
Noor Andryan Ilsan
Graduate School, Departement of Biology, Faculty of Mathematics and Natural Science, Bogor Agricultural University, Darmaga, Bogor 16680, West Java, Indonesia
Abdjad Asih Nawangsih
Department of Plant Protection, Faculty of Agriculture, Bogor Agricultural University, Darmaga, Bogor 16680, West Java, Indonesia
Aris Tri Wahyudi
Graduate School, Departement of Biology, Faculty of Mathematics and Natural Science, Bogor Agricultural University, Darmaga, Bogor 16680, West Java, Indonesia
Asian Journal of Plant Pathology
Year: 2016 | Volume: 10 | Issue: 1-2 | Page No.: 1-8







ABSTRACT
Bacterial Leaf Blight (BLB) caused by Xanthomonas oryzae pv. oryzae (Xoo) is the most destructive and serious disease causing productivity loss, especially in rice, in Asia. The chemical pesticide usage over a long period of time brings about adverse toxic effect on the potential production of the land and the main consumers of the products. Microbes as biocontrol agents have efficiency and safety for humans and other non-target organisms. They leave small amount or no residue in foods. Soil microbes cannot be used directly as biopesticides because common root-colonizers fail to establish on leaves. Actinomycetes are Gram positive bacteria that are known to produce bioactive compound up to 70% of the total compound produced by bacteria, including antimicrobes. Several strains of actinomycetes are known capable of protecting plants against plant disease. The aims of this study were to isolate and screen non-pathogenic phyllosphere actinomycetes of rice which are capable of controlling BLB disease in rice. A total eight isolates are positively capable of controlling Xoo in vitro and four isolates significantly reduce disease severity of BLB. Pellet of STG 15, which is the best, showed 25.87% control efficiency of BLB severity at 14 Days After Inoculation (DAI) using Xoo compared to control. Molecular identification based on 16S rRNA gene performed for six isolates with the highest activity showed that they belongs to genus Streptomyces, Actinomadura, Nonomuraea, Micromonospora. Isolate STG 15 which has the highest capability to control BLB incidenceis identified as Nonomuraea sp.
PDF Abstract XML References Citation
Received: October 23, 2015;
Accepted: December 28, 2015;
Published: March 15, 2016
How to cite this article
Noor Andryan Ilsan, Abdjad Asih Nawangsih and Aris Tri Wahyudi, 2016. Rice Phyllosphere Actinomycetes as Biocontrol Agent of Bacterial Leaf Blight Disease on Rice. Asian Journal of Plant Pathology, 10: 1-8.
URL: https://scialert.net/abstract/?doi=ajppaj.2016.1.8
URL: https://scialert.net/abstract/?doi=ajppaj.2016.1.8
INTRODUCTION
Rice is a staple food for Asian, especially Indonesia. National rice production in 2014 reached 70.61 million tons of grains. This production has decreased by 0.67 million tons (0.94%) compared to 2013 (BPS., 2014). Bacterial Leaf Blight (BLB) disease caused by Xanthomonas oryzae pv. oryzae (Xoo) is one of the common causes of rice production decrease in some regions of Asia. Several rice plant areas experience 10-20% productivity loss because of BLB in moderate condition and even up to 50% in optimum condition. The intensity of the disease increases during rainy season because Xoo enters plants through natural openings such as hydathode, stomata or wound (Mew et al., 1993).
