
M. Mallikarjuna
Department of Microbiology, Yogi Vemana University, Kadapa, 516003, India
B. Jayapal Gowdu
Department of Microbiology, Yogi Vemana University, Kadapa, 516003, India
Asian Journal of Plant Pathology
Year: 2015 | Volume: 9 | Issue: 4 | Page No.: 175-188







ABSTRACT
The present study was aimed to identify charcoal rot pathogen of groundnut (Arachis hypogaea L.) in Anantapur district, Andhra Pradesh and assess the interactions among the fungal isolates from rhizosphere. During field survey, a potential charcoal rot pathogen Macrophomina phaseolina was isolated and identified. About 10 different rhizosphere soils of healthy groundnut plants were selected for isolation of fungi. Among 25 isolates assayed for antagonism, ten different fungi significantly inhibited colony growth of M. phaseolina in dual culture plates and morphological identification was done at (400X). Antagonistic fungi inflicted 25.6-41% mycelial growth inhibition in M. phaseolina. Colony growth and sclerotia production of M. phaseolina ceased effectively by Emericella nidulans and E. rugulosus, respectively. Mutual inhibition between Alternaria alteranata, Aspergillus flavus, A. terreus, Penicillium chrysogenum and M. phaseolina was observed. Penicillium oxalicum, Gliocladium roseum and Aspergillus versicolor significantly reduces growth with <1 mm demarcation lineat 7 and 14 Days after Inoculation (DAI) on PDA medium. These antagonistic interactions influence the growth of the pathogen.
PDF Abstract XML References Citation
Received: September 13, 2015;
Accepted: November 16, 2015;
Published: November 23, 2015
How to cite this article
M. Mallikarjuna and B. Jayapal Gowdu, 2015. Isolation, Identification and in vitro Screening of Rhizospheric Fungi for Biological Control of Macrophomina phaseolina. Asian Journal of Plant Pathology, 9: 175-188.
URL: https://scialert.net/abstract/?doi=ajppaj.2015.175.188
URL: https://scialert.net/abstract/?doi=ajppaj.2015.175.188
INTRODUCTION
Peanut or groundnut (Arachis hypogaea L.) is mainly produced in the states of Andhra Pradesh, Tamil Nadu, Gujarat, Karnataka and Maharashtra in India (Talawar, 2004). In arid regions of India, many annual crops and weeds are susceptible to the charcoal rot pathogen M. phaseolina (Tassi) goid = Rhizoctonia bataticola (Taub.) Butler (Lodha et al., 1986). In groundnut it is known to cause wilt as well as root, stem, peg and pod rots and leaf spots in seedlings and older plants (Pattee and Young, 1982). This pathogen also causes seedling blight, root rot and charcoal rot diseases on more than 500 plant species from more than 100 families (Singleton et al., 1992) distributed worldwide. The disease appears in hot and dry weather when soil temperature is 80-95°F (27-35°C) for 2-3 weeks. The disease control is inefficient or difficult by using the chemical fungicides. The non-judicious use of a wide range of fungicides has led to harmful effects on to environment and increased resistant pathogen populations (Soytong et al., 2001).
The search for effective alternative approaches to chemical control which have minimal deleterious effects, more eco-friendly and will contribute to the goal of sustainability in agriculture is needed (Jain et al., 2009). Because of the increasing awareness about problems associated with the chemical based disease control, biological control has received considerable attention during the last 40 years as a potential alternative (Cook and Baker, 1983). Biological control of plant disease by microorganisms, especially of soil-borne plant pathogens and nematodes, has been considered as a more natural and environmentally acceptable alternative to existing chemical treatment methods (Baker and Paulitz, 1996). Disease suppression by biocontrol agents is the sustained manifestation of interactions among plant, pathogen, bioagent, microbial community on and around the plant and the physical environment (Handelsman and Stabb, 1996). In their natural habitats, interactions between microbial cells can be as complex as they are dynamic. The outcome of each interaction is determined by factors such as the species, physiological condition and microbial load present; diverse physicochemical and biophysical parameters including temperature, water activity, chaotropicity and pH; motility of and/or distance between competing microbes and antimicrobials produced by the stress biology and other metabolic attributes of the microbial participants (Cray et al., 2013). Fungal colonies growing in vicinity may effect important changes in one another or they may require close hyphal connection. Variations include trophic stimulation, morphogenetic alterations and various degrees of growth inhibition. Interactions between hyphae appear to be substantial in defining the pattern of colonization of dung. In the recent years, there has been a worldwide swing towards the use of eco-friendly methods for protecting crops from pests and diseases (Rao et al., 1998). Although the number of biocontrol products is increasing, they still represent only about 1% of agricultural chemical sales (Fravel, 2005).
