aggravate to high surface soil temperature11. The study also found that the cashew rooting system has five to seven root patterns originating from the "root collar" and grow horizontally through the soil. Their points of attachment to the tree trunk are usually at or near ground level and are associated with a marked swelling of the tree trunk (Fig. 8b). These major roots branches decrease in diameter over a distance 50-200 cm from the trunk to form an extensive netstudy of long rope-like roots.
In severely affected trees, cross section of transport roots showed very dark brown streaks in the vascular system indicating root internal injury. It was noted that when few roots less than 50% of the roots are affected the plant remain asymptomatic. Above 50% the symptom began usually on one side of the plant branch showing that probably few vascular tissues have been clogged with pathogen macroconidia. Since the pathogen sporulates fast in the collar part of the plant root (Fig. 8d), after the first symptom of chlorosis it takes a few months for the whole plant to undergo chlorosis and succumb to death by Agrios27 who established that most vascular wilt pathogens are confined to xylem vessels, some of them degrade xylem vessel walls to colonize adjacent parenchyma cells.
 | |
| Fig. 8(a-d): | Cashew tree (a) Dieback, (b) Rooting system, 5-7 root patterns originating from the "root collar" above which vascular tissues are completely unaffected, (c) Tap root cross sections and (d) Collar region of the root showing dark streak/spot due to internal injury |
It was interesting to note in the field that there were no new roots after injuries, which suggest that injury and death of roots might probably be caused by many agents. The study thus examined further for a progression of disease symptoms in plant exhibiting problems hence, examined all stages severely, moderately and non affected or asymptomatic plants (Fig. 2). The result showed that even asymptomatic plants have been infected by the pathogen only that the effect has not spread to over 50% of the roots. This thus envisage a thoroughly study to screen the spectrum of the effect in order to detain the spread to unaffected areas.
Culture sensitivity, soil analysis and field environmental parameters: The results for pathogen sensitivity test against five broad-spectrum antifungal chemicals showed that all had considerable inhibitory action with generally no big differences among them. Based on this result and the general fact that Fusarium oxysporum spores are freely found in the soil, this study recommends systemic antifungal chemicals in case of chemical method application. The systemic fungicides are taken up and redistributed through the xylem vessels and remain there for quite sometime thus enhance long resistance to pathogen attack29. For the analysis, results showed no salt accumulations in any profile with the electro conductivity ranging from 20-90 μS cm–1. Soil pH were normal and ranged from 5.6-6.3. Toxic heavy metals were in levels that could not be detected by AAS-IC p<0.0001 ppm.
With regard to field environmental parameters observed; Fusarium oxysporum is a well-known soil saprophyte with an ability to survive in most soil-arctic, tropical, desert and in both cultivated and non-cultivated fields for long periods of time. Observation in the field showed excess temperature of up to 32-35°C and high relative humidity of up to 76%, the parameters which are well known to trigger Fusarium into a soil-borne pathogen13,31. In soil, the optimum temperature for root infection is 30°C and above, while growth on artificial media is between 25 and 30°C.
Farmers observation and possible scientific enlightenment: It was reported from the Magawa farmers that ‘the disease worsens after the rain season’. This could best be explained by two main reasons:
| • | Disease and symptoms development of Fusarium oxysporum are extremely dependent upon air and soil temperatures. Symptoms are usually absent or mild at temperatures below 24°C and are most severe at constant temperatures of 32°C and above13. In the coast region where Magawa is found, usually the area experience low temperature during the rain season. During this time plants may be infected, yet show no symptoms until the temperature rises |
| • | During rain season water runoff and seepage play a big role of spreading the pathogens to unaffected plants such that after rains period, follow the period characterized by high temperature and humidity, which favours pathogen sporulation hence, symptoms become more evident |
| • | On the other hand, water is plenty during rain season where by even few roots which remain unaffected, could supply the nutrients to the plant. Visible symptoms may appear to be more severe due to restricted water movement in the plant brought about by damage done to the water conducting vessels earlier in the growing season |
It was presented and shown by the farmers that ‘one farm was singled out unaffected while surrounded by other affected farms’. This could be due to the following:
| • | The study carried out shows that the singled out unaffected farm is located in a relative lower part compared to others and that the water table in that farm is near the ground surface the fact proved by the natural water well found in that farm which serve as the source of water to their homes. Besides, the cashew tree analysed from that farm, although looked healthy and asymptomatic but yet they showed some few roots being affected hence with time the effect may accelerate to symptomatic |