The prevention for BLB disease caused by Xoo is a serious problem. The chemical pesticide usage over a long period of time brings about adverse toxic effect on the potential production of the land and the main consumers of the products (Kumari et al., 2014). Microbes as biocontrol agents have efficiency and safety for humans and the other non-target organisms. They leave small amount or no residue in foods. It is reported that some biocontrol microbes can reduce the use of agricultural chemical compounds. The sources of biocontrol microbes for plant pathogen are among others soil (Hastuti et al., 2012) or phyllosphere (Wang and Ma, 2011). Most of the biocontrol microbes from soil cannot be used as a control agent directly. The difference between the composition of bacterial community in leaves and roots is shown by the failure of roots colonization such as Rhizobium and Azospirillum to colonize leaves (O’Brien and Lindow, 1989). Commonly, parts of the plant above the ground normally contain a variety of bacteria, yeasts and fungi. Only few microbes can be isolated from plant tissues and mostly they are from the surface of healthy plants. Aerial parts of plants colonized by microbes are known as phyllosphere; while the colonizing microbes are known as epiphytic microbes. Phyllosphere microbiology focuses on leaves. Leaf microbial communities consist of various genera of bacteria, filamentous fungi, yeasts, algae and several protozoa and nematodes. Bacteria are the most important microbes living in phyllosphere (Lindow and Brandl, 2003).
Actinomycete is a Gram positive bacteria that are known producing a variety of bioactive compounds up to 70% of the total bioactive compounds produced by bacteria (Takahashi, 2004). Actinomycete is capable of producing a variety of secondary metabolites with different biological functions such as antimicrobes, enzyme inhibitors and enzymes that degrade organic matter (Emmert and Handelsman, 1999). Several strains of actinomycetes protect plants against plant disease (Minuto et al., 2006; Boukaew and Prasertsan, 2014). Several study has reported phyllosphere actinomycete as biocontrol agent. Previous study reported that actinomycete in cucumber’s phyllosphere has fungistatic activity, in addition to the culture filtrate of which is capable of inhibiting the spore germination of fungal plant pathogen species Corynespora cassiicola (Wang and Ma, 2011). The main purpose of this study was to isolate and screen phyllosphere actinomycetes from rice plants as biocontrol agents to control BLB caused by Xoo in greenhouse.
MATERIALS AND METHODS
Isolation of rice phyllosphere actinomycetes: Rice plants were collected from Situgede rice fields (Bogor, Indonesia) during dry season. Rice plants with healthy leaves were collected from rice field that showed BLB symptoms. The isolation of phyllosphere actinomycetes were carried out using washing method. A total of 10 g of rice plants’ healthy leaves in vegetative phase (±1 month), ripening phase (±2 month) and flowering phase (±2.5 month) were each cut into small pieces. The leaves were then given pre-treatments, i.e., heat shock treatment at 70°C for 15 min prior to homogenization in 90 mL water saline for 1 h. The solution was then diluted in 10–1-10–4 series on Humic Acid Vitamin (HV) agar medium added with nalidixid acid (Sigma Aldrich) and cycloheximide (Sigma Aldrich) of 50 ppm each.
Anti-Xoo assay: Primary screening was performed using plug agar method (Takahashi, 2004). The Xoo strain 8004 isolate were provided by Dr. Alina Akhdiya from Indonesian Center for Agricultural Biotechnology and Genetic Research and Development (ICABIOGRAD) Cimanggu, Bogor, Indonesia. Actinomycete isolates were inoculated on Yeast Malt (YM) agar plates and were incubated for 7 days. Agar plugs with 6 mm diameter were taken from incubated cultures on agar using sterile borer and put onto the surface of Luria Agar (LA) containing Xoo (1% Xoo with 107 CFU mL–1 culture density) prior to incubation at 37 C for 24 h. Anti-Xoo activity is showed by inhibition zone. Experiments were carried out in three replications.
Hypersensitivity Response (HR) and pathogenicity test: Selected phyllosphere actinomycetes with anti-Xoo activity were grown in YM broth (±106 CFU mL–1) and injected into tobacco leaves using needleless syringe prior inoculation into one month-susceptible rice plants (IR 64) using leaf clipping method (Krishanti et al., 2015). Plant response was examined at 48 h for Its hypersensitivity and 14 Days After Inoculation (DAI) for pathogenicity. The Xoo wild type was used as positive control and E. coli DH5α was used as negative control. Experiments were carried out in three replications.
