
Jamsari
Plant Breeding Section, Faculty of Agricultural, Andalas University, Padang, 25163, West Sumatera, Indonesia
Istino Ferita
Plant Breeding Section, Faculty of Agricultural, Andalas University, Padang, 25163, West Sumatera, Indonesia
Lily Syukriani
Plant Breeding Section, Faculty of Agricultural, Andalas University, Padang, 25163, West Sumatera, Indonesia
Helni Lalan
Plant Breeding Section, Faculty of Agricultural, Andalas University, Padang, 25163, West Sumatera, Indonesia
Friedrich Herberg
Department of Biochemistry, Kassel University, Heinrich-Plett Str. 40, 34132, Building AVZ I, 2, Stock, Kassel, Germany
Wolfgang Nellen
Department of Genetics, Kassel University, Heinrich-Plett Str. 40, 34132, Kassel, Germany
Asian Journal of Plant Pathology
Year: 2014 | Volume: 8 | Issue: 2 | Page No.: 34-44







ABSTRACT
Whiteflies have attracted intensive attention from many agriculturists due to their direct feeding and plant virus transmission. The economic losses depend on certain species population existing on the field during chilli-pepper cultivation. More over, biotype characteristic will determine the effectivity of applied pest management. Based on this, detailed information on whiteflies in chilli-pepper cultivation is very important. We applied mitochondria cytochrome oxidase unit I gene sequence for detecting species and biotype of whiteflies existing in West Sumatera. Two species were successfully identified i.e., Bemisia tabaci and Trialeourides vaporariorum that are dominant in this region. Distribution of both species are separated by altitude, where B. tabaci exist predominantly from low altitude to medium altitude, whereas in high altitude it is not common. On the other hand, T. vaporariorum distribute mainly in high altitude and not in medium altitude. Furthermore biotype determination on B. tabaci identified the existence of B biotype in all areas studied.
PDF Abstract XML References Citation
Received: September 17, 2014;
Accepted: October 31, 2014;
Published: December 08, 2014
How to cite this article
Jamsari, Istino Ferita, Lily Syukriani, Helni Lalan, Friedrich Herberg and Wolfgang Nellen, 2014. Existence of Two Distinct Whiteflies in Chilli-Pepper Cultivation in West
Sumatra-Indonesia Based on Mitochondria Cytochrome Oxidase I Gene Sequences. Asian Journal of Plant Pathology, 8: 34-44.
URL: https://scialert.net/abstract/?doi=ajppaj.2014.34.44
URL: https://scialert.net/abstract/?doi=ajppaj.2014.34.44
INTRODUCTION
Whiteflies cover aproximately 1,500 species distributed almost all over the world (Martin, 2004). Their effect on agricultural production was reported by many authors from tropical to sub-tropical regions and amounts to billions of dollars. Economic loses caused by whitefly are not only due to crop losses by direct feeding, sucking the sap from the plant and finally disturbing plant growth. The pest also produce honeymildew, encouraging sooty mould growth and finally decreasing plant vigour (Jelinek, 2010). The most dangerous, whiteflies are responsible for transmission of about 110 plant virus species (Jones, 2003; Martin et al., 2000; Mugiira et al., 2008; Qiu et al., 2009).
The family of whitefly is groupped in the order of Hemiptera (Von Dohlen and Moran, 1995) and contains two subfamilies, Aleurodicinae and Aleyrodinae (Mound and Halsey, 1978). Two species from subfamily Aleyrodinae; Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) and Trialeurodes vaporariorum (West.) (Hemiptera: Aleyrodidae) attract great attention from many agricultural practicists as well as scientists from many tropical and sub tropical continents. This is due to their role in virus transmission and spreading in a wide variety of vegetable, ornamental plants and other crops worldwide (Jones, 2003; Ma et al., 2007).
West Sumatera is one of the chilli pepper production centres in Indonesia. In this region chilli fruit is one of the food components playing an important role in people’s diet. Based on this reason, chilli production has a significant economic role not only in the province but also on the national level. However, pepper yellow leafcurl disease (PepYLCD) which is caused by a group of begomovirus attributed as peppper yellow leaf curl virus (PepYLCV) which is transmitted by whitefly has reduced chilli production during in the last two decades (Meliansyah et al., 2011).