| • | Furthermore, soil analysis showed that the soil physical structure in that farm; type of soil, drainage condition, revealed a relatively clay soils with high chroma which make it not be overheated compared to sandy soil thus disfavour the sporulation of the parasite which needs high temperature. This type of soil is referred to as "suppressive soil" which is very important in biological control of Fusarium wilt. In this soil, despite the presence of a virulent pathogen and susceptible host, disease either does not develop or the severity and spread of disease through the site is restricted32. Moreover, the suppression soil has generally been associated with nutritional status and microbial composition (Fungi, bacteria and actinomycetes) and biological factors which also play a major role in Fusarium wilt suppression as it was established by Scher and Baker33 and Alabouvette et al.34 but this was however beyond the scope of this study |
It was presented by the farmers that ‘once a symptom occurs the plant succumbs to death within a short time’. This could be due to the following:
| • | The affected plant usually can survive the pathogen and remain asymptomatic until 50% of the roots are affected. The symptoms become vivid when the effect goes beyond 50%. At this stage the pathogen mycelia and micro and macroconidia are so plenty in affected root and to the collar part. In response to this, the cashew tree secrete exudes which are gum/wax in nature, these exudes completely seals the vascular tissues which hasten the wilting of the plant due to complete blockage of nutrient transport |
It was observed and presented by the farmers that ‘only cashew tree wilt out amid other wild plants including domesticated plants such as mango trees, lemon trees etc’. This could be due to the following:
| • | The pathogen may even attack other plants but the anatomy of the cashew tree of having restricted, one-sided plumbing systems usually results in the death of the corresponding branches unlike others where roots of the tree serve all the branches and leaves, such that death or injury to one or some roots does not lead to one sided death in the crown of the tree. Thence, their concession to effects may take much longer time compared to cashew trees thus, their survival |
| • | Pathogenic isolates of Fusarium oxysporum often display a high degree of host specificity and can be subdivided into formal specialities on the basis of the plant species affected32-35 |
Recommendations on how to mitigate cashew Fusarium wilt disease: The emergence of these pathogenic fungi has to be tackled immediately and with great concern since the Magawa villagers depend heavily on cashew as their source of income hence their lives. In order to come-up with a rational mitigation measures, knowledge of the mechanisms by which the pathogen spread to cashew, especially exploring the presence of primary infester if any and how to control them as well as the spread of the pathogen spore to un affected land is crucial. Among the ways and practices, which may help in mitigating the problem are as follows:
| • | Farmers to understand better management practices by make themselves familiar with Fusarium wilt information material in order to recognise the disease symptoms and comply with all recommended mitigation measures. For example, in plantation where weeding is often done by hoeing, farmers have to use hands in order to avoid damaging the roots which might provide opportunity for the pathogen to get into the plant |
| • | Combined strategies used in other crops such as banana and cotton affected by the same disease including application of soil fumigation as detailed by Herbert and Marx36, fungicides described by Lakshmanan et al.37, crop rotation by Hwang38 and Su et al.39, flood -fallowing by Wardlaw40 and Stover and organic amendments by Stover41 might be applied. However, these methods study effectively in small areas while in large area the disease could be controlled effectively by planting of resistant cultivars28. For the case of the studied area Magawa and if at all the disease is still limited to the same place, other strategies that can be deployed in short and long-term measures to control the spread of the disease and eradicate the infection includes: |
| • | Fungicides as reported by Paul and Sharma42, flood furrows and burning affected plants To help restricting movement of contaminated soils and water flow within the infected areas. Adjusting the soil pH by liming since Fusarium pathogen usually flourishes well at pH range of 4 and 8. Increase the soil chroma to help temperature regulations thus minimizes soil overheating which favours the pathogen sporulation as well as the use of biological control although cashew Fusarium resistant varieties are not yet developed and the resistant varieties implementation usually faces challenges of consumer preference as it has been noted in other crops43. Nevertheless biological control of Fusarium wilt disease has become an increasingly popular disease management consideration in other crops because of its environmental friendly nature32,44-46. Under these circumstances, use of antagonistic microbes especially bacteria in genus Bacillus, which protect and promote plant growth by colonizing and multiplying in both rhizosphere and plant system could be the best potential alternative approach for the management of Fusarium wilt of cashew crop |
CONCLUSION
This is the first study to establish that Fusarium oxysporum species cause cashew wilt disease. The study thus, draws an attention to other cashew growers in the world of the emerging threat of Fusarium wilting diseases to cashew crop. To the studied area, it is important that the responsible authorities improvise and popularise the recommended strategies in order to prevent the spread of the disease toward its eradication.