Molecular identification based on 16S rRNA gene: Bacterial genomic DNA was extracted using bacterial genome isolation (Genomic DNA Mini Kit for Blood/Cultured Cell, Geneaid). The 16S rRNA gene sequences were amplified using universal primer 16S rRNA gene for bacterial domain 63f (5’-CAG GCC TAA CAC ATG CAA GTC-3’) and 1387r (5’-GGG CGG WGT GTA CAA GGC-3’) (Marchesi et al., 1998) with amplicon targets length of ~1300 bp. Polymerase Chain Reaction (PCR) mixture for 25 μL total volume consisted of 12.5 μL Go Taq Green Master Mix (Promega, USA), 2 μL reverse primer and forward each, 4 μL DNA and 4.5 μL nuclease free water. The PCR (using SwiftTM Mini Pro Thermal Cycler Esco, Singapore) conditions began with initial denaturation for 4 min at 94°C, followed with 30 cycles of denaturation process for 30 sec at 94°C, annealing for 30 sec at 55°C, elongation for 1 min at 72°C and final elongation for 7 min at 72°C. The PCR products were sequenced, analyzed using BLAST and the phylogenetic tree of which was constructed using Mega 5.2 program.
Preparation of biocontrol application in greenhouse: Isolates were mutated by using rifampicin (Sigma Aldrich) up to 75 μg mL–1 in concentration (Wahyudi et al., 2011). A total of two formulations were used in greenhouse application, i.e., pellet and supernatant. Pellet and supernatant formulation was prepared by growing the isolates in 100 mL YM/MNG broth media for 7 days at 37°C on rotary shaker at 150 rpm. The pellet was harvested using centrifugation at 3820 g for 20 min. Pellet and supernatant were mixed in ratio 1:1 and 20 mL of this mixture was added with 1% gum Arabic (Hastuti et al., 2012). Supernatant formulation was prepared by separating the pellet using centrifugation at 3820 g for 20 min (Boukaew and Prasertsan, 2014).
The Xoo inoculum was prepared 100 mL LB medium and incubated in shaker incubator at 150 rpm for 2 days at 37°C. Xoo Inoculum was used at density of 107-108 CFU mL–1. Infection of Xoo was done using clipping method. The leaves were cut by scissor which had been dipped in Xoo culture of 14 Days After Planting (DAP). Rice seeds cultivar IR 64 (susceptible to Xoo) were surface-sterilized in 2% sodium hypochlorite for 2 min, rinsed three times in sterile distilled water and left dipped in sterile distilled water for 1 day for imbibitions prior to germination experiment. Germinated seedlings were cultivated in paddy soil with nursery plastic baskets in greenhouse. After 15 days, rice seedlings were relocated onto pots (of 25 cm diameter×25 cm height containing 3 kg of paddy soil) with three rice plants each. A total of 30 g compost fertilizer was applied each pot.
Application of biocontrol in greenhouse experiment: A total of six actinomycete isolates were selected (STG 11, STG 8, STG 15, STG 1, STG 2 and STG 4) as biocontrol of Xoo. Positive control was only inoculated with Xoo. Negative control was cut without Xoo inoculation. Each treatment was carried out in three replications. Inoculating pellet (pellet was inoculated by greasing the leaves using sterile paint brush) and supernatant spraying was conducted every week, started at 7 until 21 DAP. Infection of Xoo was carried out using clipping method at 24 DAP. BLB disease visual assessment was determined 14 (DAI) of Xoo (38 DAP). The disease incidence and control efficiency were calculated using the following formulae (Gnanamanickam et al., 1999):
![]()
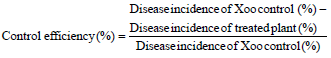
The data were subjected to analysis of variance (ANOVA) using SPSS software version 21 for windows. Statistical significance was evaluated using Duncan’s Multiple Range Test (DMRT) and p<0.05 was considered significantly different.