Effectivity of virus transmission is determined by its vector. Therefore, knowledge of the species is indispensable for providing basic knowledge of pest and disease interaction in terms of epidemiology studies in integrated pest management. It is also well known that compatibility plays an important role in the success of virus transmisssion via whiteflies. For this reason understanding of genetic variation between pathogens and their vectors could disclose compatibility mechanisms that could be useful for development of disease management.
So far, in Indonesia, Bemisis tabacci is the only species of whitefly reported as vector for PepYLCV transmision, hence existence of another whitefly species has been reported by some authors in Java Island (Fitriasari, 2010; Hartono and Wijonarko, 2007). Currently, based on morphological and bioassay analysis only two biotypes namely B-biotype and non-B biotype are currently reported (Hidayat et al., 2004). The difference in biotype was believed also to be associated with fast growing and spreading of viruses and also their resistence against some insecticides (Horowitz et al., 2011; Shadmany et al., 2013). Certain biotype were reported to be more resistent against organophosphates, carbamates and pyrethroids (Scott et al., 2007). Q-Biotype for instnace is known to be more resistant, or at least has reduced susceptibility, to a number of neonicotinoids (Pascual and Callejas, 2004; Horowitz et al., 2005) and some insect growth regulators. This in turn forces to adapt chemical treatment. Therefore, comprehensive information on genetic variability of whitefly including their biotype could enhance the effectivitiy of disease management in chilli pepper cultivation.
Many studies dealing with assessing genetic variation of whitefly have been carried out by using random DNA sequence for instance RAPD-PCR (Gawel and Bartlett, 1993; Perring et al., 1993; De Barro and Driver, 1997; Khasdan et al., 2005) and AFLP (Amplified Fragment Length Polymorphism) (Cervera et al., 2000). Currently, analysis based on specific conserved sequence region for instance ribosomal and mitochondria cytochrome oxidase unit I (mtCOI) gene sequence (De Barro et al., 2005; Dinsdale et al., 2010; Rocha et al., 2011) become to be more common in this field. Biotype determination is also suggested to be possible by this method.
The present study was reported on genetic variation of whitefly population in chilli cultivation in West Sumatera. Genetic variation were assessed using mtCOI gene sequence and used the data also for biotype determination among species.
MATERIALS AND METHODS
Sample collection: Fourtytwo whiteflies were collected randomly from three different altitudes representing low, medium and highland. Lowland region (0-300 m above sea level (asl)) was represented by three regencies: Padang, Pesisir Selatan and Pasaman Barat. Mediumland region (301-700 m asl) was represented by Solok and 50 Kota and highland region (700 m asl) was represented by Agam and Tanah Datar. The number of samples collected from each region is listed in Table 1.
| Table 1: | Number of whitefly samples collected from chilli pepper plant from different locations in the study* |
 | |
| *One sample was provided from Bogor, designated as: BGR-R and used as reference sample | |
Whitefly from Bogor was used as a reference sample, so that total number of sample is 43. Sample collection was done by capturing nymph and adult whitefly using a zipper bag with caps on the bottom side so that the imago entered into zipper bag. Each sample was labelled and in each zipper bag contained only 1 single whitefly. Samples were stored at -20°C before use.
DNA isolation: The DNA of whitefly was isolated using a protocol by Goodwin et al. (1994) with minor modification. Briefly, DNA isolation was done as followed: The 1 single whitefly was placed into 1.5 mL centrifuge tube and crushed into powder with a micro plastic pestel with the presence of liquid nitrogen. One hundred twenty five microliter of CTAB extraction buffer containing 2% CTAB (w/v), 1.4 M NaCl, 100 mM Tris-HCl, 20 mM EDTA and 1% polyvinylpyrrolidone (PVP-40) was added to the powder, vortexed and subsequently incubated at 65°C for 5 min. One hundred twenty five microliter of chloroform:isoamyl alcohol (CI) (24:1) mixture was added and further incubated at room temperature for 20 min before centrifugation at 8,000 rpm for 5 min. The supernatant was transferred into 1.5 μL sterile centrifuge tubes and after that 10 μL of 3M sodium acetate (pH 5.2) was added. Two hundred fifty microliter of cold absolute ethanol was added and incubated for 30 min at -20°C. Supernatant was discarded after centrifugation at 11,500 rpm for 15 min. The Pellet was washed with 200 μL of 70% cold ethanol and recentrifuged again at 11,500 rpm for 2 min. Ethanol was then discarded and white pellets were dried for 10 min at room temperature. The pellets were resuspended in 10 μL of sterile water and stored at -20°C before use.