With this in view, the present investigation was carried out to isolate, identify and examine the efficacy of potential soil fungi from rhizosphere of peanut against Macrophomina phaseolina, a serious plant pathogen of economic importance under in vitro conditions.
MATERIALS AND METHODS
Soil sampling and isolation of fungi: Soil trials were taken from 10 different groundnut rising areas near Yogi Vemana University, Kadapa, Pendlimarri, Siddautavam (Lankamalleswara forest) and Kadiri during September 2012. Soil was taken at 10 cm depth. Samples were sieved and air dried for 3-5 days at 18°C. After drying, samples were kept at 10°C until used. Normal microflora were isolated from rhizosphere soil of healthy groundnut plants by serial dilution technique on Rose Bengal agar medium (Glucose-10 g, peptone-5 g, KH2PO4-1 g, MgSO4.7H2O-0.05 g, streptomycin-30 mg, RoseBengal-0.035 g, agar-15 g, distilled water-1000 mL ) (Martin, 1950) and Trichoderma selective medium (MgSO4.7H2O-0.2 g, K2HPO4-0.9 g, KCl-0.15 g, NH4NO3-3.0 g, glucose-3.0 g, agar-15 g, Rose Bengal-0.15 g, chloramphenicol-0.25 g, distilled water-1000 mL, pH-6.5) (Elad et al., 1981) and incubated at 27±3°C for a week (Rabeendran et al., 1998). Resulting colonies were purified on PDA plates amended with streptomycin (20 mg L–1) and identified using standard mycological keys.
Identification of soil fungi: The morphological characteristics used for identification of isolates were mycelial growth, pigmentation and presence of fungal structures such as conidia, pycnidia and cleistothecia. Mycelial disc from single spore culture was placed in the center of a 90 mm diameter PDA plate and incubated at room temperature for seven days. Isolates were grouped according to colony appearance and pigmentation. The mycelial growth rates of the fungal isolates were determined from PDA plates. Petri dishes containing PDA were each centrally inoculated with a 5 mm diameter agar plug from 7 day old cultures of each isolate, to determine their Average Linear Growth Rates (ALGR) (Table 1). Radial mycelial growth was recorded every 24 h during this period.
| Table 1: | Average linear growth rates of fungi isolates |
 | |
ALGR: Average linear growth rates, Each value is an average of 3 replicate samples, ±Standard Error | |
The colony diameter was measured as the mean of two perpendicular diameters, based on the method of Elad et al. (1981). Average Linear Growth Rates (ALGR) were calculated using following equation:
ALGR (mm per day) = (C7-C1)/C1
where, C7 is colony diameter in milliliter after seven days and C1 is colony diameter in milliliter after one day of incubation.
Three replicates were run for each treatment. Micromorphological features in fungi were studied by mounting fungal mycelium and examining under the light microscope (Olympus model BX-50F4). Images were captured using a camera (Canon model PC1737). Perennial structures that were formed in the agar medium, such as conidia were cut out from the medium and mounted in lactic acid. The sizes of the various structures such as conidia were determined by averaging the measurements of 5 samples of each structure. The morphological features of the conidia such as the shape and septation of conidia were recorded photographically; the size of conidia was measured and compared with the available literature.
Isolation of Macrophomina phaseolina: From June to January, charcoal rot-affected stems or roots of peanut (Arachis hypogaea L.) were identified, collected from different cropping regions of Anantapur and Kadapa. Two different methods were used for isolation of M. phaseolina.
Soil isolation: Isolates were obtained from soil using a soil assay technique based on the method of Alabouvette (1976). Soil samples were dried in an oven at 37°C, crushed through a 1 mm mesh sieve, mixed thoroughly and subsamples of 5 g were taken. Each subsample was submerged in 0.52% NaOCl for 10 min. The soil-NaOCl mixture was washed with distilled water through two sieves of mesh sizes, 180 and 45 μm, respectively. The residue retained on the 45 μm mesh sieve was incorporated into 100 mL of a Semi Selective medium (SS medium) for M. phaseolina. This consisted of Potato Dextrose Agar (PDA; 39 g L–1) maintained at 55°C in a water bath, with the following ingredients added: 1.5 mL 0.52% NaOCl, 1 mL 0.5% chloramphenicol dissolved in 95% alcohol and 10 mL 2.25% quintozene (PCNB). The medium was poured into 90 mm petri dishes to solidify and then incubated at 27±3°C.