RESULTS
Rice phyllosphere actinomycete Isolation and anti-Xoo assay: A Total of 22 isolates were successfully obtained, 17 of which were from leaves in vegetative phase, 4 from flowering phase and 1 from ripening phase (Fig. 1). A total of 8 isolates were capable of inhibiting Xoo using plug agar method in LA medium (Fig. 2 and Table 1). All isolates that were capable of inhibiting Xoo were from leaves in vegetative phase.
Hipersensitivity and pathogenecity test: Biocontrol agent was selected using hypersensitivity test on tobacco leaves and pathogenecity assay on rice plant (IR 64).
| Table 1: | Primary screening of isolates against Xoo using plug agar method |
 | |
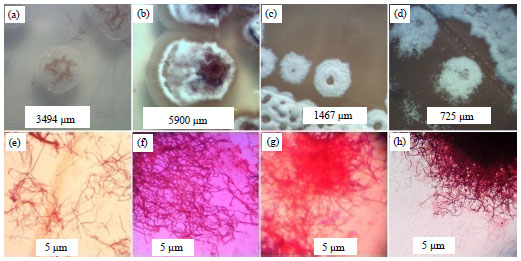 | |
| Fig. 1(a-h): | Morphology of isolates on YM agar medium at 14 days incubation and Gram staining of isolates, (a, e) STG 11, (b, f) STG 15, (c, g) STG 4 and (d, h) STG 8 |
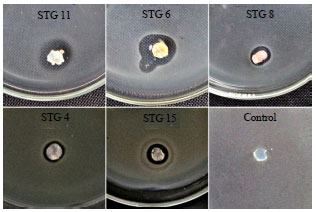 | |
| Fig. 2: | Primary screening against Xoo using plug agar method |
 | |
| Fig. 3(a-h): | HR and pathogenecity test, (a) E. coli DH5α as negative control in HR test, (b) Xoo as positive control, (c) Negative response of STG 8, (d) Positive response of STG 6, (e) Xoo as positive control in pathogenecity test, (f) E. coli DH5α as negative control, (g) Positive response of STG 16 and (h) Negative response of STG 11 |
Hypersensitivity and pathogenecity reaction result showed that 6 of 8 selected isolates had negative response, indicated by no disease symptoms occurred (Fig. 3). The Xoo wild type was used as positive control while E. coli DH5α as the negative control. One month-old rice plants was inoculated with isolates. Xoo wild type inoculation showed the presence of bacterial leaf blight disease symptoms which were observed at 14 DAI. A total of 6 non-phytopathogenic isolates are potential as biocontrol agents against Xoo.
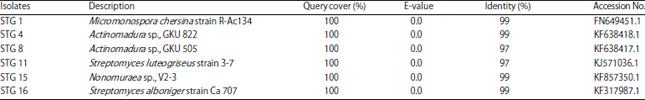
Molecular taxonomy of isolates as biocontrol agents: The 16S rRNA gene of 6 selected isolates of rice phyllosphere actinomycetes with DNA fragment estimated size of ~1300 bp were compared with 16S rRNA gene sequences in GenBank database. The STG 1 is related closely to Micromonospora chersina strain R-Ac134 with 99% maximum identity, STG 4 and STG 8 are to Actinomadura sp., with 99 and 97% maximum identity, respectively, STG 15 is to Nonomuraea sp., V2-3 with 99% maximum identity, STG 11 is to Streptomyces luteogriseus strain 3-7 with 97% maximum identity and STG 16 is to Streptomyces alboniger strain Ca 707 with 99% maximum identity (Table 2). The phylogenetic dendrogram showed that STG 1, STG 4, STG 8 and STG 15 fell in the same cluster (cluster I) while, STG 11 and STG 15 in cluster II (Fig. 4). Both cluster I and cluster II were separated from outgroup cluster (X. oryzae).