DNA amplification and partial sequencing of COI-gene: The extracted DNA was amplified using a primer pair designed from the Cytochrome Oxidase-subunit I (COI) gene sequence generated from mitochondria genome (Frohlich et al., 1999). The sequence of primer was as follow: C1-J-2195-FW (5'-TTGATTTTTTGGTCATCCAGAAGT-3') and C1-J-2195-RV (5-'TCCAATGCAATAA-TCTGCCATATTA-3') (Simon et al., 1994; Frohlich et al., 1999). Amplification using this primer pair should produce a fragment about 880 bp in size (Rocha et al., 2011). The PCR reactions were performed in a total volume of 25 μL consisting of 2 μL of DNA template, 4 μL of each forward and reverse primer mtCOI (C1-J-2195-FW/RV) (5 pmol μL-1), RTG-PCR bead (GE-Healthcare, UK) and 19 μL ddH2O. Main amplification was performed in 30 cycles using a PCR machine (Biometra-Germany), started with initial denaturation of 95°C for 3 min. Main PCR condition was set as follow: Denaturation, 95°C for 5 min, annealing temperature at 55°C for 1 min, extension for 1 min at a temperature of 72oC. One additional extensions was done at 72°C for 5 min. The PCR product was stored at 4°C before use. Controlling the success of the amplification reaction, electrophoresis was performed on 1% agarose in 0.5x TBE buffer at 100 VOLTS for 60 min (Sambrook et al., 1989). Visualization of PCR product was done using UV-transiluminator (Biometra-Germany) after gel staining with Ethidium bromide. Data was documented and stored in jpeg format. Sequencing of PCR products was performed in PT CPI (Charoen Phokphand Indonesia-Indonesia). After purification sequencing was done directly in one direction using primer C1-J-2195-FW with 20 μL of PCR products.
Sequence and bioinformatic analysis: Sequence data was edited and analyzed using BioEdit (http://www.mbio.ncsu. edu/BioEdit/bioedit.html). Homology search of the sequence product was done using BLAST programme at http://blast.ncbi.nlm.nih.gov/Blast.cgi. (Altschul et al., 1990) Alignment was done using ClustalW2 provided at: http://www.ebi.ac.uk/Tools/msa/clustalw2/ (Thompson et al., 1994) and determination of biotype was simulated in-silico by means clustering analysis. In this respect two mtCOI gene sequence from each B-, A-, Q and M-biotype which is available in NCBI nucleotide database were used as comparison during clustering.
RESULTS AND DISCUSSION
Agricultural practices in sampling location: Fourtytwo whiteflies in total were collected from three different geographical conditions: Low, medium and high altitude. High altitute (Agam and Tanah Datar) is a region where agricultural cultivation is dominated by vegetable (chilli pepper, bean and tomato). Those three vegetables are intensively cultivated in this region (Badan Pusat Statistik Kabupaten Tanah Datar, 2013). Furthermore, in some regions intensive polyculture system or rotating cropping systems composed of those vegetables is commonly practiced in the regions.
The whitefly problem is supported by favorable wet tropical agroclimatic condition, where temperature range between 15-25°C (Badan Pusat Statistik Kabupaten Tanah Datar, 2013). Due to the intensive vegetable cultivation, the amount of pesticide application particularly insecticides in this region is very high. Sometimes, farmers use pesticides concentration even more than recommended, thus provoking increase in insect resistencies. Medium altitude region (Solok and 50 Kota) generally is dominated by rainforest. Agricultural practices are dominated with cereal cultivation, whereas only some vegetables ie., chilli pepper, bean and tomato cultivation are sporadically practiced. Similarly in lowland region (Padang, Pesisir Selatan and Pasaman Barat), vegetable is only grown sporadically. The agricultural system practiced in the regions seemed to contributed to the different dominance of the whitefly species (Fig. 1b).
Species identification and distribution: All the 43 whitefly DNA samples were successfully amplified and produced an about 880 bp fragment (Fig. 1a). Prior to sequencing, PCR products were purified with Wizard SV-Gel and PCR Clean-Up System (Promega-USA). Since sequencing was done only from forward direction, only part of the total fragment could be identified. The sequence data was edited and trimmed for ambiguous nucleotides and the final fasta sequence was prepared for further analysis. However since not all PCR fragment could be sequenced completely, only 541 bases in average of each samples could be successfully used in BLAST search homology analysis for species determination.