Host tissue isolations: Uprooted plants were rinsed under running tap water and blotted dry with a sterile paper towel. Separate subsamples of roots and stems of each plant were treated by immersion in 0.5% NaOCl for 10 min to eliminate secondary invaders, followed by rinsing for 30 min in sterile distilled water. Tissues excised from dying rootlets, necrotic taproots and stems were cut into 2-3 mm long fragments and 18 of these were plated on three petri dishes containing the above described SS-medium and incubated at 27±3°Cfor 5-8 days. Colonies of M. phaseolina each appeared as a ring of fluffy white mycelium surrounding central area with black microsclerotia. Each colony was observed under the microscope for the presence of microsclerotia. Typical colonies from soil and host tissue isolations were propagated further onto PDA and stored at 5°C.
Dual culture technique: To study the antagonistic effects of the soil fungi isolates on M. phaseolina, dual culture plate technique (Dennis and Webster, 1971) was used. Different antagonists and pathogen combinations were examined on 20 mL of PDA in 90 mm petriplates, with three replicate plates per treatment. For dual culture technique, a mycelia plug (5 mm in diameter) taken from actively growing 7 day old culture of M. phaseolina and soil fungi isolates placed 80 mm apart from each other on the PDA. For control treatments, a plug of M. phaseolina was placed on the PDA medium against PDA plug. The plates were incubated at 27±3°C. Observations on the antagonistic activities of isolates on M. phaseolina were recorded after every 24 h for 7 days and inhibition percentage was calculated using the following formula (Edgington et al., 1971):
Inhibition percentage (%) = A1-A2/A1×100
where, A1 is the colony area of uninhibited M. phaseolina in the control plate and A2 is the colony area of M. phaseolina in dual culture plate.
RESULTS
Isolation of soil fungi: About 25 different fungi were isolated from peanut fields. Among them, Emericella nidulans, Emericella rugulosus, Alternaria alteranata, Aspergillus flavus, Aspergillus terreus, Aspergillus versicolor, Penicillium oxalicum, Penicillium chrysogenum, Rhizoctonia solani and Gliocladium roseum were selected for further characterization based on inhibition of Macrophomina phaseolina.
Identification of soil fungi: Each isolate was cultured on PDA for identification work. The various filamentous fungi isolated were identified, based on the following characteristics in accordance with the literature available.
Emericella nidulans: It is observed that an encountered isolate appear imperfect or anamorphic stage namely Aspergillus nidulans. The ALGR of culture observed as 13.5 mm per day at 27°C, conidia green from becoming brownish when mature to produce abundant. Cleistothecia, globose or oval shape, 130×350 μm in diameter and surrounded by hülle cells (Fig. 1). Hülle cells ellipsoidal to globose, 25 μm in diameter. Asci are globose to subglobose, 8-spored. Ascospores are red, smooth and 2×3.6 μm, with two narrow longitudinal furrows as similar described by (Domsch et al., 1993).
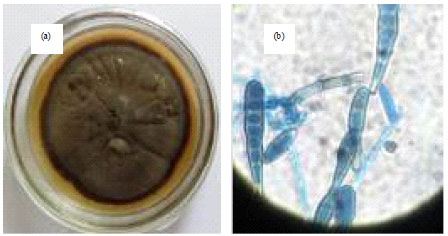
Emericella rugulosus: Colonies on PDA attains 80 mm after seven days at 27°C and its ALGR is 12.5 mm per day. Colony color dark green and on reverse brown orange to dark purple brown (Fig. 2).
 | |
| Fig. 1(a-e): | (a) Emericella nidulans, (b) Cleistothecia, (c) Hülle cells, (d) Ascospores and (e) Asci (400X) |
 | |
| Fig. 2(a-c): | (a) Emericella rugulosus, (b) Ascospores and (c) Hülle cells (400X) |
Conidial heads are short columnar, 60 μ long by 60 μ wide. Conidiophore smooth, brown in color, 60 by 4 μ in diameter. Vesicle flask shaped 10 μ in diameter. Sterigmata biseriate, matulae 5 μ long by 6 μ wide. Conidia globose, small 4 μ in diameter. Cleistothecia 325 μ in diameter.
 | |
| Fig. 3(a-b): | (a) Aspergillus flavus and (b) Conidia |
 | |
| Fig. 4(a-b): | (a) Alternaria alternata and (b) Muriformconidia |
Ascospores purple brown in color, having two 0.8 μ thick equatorial crests, rugulose surface, spores body having double crests 6 μ long, excluding crest 4.4 μ long by 3.8 μ wide, Hülle cells globose, 20 μ in diameter (Afzal et al., 2013).