Application of biocontrol in greenhouse experiment: Response was observed at 14 DAI of Xoo (38 DAP) and the parameters observed were disease severity, plant height and number of tillers. The result of isolate application showed that formulations of pellet STG 2, STG 4, STG 8, STG 11, STG 15 and formulations of supernatant STG 11 and STG 8 were significantly capable of suppressing BLB disease severity compared to Xoo control (Table 3). Pellet STG 15, which is the best out of all, controlled BLB severity at 25.87% efficiency (necrotic symptom up to 62.23%) compared to Xoo control (necrotic symptom up to 88%).
| Table 2: | BLAST result of 16S rRNA gene sequences |
 | |
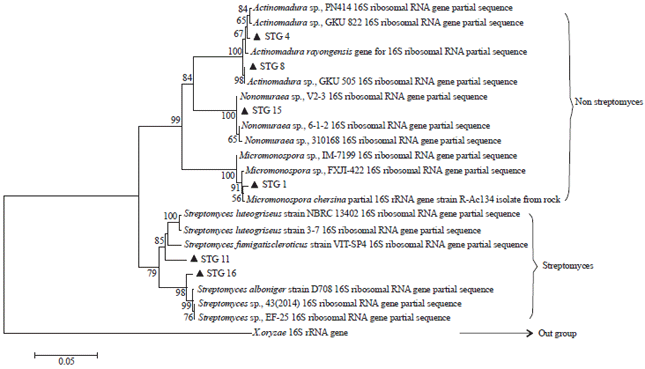 | |
| Fig. 4: | Phylogenetic tree based on the 16S rRNA gene of 6 isolates of rice phyllosphere actinomycetes. Scale indicates distance evolution on branch length while the numbers on the branches indicate bootstrap values |
| Table 3: | Result of plant response at 14 DAI of Xoo (38 DAP) |
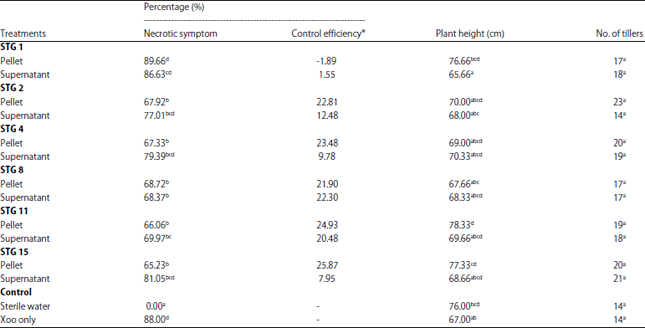 | |
| * Control efficiency compared to Xoo control, values in columns followed by same letter were not significantly different according to MDRT test (p<0.05 for necrotic symptom and p>0.05 for plant height and number of tillers) | |
Reisolation result showed that single colony of STG 15 in 10–4dilution, single colony of STG 2 in 10–3 dilution and single colony of STG 8 in 10–3 dilution.
DISCUSSION
The discovery and development of biopesticide to prevent plant disease caused by pathogen are important steps to reduce crop losses. In addition, biopesticide also has positive effects on the environment compared to chemical pesticide. The discovery of microbial biocontrol agent against plant pathogens focuses more on soil microbes, including actinomycetes. Actinomycetes are capable of producing a variety of secondary metabolites with different biological functions such as antimicrobial, enzyme inhibitors and enzymes that degrade organic matter (Emmert and Handelsman, 1999). Several strains of actinomycetes have been known suppressing plant diseases caused by pathogen. Streptomyces griseoviridis has been commercialized as biopesticides (MycostopTM) (Minuto et al., 2006). References underlay how actinomycetes, especially from phyllosphere, were discovered can be used as biopesticides directly. To the best of our knowledge, not many studies on direct actinomycetes isolation from phyllosphere have been carried out.