BLAST homology search showed that 36 samples had significant (99% identity) with Cytochrome oxydase I gene of Bemisia tabacci sequence, whereas seven samples (TD-01 to TD-05, 50K-04 and AG-04) showed significant homology (99% identity) with Cytochrome oxydase I mitochondrial gene of Trialeurodes vaporariorum sequence. Based on this data distribution of both species is shown in Fig. 1b.
Figure 1b showed that all B. tabaci, could be found in both lowland (0-300 asl) (Padang, Pesisir Selatan, Pasaman Barat) and mediumland (301-700 asl) (Solok, 50 Kota) but on the other hand T. vaporariorum mainly (all five samples) could be found only at highland (>700 asl) (Tanah Datar) and are comparatively rare (1 from 5 in mediuml and 50 Kota and 1 from 6 at highland (Agam).
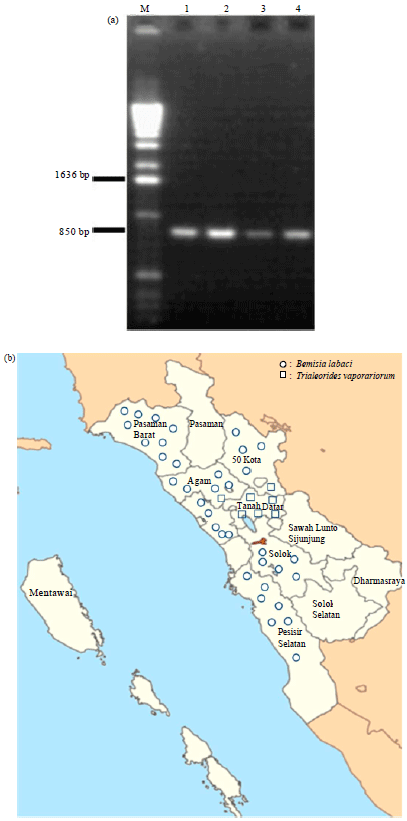 | |
| Fig. 1(a-b): | Representative of PCR products from collected whitefly samples with (a) Primer combination C1-J-2195-FW/RV and (b) Whitefly species distribution in West Sumatera. Panel A, a single 880 bp fragment is expected from the amplification of mtCOI gene. Panel B, distribution of sampling location and species of whitefly based on mtCOI sequence DNA in three different altitudes |
Locations where T. vaporariorum could be found mainly in highland region (T. Datar and Agam), eventhough in mediuml and (50 Kota) T. vaporariorum rarely could also be found. This phenomenone is in concordance with data published by Caballero (1994) where B. tabaci predominantly populated in lowland region (lower than 1000 asl) whereas T. vaporariorum predominantly populated in highland region (above 1000 m asl). This, however, was valid for equatorial region. In other lower altitude tropical region (below 700 m asl), where temperature range from 19.9-20.3°C. Trialeourides vaporariorum could still also be predominant (Lourencao et al., 2008). In Indonesia, distribution of T. vaporariorum was reported predominantly in highland region (Fitriasari, 2010). This however was observed in tomatoe production centre in West Java (Hartono and Wijonarko, 2007) and no data so far reported from chilli pepper cultivation area.
Xie et al. (2007) reported that the highland preference of T. vaporariorum is supported by a broad adaptation of all growth phases of T. vaporariorum to low temperature compared to B. tabaci. Therefore it is reasonable why T. vaporariorum is more dominant in highland regions. Another reason why T. vaporariorum is more dominant in highland region compared to B. tabaci is probably due to its broad resistancy to diverse pesticides especially insecticides. Erdogan et al. (2012) reported resistance potential to chlorpyrifos ethyl-oxon and acetylcholinesterase in Turkey, whereas Gorman et al. (2007) reported resistance to neonicotinoid and imidacloprid.
The data indicated that favorable conditions for B. tabaci and T. vaporarium are dominantly determined by agroclimatic and agricultural system.