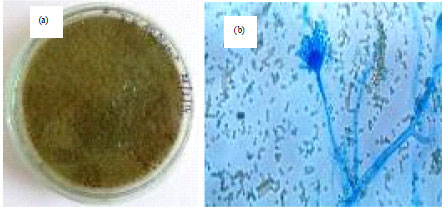
Aspergillus flavus: Colonies on potato dextrose agar widely spreading, growing rapidly, with ALGR 13.5 mm per day: Conidial heads pale yellow when young, becoming dark green in age, in older cultures deep grey-green, reverse colourless to pale yellow brown (Fig. 3). Conidiophores across separately from the substratum, 800 of 1200 μm long, 18 μm in diameter, broadening upward. Walls were pitted as to appear rough of spiny with low magnification, gradually enlarging upward to form a vesicle 30 and 60 μm in diameter (Xalxo et al., 2013).
Alternaria alternata:Colonies grew on PDA reach 70 to 80 mm in 7 days and ALGR is 10 mm per day. Olive black colony without aerial mycelium (Fig. 4). Conidiophores short, simple, straight, branched or unbranched, up to 40 μm long conidia often in long branched chains of 10 or, uniform with 6 transverse septa, longitudinal or obligue septa present. The conidia were muriform shape and light brown colour. The length of their conidia was varied from 42 μm (Watanabe, 2010).
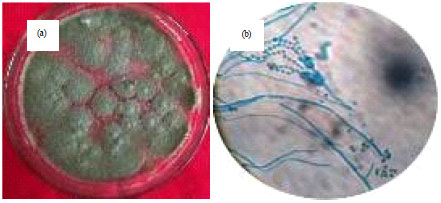
Aspergillus terreus: Colonies on PDA media appeared as rapidly growing powdery colonies with a characteristic cinnamon-brown colour on the surface and a yellow to beige-brown color on the reverse.
 | |
| Fig. 5(a-b): | (a) Aspergillus terreus and (b) Conidia |
 | |
| Fig. 6(a-b): | (a) Penicillium oxalicum and (b) Phialides |
The ALGR of colony calculated as 8.3 mm per day, exudates amber colored (Fig. 5). Conidial heads long columnar, compact with uniform diameter, cinnamon brown, 550×30 μm, conidiophores smooth, colorless, 250×6 μm, vesicles hemispherical, dome-like ,16 μm in diameter, phialides biseriate; conidia globose to subglobose and 2.4 μm in diameter (Xalxo et al., 2013).
Penicillium oxalicum: Colony attains 54 mm diameter in 7 days and ALGR is 8 mm per day. Colonies are radially sulcate, sometimes convolute, centrally depressed, moderately deep to deep, surface texture velutinous to floccose, mycelium white, conidiogenesis light to moderate, greyish olive, clear brown exudate sometimes present and reverse beige to reddish brown (Fig. 6). Conidiophores arising from surface mycelium; stipes 290 μm in length with thin, smooth walls, characteristically terminating in verticils of 3 metulae; metulae 20 μm long; phialides were ampulliform. Conidia measured 3.02 μm ellipsoidal, smooth and in columns (Molar et al., 2013).
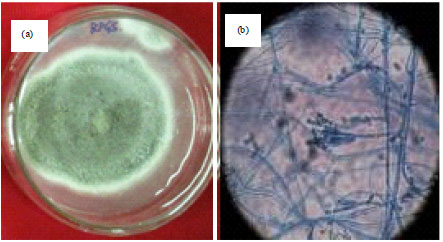
Gliocladium roseum: Colony appears as green in color with white edged margins reaches 50 mm in 7 days on PDA and ALGR is 8.8 mm per day, at maturity color of the colony turns into powdery green (Fig. 7). Conidiophores hyaline, erect, branched with penicillate phialides bearing single spore masses. Conidia hyaline, long-ellipsoidal, subglobose. Perithecia 250×300 μm. Asci 40×7.5 μm, ascospores 11.3×3.8 μm (Watanabe, 2010).