A total of 22 isolates were obtained from samples during dry season. Based on morphological characteristics, most of them were belong to non-Streptomyces group. The characterization of non-Streptomyces spp., species is mainly based on their substrate mycelia color, soluble pigment production and the shape (Ghadin et al., 2008). The highest number of actinomycete isolates was from rice leaves in vegetative phase (17 isolates), followed with in ripening phase (4 isolates) and flowering phase (1 isolate). Several studies reported the differences of the number of bacteria during different leaf ages (Krishanti et al., 2015). For phyllosphere, it is well known that environmental factors (radiation, pollution and nitrogen fertilization) as well as biotic factors (such as leaf age and the presence of other microorganisms) affect bacterial community structure. The results of an MLTreeMap analysis from four different phyllosphere samples: Glycine max (soybean), Trifolium repens (white clover), Arabidopsis thaliana, Oryza sativa (rice) showed that proteobacteri was the most represent phylum, with members of the class Alphaproteobacteria constituting up to -70% of the community in each of the four phyllosphere samples. Interestingly, the class Actinobacteria (including actinomycete) accounted for ~40% of the rice community but constituted a lower percentage of other samples (Vorholt, 2012).
Hipersensitivity Reaction (HR) and pathogenicity tests were carried out to determine the possibility of isolates pathogenic for rice plant. The HR is cell death area that formed at the entrance point of a pathogen and correlates to resistance exhibition (Mur et al., 2008). Positive response is when necrosis on leaves appears. A total of 6 isolates showed negative response for HR and pathogenicity tests. Those 6 isolates are recommended as biocontrol agents against Xoo.
Primary screening of anti-Xoo assay using plug agar indicates that two best isolates were STG 11 and STG 15 (Table 1). Two isolates that are the highest fast-growing isolates are STG 4 in YM broth medium (5.4 g L–1 7 days–1) and STG 15 in MNG broth medium (2.05 g L–1 7 days–1). The growth of isolates in YM and MNG broth media was observed by measuring dry weight. While on biocontrol application in greenhouse, pellet treatment of STG 15 and STG 11 showed the highest control efficiency (25.87 and 24.93%, respectively) compared to Xoo control. This data indicates that there is a connectedness between the application response test to suppress BLB disease severity and both in vitro assay (Hastuti et al., 2012) and isolates growth rate. Field test that conducted by Hastuti et al. (2012) using endophitic actinomycetes in suppressing BLB disease up to 70 days after planting showed a decrease of BLB symptom compared to control, however the differences was not significant. Based on statistical analysis, the treatments brought about no significant effect on plant height and number of tillers (p>0.05). Treatments also did not capable of inhibiting plant growth, seen from the response of plant height and number of tillers.
Based on 16S rRNA molecular identification on isolates, two of six biocontrol agents were identified belong to Streptomyces which is a known antibiotics producer genera and even serves as commercial antifungal biopesticides (Minuto et al., 2006). Antibiosis process may be involved and play an important role in suppressing plant disease by certain bacteria and fungi. The process is defined as the interactions involving compounds with low-molecular weight or antibiotics produced by microorganisms which directly affect other microorganisms (Weller, 1988). Nonomuraea and Actinomadura genera are groups of rare actinomycetes producing antimicrobial (Technikova-Dobrova et al., 2004; Nakaew et al., 2012). The STG 15 which was identified belongs to Nonomuraea is the best isolate in suppressing disease severity. Polyketide synthase I (PKS I) gene sequence of STG 15 relates closely to Polyketide synthase Kitasatospora azatica. Primer of PKS I and amplification condition references from Ayuso-Sacido and Genilloud (2005). Polyketides are a class of structurally diverse natural products which posses a wide range of biological activities (Hopwood, 1997). These compounds are used throughout medicine and agriculture as antimicrobials, immunosuppressants, antiparasitics and anticancer agents.