In order to study whether T. vaporariorum plays also a role in PepYLCV distribution, we amplified all four DNA from 50K-04, AG-04, TD-05 and TD-03 using CPc forward (5’-GTACAWGCCATATACAATAACAAGGC-3) and Cpv reverse primer combination (5’-ACGCCCGYCTCGAAGGTTCG-3) (Gorsane et al., 2004). Say that these are PepYLCV primers and possibly also where they match the genome of the virus all four samples could in fact produced an about 1,600 bp fragment as expected. Do you have a negative control for this? This indicated, that T. vaporariorum in fact containing PepYLCV DNA particle. However there is not yet any proof that T. vaporariorum is involved in geminivirus transmission, although T. vaporariorum can, like B. tabaci, ingest geminivirus particles from virus infected plants (Polston et al., 1990; Rosell et al., 1999). However, almost all publications dealing with relationship between T. vaporariorum and geminivirus come to the conclusion that it has no vector function (Brown and Bird, 1992; Duffus, 1987; Polston et al., 1990). However, the role of T. vaporariorum in virus transmission must not be underestimated. Recent publications showed that this whitefly plays an important role in transmission of Tomato Infectious Chlorosis Virus (TICV) (Duffus et al., 1996b; Wisler et al., 1998). They act also in Cucumber yellows virus (CuYV) transmission which causes a yellowing disease on cucumber (Cucumis sativus L.) and melon (C. melo L.) (Yamashita et al., 1979; Hartono et al., 2003; Lourencao et al., 2008). Moreover, T. vaporariorum also transmit some other known whitefly-transmitted yellowing viruses i.e., Lettuce infectious yellows virus (LIYV) (Duffus et al., 1986), Tomato chlorosis virus (ToCV) (Wisler et al., 1998), Sweet potato chlorotic stunt virus (SPCSV) (Pio-Ribeiro et al., 1996), Cucurbit yellow stunting disorder virus (CYSDV) (Celix et al., 1996), Potato yellow vein virus (PYVV) (Salazar et al., 2000) and Lettuce chlorosis virus (LCV) (Duffus et al., 1996a). All those viruses are classified as definitive or tentative members of the genus Crinivirus in the family Closteroviridae (Hartono et al., 2003). Because of this, cultivation of tomatoe, cucumber, lettuce, sweet potatoe and potatoe in highland altitude of West Sumatera should considered at risk.
Biotype determination of Bemisia tabaci based on mtCOI gene sequence data: In order to get an overview of genetic variability of our samples we aligned all 43 sequences using Clustal W software provided at http://www.ebi.ac.uk/. The result is shown in Fig. 2. Clustering analysis successfully differentiated all samples into two main clades. Main clade I was composed from 36 samples mainly collected from low altitude (Padang, Pesisir Selatan and Pasaman Barat) and medium altitude (Solok and 50 Kota except 50K-04).
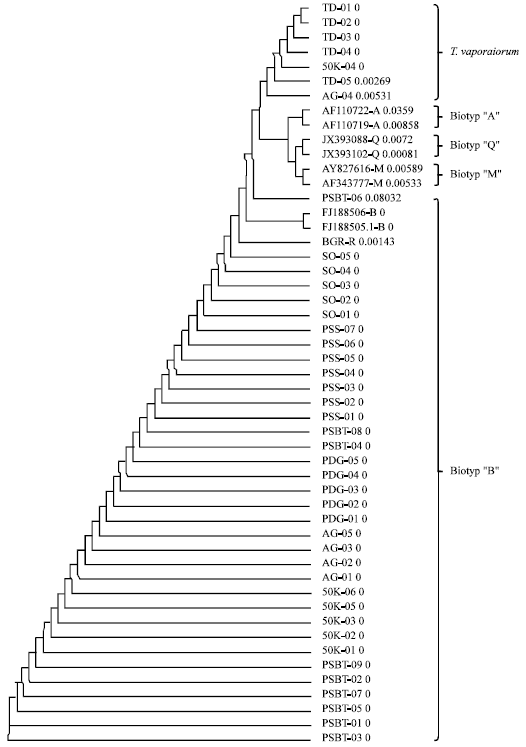 | |
| Fig. 2: | Cladogram of 43 whitefly samples collected from three different geographical regions in West Sumatera. Two representative of each A-biotype (AF110722, AF110719), M (AY827616, AF343777) and Q (JX393088, JX393102) and one biotype B (FJ188505.1) were used as reference for biotype classification |
All samples in the clade I could be definitely identified as B. tabaci. Almost all samples from Agam from high altitude region (except AG-04) were clustered into main clade 1. On the other hand, all samples from Tanah Datar (high altitude) were clustered into main clade II which could be identified as T. vaporariorum.