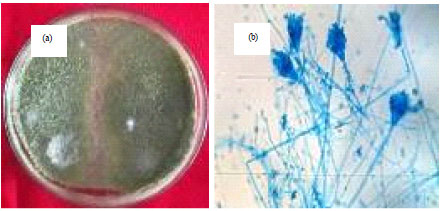
Penicillium chrysogenum: Colonies on PDA appear to grow 8.4 mm per day. Green-blue velvety with a heap of white mycelium at the centre and reverse is yellow-white (Fig. 8).
 | |
| Fig. 7(a-b): | (a) Gliocladium roseum and (b) Conidia |
 | |
| Fig. 8(a-b): | (a) Penicillium chrysogenum and (b) Phialides |
The penicillia are asymmetrical and often complex with diverging branching patterns. The conidiophores are smooth-walled with size 1000 μm. There are 3 to 5 metulae of 10×2 μm. The phialides are aggregated in clusters of 4-6; they are ampulliform and are 9×2 μm. The conidia are subglobose to elliptical, smooth-walled arranged in long irregular chains. They are 4×3.5 μm.
Aspergillus versicolor: Colonies on PDA at 27°C grew slowly, reaching 65 mm in diameter after 7 days its ALGR is 10.5 mm per day. Colonies centrally rising, velvety to rather floccose, colour blue-green, reverse pale yellowish and exudates absent (Fig. 9). It was interesting to find that a large number of heads were fragmentary and like penicillium. Conidiophores monoverticillate, hyaline, smooth-walled, 8×3 μm. Metulae 9×4 μm long. Phialides, 8×3 μm long (Xalxo et al., 2013).
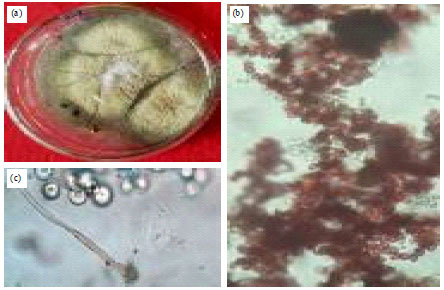
Rhizoctonia solani: Colonies on Potato Dextrose Agar (PDA) grow at a rate of 20 mm per day at 27°C. The colony range in color from buff to black (Fig. 10). Sclerotia are produced on the surface of cultures after 4 to 6 weeks and are irregularly shaped, light tan to black and usually >1 mm (0.04 in.) in width (Watanabe, 2010).
Identification of Macrophomina phaseolina: Macrophomina phaseolina was isolated by both soil and host tissue methods were identified following (Watanabe, 2010).
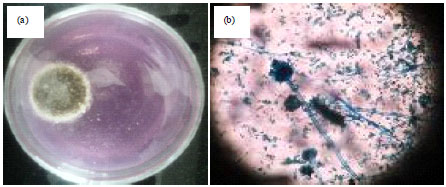 | |
| Fig. 9(a-b): | (a) Aspergillus versicolor and (b) Conidia |
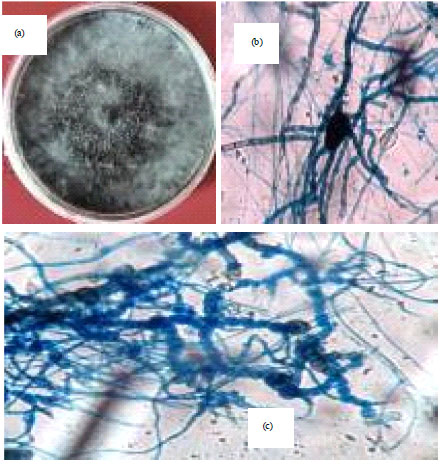 | |
| Fig. 10(a-c): | (a) Rhizoctonia solani, (b) Microsclerotia and (c) Mycelia (400x) |
Observations were made with a light microscope (Olympus model BX-50F4) and using a camera (Canon model PC1737). Colony characters and pigment production were noted after 3 day of growth on PDA at 27±3°C (Fig. 11). Pycnidia black, globose, ostiolate apically. Conidiophores hyaline, simple, cylindrical, narrowing apically. Conidia hyaline, cylindrical, 1-celled. Microsclerotia black, homogenous in size. Pycnidia 200 μm in diameter; ostioles 30 μm in diameter. Conidiophores 20×6 μm. Conidia 30×6 μm. Sclerotia 120 μm in diameter. Hyphae 7.5 μm wide (Watanabe, 2010).