Potential organisms that previously successful during in vitro screenings were generally not so successful in vivo, i.e., in greenhouse or field. In vitro tests do not involve environmental and microbiological factors a particular biocontrol agent has to interact to in the field (Saika, 2008). Today, many people view that biopesticides should be applied using existing spray technologies but the success is highly limited because of improper equipment, poor formulation efficacy, or combination of both (Navon, 2000). Leaf surface has long been considered a hostile environment for bacterial colonies as it is exposed to rapidly fluctuating temperature and relative humidity. The leaf also provides limited nutrient resources to bacterial colonies (Lindow and Brandl, 2003). Most of phyllosphere bacterial colonies are easily killed by non-penetrating agents such as peroxide or UV light. Therefore, further studies are necessary to develop appropriate formulation to maintain the survival of biocontrol agent on the leaves.
CONCLUSION
A Total of 22 isolates of rice phyllosphere actinomycetes were obtained and based on morphological characteristics, most of them belong to non-Streptomyces group. A total of eight isolates out of which have anti-Xoo activity. Hypersensitivity and pathogenecity tests indicates that 6 isolates are not pathogenic for rice plant and five isolates out of which are significantly capable of suppressing BLB disease severity, through bioactive compounds that they have. Biocontrol application of potential isolates in greenhouse showed that pellet STG 2, STG 4, STG 8, STG 11 and STG 15, as well as supernatant STG 11 and STG 8 are significantly capable of suppressing BLB disease severity compared to Xoo control. Based on 16S rRNA identification, STG 4 is related closely to Actinomadura strain GKU 822, STG 8 is to Actinomadura strain GKU 505, STG 11 is to Streptomyces luteogriseus strain 3-7 and STG 15 is to Nonomuraea sp., V2-3. Based on detection of PKS I gene, STG 15 isolate is related closely to PKS Kitasatospora azatica.
ACKNOWLEDGMENT
This study was supported by research project from Partnership Cooperation Program of the National Agricultural Research and Development (KKP3N) of Indonesian Ministry of Agriculture that was granted to Prof Aris Tri Wahyudi. Therefore, we are grateful for this funding and support of this research.
REFERENCES
- Ayuso-Sacido, A. and O. Genilloud, 2005. New PCR primers for the screening of NRPS and PKS-I systems in actinomycetes: Detection and distribution of these biosynthetic gene sequences in major taxonomic groups. Microb. Ecol., 49: 10-24.
CrossRefDirect Link - Boukaew, S. and P. Prasertsan, 2014. Suppression of rice sheath blight disease using a heat stable culture filtrate from Streptomyces philanthi RM-1-138. Crop Protect., 61: 1-10.
CrossRefDirect Link - Technikova-Dobrova, Z., F. Damiano, S.M. Tredici, G. Vigliotta and R. di Summa et al., 2004. Design of mineral medium for growth of Actinomadura sp. ATCC 39727, producer of the glycopeptide A40926: Effects of calcium ions and nitrogen sources. Applied Microbiol. Biotechnol., 65: 671-677.
CrossRefDirect Link - Emmert, E.A.B. and J. Handelsman, 1999. Biocontrol of plant disease: A (Gram-) positive perspective. FEMS Microbiol. Lett., 171: 1-9.
CrossRefPubMedDirect Link - Ghadin, N., N.M. Zin, V. Sabaratnam, N. Badya, D.F. Basri, H.H. Lian and N.M. Sidik, 2008. Isolation and characterization of a novel endophytic Streptomyces SUK 06 with antimicrobial activity from Malaysian plant. Asian J. Plant Sci., 7: 189-194.
CrossRefDirect Link - Gnanamanickam, S.S., V.B. Priyadarisini, N.N. Narayanan, P. Vasudevan and S. Kavitha, 1999. An overview of bacterial blight disease of rice and strategies for its management. Curr. Sci., 77: 1435-1444.
Direct Link - Hastuti, R.D., Y. Lestari, R. Saraswati, A. Suwanto and Chaerani, 2012. Capability of Streptomyces spp. in controlling bacterial leaf blight disease in rice plants. Am. J. Agric. Biol. Sci., 7: 217-223.