SIGNIFICANCE STATEMENT
Through both conventionally and molecular characterization together with pathogenicity test, the result of this study for the first time renowned Fusarium oxysporum species as a pathogen causing cashew wilt disease. The study also provides scientific explanations to the farmer’s observations on the emerged unknown disease and provides recommendations on how to mitigate the problem.
ACKNOWLEDGMENT
This study was funded by Naliendele Agriculture Research Institute (NARI) which is highly acknowledged. The study also acknowledges the assistance of Mr. Charles Kweyunga of Botany department for technical assistance, the Department of Molecular Biology and Biotechnology of the University of Dar es Salaam for providing venue and most of the needed facilities throughout the study, the Magawa farmers for their cooperation throughout the study and moreover, the Rockefeller foundation that offered me a practitioner residency at Bellagio Centre, that availed me an ample time to finalise the manuscript.
REFERENCES
- Afshin, A, R. Micha, S. Khatibzadeh and D. Mozaffarian, 2014. Consumption of nuts and legumes and risk of incident ischemic heart disease, stroke and diabetes: A systematic review and meta-analysis. Am. J. Clin. Nutr., 100: 278-288.
CrossRefDirect Link - Sijaona, M.E.R. and J.W. Mansfieid, 1999. Structural resistance of cashew (Anacardium occidentaie) against powdery mildew (Oidium wulcardu). Tanzania J. Agric. Sci., 2: 115-125.
Direct Link - Zhou, B.J., P.S. Jia, F. Gao and H.S. Guo, 2012. Molecular characterization and functional analysis of a necrosis- and ethylene-inducing, protein-encoding gene family from Verticillium dahliae. Mol. Plant-Microbe Interact., 25: 964-975.
CrossRefDirect Link - Santhanam, P., H.P. van Esse, I. Albert, L. Faino, T. Nurnberger and B.P.H.J. Thomma, 2013. Evidence for functional diversification within a fungal NEP1-like protein family. Mol. Plant-Microbe Interact., 26: 278-286.
CrossRefDirect Link - Perez-Vicente, L., M.A. Dita and E. Martinez-de la Parte, 2014. Prevention and diagnostic of Fusarium wilt (Panama disease) of banana caused by Fusarium oxysporum f. sp. cubense Tropical Race 4 (TR4). Technical Manual Prepared for the Regional Training Workshop on the Diagnosis of Fusarium wilt in St. Augustine, Trinidad and Tobago, May 2014.
- Leslie, J.F. and B.A. Summerell, 2006. The Fusarium Laboratory Manual. Blackwell Publishing, Hoboken, New Jersey, ISBN-13: 9780813819198, Pages: 388.
CrossRefDirect Link - Nelson, P.E., T.A. Toussoun and W.F.O. Marasas, 1983. Fusarium Species: An Illustrated Manual for Identification. Pennsylvania State University Press, Philadelphia, Pennsylvania, ISBN: 9780271003498, Pages: 193.
Direct Link - Thangavelu, R. and M.M. Mustaffa, 2012. Current Advances in the Fusarium Wilt Disease Management in Banana with Emphasis on Biological Control. In: Plant Pathology, Cumagun, C.J. (Ed.). Chapter 11, InTech Publisher, Rijeka, Croatia, ISBN 978-953-51-0489-6, pp: 273-298.
Direct Link - Al-Juboory, H.H. and K.S. Juber, 2013. Efficiency of some inoculation methods of Fusarium proliferatum and F. verticilloides on the systemic infection and seed transmission on maize under field conditions. Agric. Biol. J. North Am., 4: 583-589.
Direct Link - Tijjani, N., I.J. Dioha, B. Alhassan, A.I. Eleri, A.M. Lawal and I. Muhammad, 2013. Determination of some heavy metals in soil samples obtained from Rimi local government in Katsina State, Nigeria. Chem. Mater. Res., 3: 27-30.
Direct Link - Geiser, D.M., M. del Mar Jimenez-Gasco, S.C. Kang, I. Makalowska and N. Veeraraghavan et al., 2004. FUSARIUM-ID v. 1.0: A DNA sequence database for identifying Fusarium. Eur. J. Plant Pathol., 110: 473-479.
CrossRefDirect Link - Cianchetta, A.N., T.W. Allen, R.B. Hutmacher, R.C. Kemerait and T.L. Kirkpatrick et al., 2015. Survey of Fusarium oxysporum f. sp. vasinfectum in the United States. J. Cotton Sci., 19: 328-336.