Effect of antagonistic fungi on mycelial growth of Macrophomina phaseolina in vitro: All the ten soil isolates exhibited antagonistic potential against M. phaseolina by inhibiting its mycelial growth.
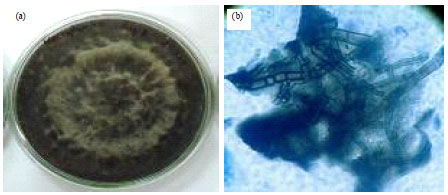 | |
| Fig. 11(a-b): | (a) Macrophomina phaseolina and (b) Microsclerotia (400x) |
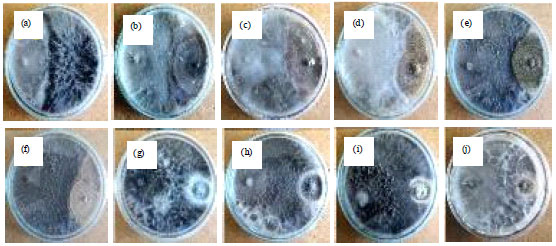 | |
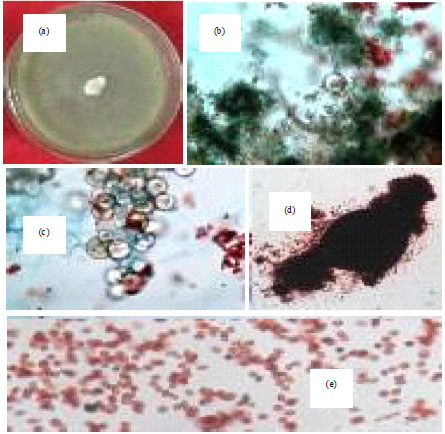
| Fig. 12(a-j): | Dual culture and colony interactions of rhizosphere fungi and M. phaseolina, (a) M. phaseolina+R. solani, (b) M. phaseolina+E. nidulans, (c) M. phaseolina +E. rugulosus, (d) M. phaseolina+A. flavus, (e) M. phaseolina+A. alternata, (f) M. phaseolina+A. terreus, (g) M. phaseolina+P. oxalicum, (h) M. phaseolina +G. roseum, (i) M. phaseolina+P. chrysogenum and (j) M. phaseolina+A. versicolor |
The mycelial inhibition on seventh day ranged from 26.9 to 51.9% for M. phaseolina. After inoculation, growth of M. phaseolina was found to be inhibited by isolates and attained a growth of 47.3, 41.2, 36.8, 35.1, 35, 33.4, 32.4, 30.6, 27.4, 26.4 and 25.6 mm (Table 2). This indicates that isolate E. nidulans (40.9%) proving more effective against the pathogen, followed by E. rugulosus (39.0%) had maximum antifungal activity against M. phaseolina compared to the other isolated fungi (Fig. 12). Rhizoctonia solani inhibits the growth of M. phaseolina by 51%. E. nidulans was more antagonistic at the day of 14 than at 7dai and significantly suppresses the growth of pathogen and decreases sclerotia production. Mutually inhibitory interactions of M. phaseolina with E. rugulosus, A. flavus and P. oxalicum were observed and rated 4 according to colony interactions chart proposed by Skidmore and Dickinson (1976). Intermingling growth where the M. phaseolina has ceased growth and is being overgrown by R. solani and rated as 3 as given in Table 3. Mutually slight inhibition of hyphal growth was observed between A. alternata and M. phaseolina rated as 3. Initially, A. terreus shows inhibition >2 mm over M. phaseolina and later it comes to 1 mm of inhibition. Slight inhibition of M. phaseolina with P. oxalicum, G. roseum, A. versicolor and P. chrysogenum where <1 mm demarcation line was clearly visible as shown in (Fig. 12).