CrossRefDirect Link - Hopwood, D.A., 1997. Genetic contributions to understanding polyketide synthases. Chem. Rev., 97: 2465-2498.
CrossRefDirect Link - Krishanti, N.P.R.A., A.T. Wahyudi and A.A. Nawangsih, 2015. Non-pathogenic phyllosphere bacteria producing bioactive compounds as biological control of Xanthomonas oryzae pv oryzae. Int. J. Pharma Bio Sci., 6: 801-810.
Direct Link - Kumari, K.A., K.N.R. Kumar and C.N. Rao, 2014. Adverses effect of chemical fertilizers and pesticides on human health and environment. J. Chem. Pharmaceut. Sci., 3: 150-151.
Direct Link - Lindow, S.E. and M.T. Brandl, 2003. Microbiology of the phyllosphere. Applied Environ. Microbiol., 69: 1875-1883.
CrossRefPubMedDirect Link - Marchesi, J.R., T. Sato, A.J. Weightman, T.A. Martin, J.C. Fry, S.J. Hiom and W.G. Wade, 1998. Design and evaluation of useful bacterium-specific PCR primers that amplify genes coding for bacterial 16S rRNA. Applied Environ. Microbiol., 64: 795-799.
Direct Link - Mew, T.W., A.M. Alvarez, J.E. Leach and J. Swings, 1993. Focus on bacterial blight of rice. Plant Dis., 77: 5-12.
Direct Link - Minuto, A., D. Spadaro, A. Garibaldi and M.L. Gullino, 2006. Control of soilborne pathogens of tomato using a commercial formulation of Streptomyces griseoviridis and solarization. Crop Prot., 25: 468-475.
CrossRefDirect Link - Mur, L.A.J., P. Kenton, A.J. Lloyd, H. Ougham and E. Prats, 2008. The hypersensitive response; the centenary is upon us but how much do we know? J. Exp. Bot., 59: 501-520.
CrossRefPubMedDirect Link - Nakaew, N., R. Sungthong, A. Yokota and S. Lumyong, 2012. Nonomuraea monospora sp. nov., an actinomycete isolated from cave soil in Thailand, and emended description of the genus Nonomuraea. Int. J. Syst. Evol. Microbiol., 62: 3007-3012.
CrossRefPubMedDirect Link - Navon, A., 2000. Bacillus thuringiensis insecticides in crop protection-reality and prospects. Crop Protect., 19: 669-676.
CrossRefDirect Link - O'Brien, R.D. and S.E. Lindow, 1989. Effect of plant species and environmental conditions on epiphytic population sizes of Pseudomonas syringae and other bacteria. Phytopathology, 79: 619-627.
CrossRefDirect Link - Takahashi, Y., 2004. Exploitation of new microbial resources for bioactive compounds and discovery of new actinomycetes. Actinomycetologica, 18: 54-61.
CrossRefDirect Link - Vorholt, J.A., 2012. Microbial life in the phyllosphere. Nat. Rev. Microbiol., 10: 828-840.
CrossRefDirect Link - Wahyudi, A.T., S. Meliah and A.A. Nawangsih, 2011. Xanthomonas oryzae pv. oryzae bakteri penyebab hawar daun pada padi: Isolasi, karakterisasi, dan telaah mutagenesis dengan transposon. Makara Sains, 15: 89-96.
Direct Link - Wang, M. and Q. Ma, 2011. Antagonistic actinomycete XN-1 from Phyllosphere microorganisms of cucumber to control Corynespora cassiicola. Cucurbit Gen. Coop. Rep., 33: 17-21.
Direct Link - Weller, D.M., 1988. Biological control of soilborne plant pathogens in the rhizosphere with bacteria. Annu. Rev. Phytopathol., 26: 379-407.
CrossRefDirect Link