Direct Link - O'Donnell, K., E. Cigelnik and H.I. Nirenberg, 1998. Molecular systematics and phylogeography of the Gibberella fujikuroi species complex. Mycologia, 90: 465-493.
CrossRefDirect Link - Tibuhwa, D.D., S. Savic, L. Tibell and A.K. Kivaisi, 2012. Afrocantharellus gen. stat. nov. is part of a rich diversity of African Cantharellaceae. IMA Fungus, 3: 25-38.
Direct Link - Katoh, K., K.I. Kuma, H. Toh and T. Miyata, 2005. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res., 33: 511-518.
CrossRefDirect Link - Tamura, K., G. Stecher, D. Peterson, A. Filipski and S. Kumar, 2013. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol., 30: 2725-2729.
CrossRefPubMedDirect Link - Tibuhwa, D.D., 2013. Wild mushroom-an underutilized healthy food resource and income generator: Experience from Tanzania rural areas. J. Ethnobiol. Ethnomed., Vol. 9.
CrossRefDirect Link - Genin, S., 2010. Molecular traits controlling host range and adaptation to plants in Ralstonia solanacearum. New Phytol., 187: 920-928.
CrossRefDirect Link - Klosterman, S.J., K.V. Subbarao, S. Kang, P. Veronese and S.E. Gold et al., 2011. Comparative genomics yields insights into niche adaptation of plant vascular wilt Pathogens. PLoS Pathog., Vol. 7.
CrossRef - Agrios, G.N., 2005. Plant Pathology. 5th Edn., Elsevier, Cambridge, Massachusetts, ISBN-13: 9780080473789, Pages: 922.
Direct Link - Perry, T.O., 1982. The ecology of tree roots and the practical significance thereof. J. Arboricult., 8: 197-211.
Direct Link - Lazarotto, M., R. Mezzomo, C.G. Maciel, G. Finger and M.F.B. Muniz, 2014. Mycelia growth and sporulation of Fusarium chlamydosporum species complex under different culture conditions. Amazonian J. Agric. Environ. Sci., 57: 35-40.
CrossRefDirect Link - Scher, F.M. and R. Baker, 1982. Effect of Pseudomonas putida and a synthetic iron chelator on induction of soil suppressiveness to Fusarium wilt pathogens. Phytopathology, 72: 1567-1573.
CrossRefDirect Link - Alabouvette, C., P. Lemanceau and C. Steinberg, 1993. Recent advances in the biological control of fusarium wilts. Pestic. Sci., 37: 365-373.
CrossRefDirect Link - Castano, R., B. Scherm and M. Aviles, 2014. Genetic diversity of Fusarium oxysporum f. sp. dianthi in Southern Spain. J. Mycol.
CrossRefDirect Link - Herbert, J.A. and D. Marx, 1990. Short-term control of Panama disease of bananas in South Africa. Phytophylactica, 22: 339-340.
Direct Link - Lakshmanan, P., P. Selvaraj and S. Mohan, 1987. Efficacy of different methods for the control of Panama disease. Trop. Pest Manage., 33: 373-374.
CrossRefDirect Link - Su, H.J., S.C. Hwang and W.H. Ko, 1986. Fusarial wilt of Cavendish bananas in Taiwan. Plant Dis., 70: 814-818.
CrossRefDirect Link - Paul, P.K. and P.D. Sharma, 2002. Azadirachta indica leaf extract induces resistance in barley against leaf stripe disease. Physiol. Mol. Plant Pathol., 61: 3-13.
CrossRefDirect Link - Viljoen, A., 2002. The status of Fusarium wilt (Panama disease) of banana in South Africa. South Afr. J. Sci., 98: 341-344.
Direct Link - Marois, J.J., D.J. Mitchell and R.M. Sonoda, 1981. Biological control of fusarium crown rot of tomato under field conditions. Phytopathology, 71: 1257-1260.
Direct Link - Larkin, R.P. and D.R. Fravel, 1998. Efficacy of various fungal and bacterial biocontrol organisms for control of Fusarium wilt of tomato. Plant Dis., 82: 1022-1028.
CrossRefDirect Link - Thangavelu, R., A. Palaniswami and R. Velazhahan, 2004. Mass production of Trichoderma harzianum for managing fusarium wilt of banana. Agric. Ecosyst. Environ., 103: 259-263.
CrossRefDirect Link