| Table 2: | Dual culture technique |
 | |
| Each value is an average of 3 replicate samples, ± Standard Error | |
| Table 3: | Five separate modes of interacting colony growth |
 | |
DAI: Days after inoculation, (1) Mutually intermingling growth where both fungi grew into one another without any macroscopic signs of interaction, (2) Intermingling growth where the fungus being observed is growing into the opposed fungus either above or below or above and below its colony and its corollary, (3) Intermingling growth where the fungus under observation has ceased growth and is being overgrown by another colony, (4) Slight inhibition where the fungi approached each other until almost in contact and a narrow demarcation line, c. 1-1. mm, between the two colonies was clearly visible and (5) Mutual inhibition at a distance of > 2 mm | |
DISCUSSION
Macrophomina phaseolina was isolated from infected groundnut fields in Anantapur region and proved to be pathogenic isolate causing charcoal rot and seedling diseases of groundnut plants. Among 10 different isolated fungal species, A. flavus, A. terreus are dominant, E. nidulans is subdominant and E. rugulosus, A. alternata, P. oxalicum, R. solani are extant in rhizosphere as reported by Subrahmanyam and Rao (1977). Though R. solani shows antagonistic activity, it intermingles and overgrown on pathogen. The tested E. nidulans and E. rugulosus showed antibiotic mechanism to inhibit growth of M. phaseolina which was supported by Sibounnavong (2012) that antibiotic substance from E. nidulans and E. rugulosus could inhibit the growth and break the cells of F. oxysporum f. sp. lycopersici. Alternaria alternata checks the growth of M. phaseolina was held by Babu et al. (2003), who identified A. alternata produces a red metabolite in potato dextrose broth and found to be toxic to waterhyacinth. Inhibition of M. phaseolina by A. flavus was supported by Cotty and Bayman (1993) indicated that atoxigenic strains of A. flavus will competitively exclude the aflatoxin producing strains of same fungal species. A. terreus as a biocontrol agent against M. phaseolina was similarly reported by Melo et al. (2006) specified sclerotial viability of the pathogen was found to be affected by treatment of sclerotia, with conidia of the hyperparasite A. terreus. Penicillium oxalicum slightly inhibits M. phaseolina supported by De Cal et al. (1997) identified disease reduction was observed when P. oxalicum was applied before or after inoculation with the pathogen. Penicillium chrysogenum constrains the growth of M. phaseolina (Dong et al., 2006) reported that dry mycelium of P. chrysogenum contributed to induced resistance against Verticillium dahliae in cotton. Gliocladium roseum slightly obliges the growth of M. phaseolina supported by Rodriguez-Kabana and Morgan-Jones (1988) indicated that G. roseum produce a diffusible yellow pigment in PDA cultures and its correlation to biocontrol efficacy. Aspergillus versicolor slightly hinders M. phaseolina strengthen by (Bhattacharyya et al., 1985) identified A. versicolor as a Biological Control Agent (BCA) against M. phaseolina.
CONCLUSION
It is concluded that this research finding E. nidulans, E. rugulosus, A. alternata, A. flavus, A. terreus, P. oxalicum, P. chrysogenum, G. roseum and A. versicolor developed as bioagents to control M. phaseolina under in vitro conditions. Further research has to be done to characterize the volatile and non-volatile metabolites from potential antagonistic fungi to assess their effect on M. phaseolina and in vivo control of charcoal rot of groundnut caused by M. phaseolina.
REFERENCES
- Afzal, H., S. Shazad and S.Q.U. Nisa, 2013. Morphological identification of Aspergillus species from the soil of larkana district (Sindh, Pakistan). Asian J. Agri. Biol., 1: 105-117.
Direct Link - Alabouvette, C., 1976. [The ecology of parasitic fungi in soil. VIII. An ecological study, by means of a selective method of investigation, of Macrophomina phaseolina in soil]. Annales Phytopathologie, 8: 147-157, (In Spanish).
Direct Link - Molar, R.A., P.A.P. Munoz, I.M. Zequera, J.A.C. Fasio, R.S.G. Estrada and J.L. Felix, 2013. [Morphological and molecular identification of Penicillium oxalicum causing stem and fruit rot in tomato]. Revista Mexicana Fitopatologia, 31: 13-19, (In Spanish).
Direct Link - Babu, R.M., A. Sajeena and K. Seetharaman, 2003. Bioassay of the potentiality of Alternaria alternata (Fr.) keissler as a bioherbicide to control waterhyacinth and other aquatic weeds. Crop Prot., 22: 1005-1013.
CrossRefDirect Link - Bhattacharyya, D., S. Basu, J.P. Chattapadhyay and S.K. Bose, 1985. Biocontrol of macrophomina root-rot disease of jute by an antagonistic organism, Aspergillus versicolor. Plant Soil, 87: 435-438.
CrossRefDirect Link - Cotty, P.J. and P. Bayman, 1993. Competitive exclusion of a toxigenic strain of Aspergillus flavus by an atoxigenic strain. Am. Phytopathol. Soc., 83: 1283-1287.
Direct Link - Cray, J.A., A.N.W. Bell, P. Bhaganna, A.Y. Mswaka, D.J. Timson and J.E. Hallsworth, 2013. The biology of habitat dominance; can microbes behave as weeds? Microb. Biotechnol., 6: 453-492.
Direct Link - De Cal, A., S. Pascual and P. Melgarejo, 1997. Involvement of resistance induction by Penicillum oxalicum in the biocontrol of tomato wilt. Plant Pathol., 46: 72-79.
CrossRefDirect Link - Dong, H., X. Zhang, Y. Choen, Y. Zhou, W. Li and Z. Li, 2006. Dry mycelium of Penicillium chrysogenum protects cotton plants against wilt diseases and increases yield under field conditions. Crop Prot., 25: 324-330.
CrossRefDirect Link - Edgington, L.V., K.L. Khew and G.L. Barron, 1971. Fungitoxic spectrum of benzimidazole compounds. Phytopathology, 61: 42-44.
CrossRefDirect Link - Elad, Y., I. Chet and Y. Henis, 1981. A selective medium for improving quantitative isolation of Trichoderma spp. from soil. Phytoparasitica, 9: 59-67.
CrossRefDirect Link - Fravel, D., 2005. Commercialization and implementation of biocontrol. Annu. Rev. Phytopathol., 43: 337-359.
CrossRefPubMedDirect Link - Handelsman, J. and E.V. Stabb, 1996. Biocontrol of soilborne plant pathogens. Plant Cell, 8: 1855-1869.
Direct Link - Martin, J.P., 1950. Use of acid, rose bengal and streptomycin in the plate method for estimating soil fungi. Soil Sci., 69: 215-232.
Direct Link - Melo, I.S., J.L. Faull and R.S. Nascimento, 2006. Antagonism of Aspergillus terreus to Sclerotinia sclerotiorum. Braz. J. Microbiol., 37: 417-419.
CrossRefDirect Link - Rao, M.S., P.P. Reddy and M. Nagesh, 1998. Evaluation of plant based formulations of Trichoderma harzianum for the management of Meloidogyne incognita on egg plant. Nematol. Mediterr., 26: 59-62.
Direct Link - Rodriguez-Kabana, R. and G. Morgan-Jones, 1988. Potential for nematode control by mycofloras endemic in the tropics. J. Nematol., 20: 191-203.
Direct Link - Skidmore, A.M. and C.H. Dickinson, 1976. Colony interactions and hyphal interference between Septoria nodorum and phylloplane fungi. Trans. Br. Mycol. Soc., 66: 57-64.
CrossRefDirect Link - Sibounnavong, P., 2012. Screening of Emericella nidulans for biological control of tomato Fusarium wilt in Lao PDR. J. Agric. Technol., 8: 241-260.
Direct Link - Soytong, K., S. Kanokmedhakul, V. Kukongviriyapa and M. Isobe, 2001. Application of Chaetomium species (Ketomium®) as a new broad spectrum biological fungicide for plant disease control: A review article. Fungal Diversity, 7: 1-15.
Direct Link - Subrahmanyam, P. and A.S. Rao, 1977. Rhizosphere and geocarposphere mycoflora of groundnut (Arachis hypogaea Linn). Proc. Indian Acad. Sci. Sect. B, 85: 420-431.
Direct Link - Xalxo, P.C., D. Karkun and A.N. Poddar, 2013. Rhizospheric fungal associations of root knot nematode infested Cucurbits: In vitro assessment of their nematicidal potential. Res. J. Microbiol., 8: 81-91.
CrossRef - Lodha, S., G.K. Gupta and S. Singh, 1986. Crop disease situation and some new records in Indian arid zone. Ann. Arid Zone, 25: 311-320.
Direct Link - Dennis, C. and J. Webster, 1971. Antagonistic properties of species-groups of Trichoderma: I. Production of non-volatile antibiotics. Trans. Br. Mycol. Soc., 57: 25-39.
CrossRefDirect Link