Another interesting aspect of Bemisia tabacci is their biotype determination. Biotype class determine its compatibility with plant virus pathogen and thus disease management strategy. For this purpose we used two mtCOI nucleotide sequences from each A, B, M and Q-biotype and successively run multialignment analysis by means of Clustal W2. The results are shown in Fig. 2.
All 35 samples identified as B. tabaci were clusterred in main clade I together with FJ186506 and FJ188505 which were identified as B-Biotype. The reference sequence from Bogor BGR-R was also clustered together with both two biotype B. Interestingly sample PSBT-06 collected from Pasaman Barat (lowland region) was more distant from almost all B. tabaci, eventhough it still also clustered into the B-biotype group. The results are in agreement with data published by Hidayat et al. (2004) from tomatoe cultivation area in West Java. However, data obtained here showed different results compared with data published by Shadmany et al. (2013) in Malaysia. Eventhough West Sumatera and Malaysia is not far away from each other geographically. They found Q-biotype population in vegetable and flower producing area.
Data obtained from this study suggested that pest management in both islands must be tighten to control the spreading of other biotypes of B. tabaci. However, attention has to be paid to the fact that B-biotype is highly polyphagous and has spread globally with the trade in ornamentals and vegetables. Moreover, B-biotype is difficult to control since it’s high resistancy to most of the insecticides on the market and it has a wide host range and rapid rate of development and reproduction (Costa et al., 1993; De Barro and Driver, 1997; Qiu et al., 2009).
CONCLUSION
Taking all the data into account the two whiteflies namely Bemisia tabacci and Trialeurodes vaporariorum exist in the Capscium annuum production centre in West Sumatera. So far, this is the first report describing the occurence of Trialeurodes vaporariorum in addition to Bemisia tabacci in chilli pepper cultivation centre in West Sumatera-Indonesia. Biotype determination of B. tabaci based on mtCOI gene sequence showed that so far only B-biotype exists in West Sumatera.
ACKNOWLEDGMENT
We gratefully thank the Directorate of General of Higher Education, Indonesia which has supported this research via scheme of International Research Collaborration and Publication, contract number: 061/SP2H/PL/DIT.LITABMAS/IV/2013, fiscal year 2013. We thank also to DAAD (German Academic Exchange Service) via IGN-TTRC (Indonesia German Network for Teaching, Training and Research Collaboration) for supporting this collaborration. Some parts of this research was also supported by Agricultural Research and Development Agency, from fiscal year 2012.
REFERENCES
- Altschul, S.F., W. Gish, W. Miller, E.W. Myers and D.J. Lipman, 1990. Basic local alignment search tool. J. Mol. Biol., 215: 403-410.
CrossRefPubMedDirect Link - Brown, J.K. and J. Bird, 1992. Whitefly-transmitted geminiviruses and associated disorders in the Americas and the Caribbean Basin. Plant Dis., 76: 220-225.
CrossRef - Celix, A., A. Lopez-Sese, N. Almarza, M.L. Gomez-Guillamon and E. Rodriguez-Cerezo, 1996. Characterization of cucurbit yellow stunting disorder virus, a Bemisia tabaci-transmitted closterovirus. Phytopathology, 86: 1370-1376.
Direct Link - Costa, H., J.K. Brown, S. Sivasupramaniam and J. Bird, 1993. Regional distribution, insecticide resistance and reciprocal crosses between the A and B biotypes of Bemisia tabaci. Int. J. Trop. Insect Sci., 14: 255-266.
CrossRef - De Barro, P.J. and F. Driver, 1997. Use of RAPD PCR to distinguish the B biotype from other biotypes of Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae). Aust. J. Entomol., 36: 149-152.
CrossRefDirect Link - Dinsdale, A.B., L. Cook, C. Riginos, Y.M. Buckley and P.J. De Barro, 2010. Refined global analysis of Bemisia tabaci (Hemiptera: Sternorrhyncha: Aleyrodoidea: Aleyrodidae) mitochondrial cytochrome oxidase 1 to identify species level genetic boundaries. Ann. Entomol. Soc. Am., 103: 196-208.
CrossRefDirect Link - Duffus, J.E., R.C. Larsen and H.Y. Liu, 1986. Lettuce infectious yellows virus-a new type of whitefly-transmitted virus. Phytopathology, 76: 97-100.
Direct Link - Duffus, J.E., H.Y. Liu, G.C. Wisler and R. Li, 1996. Lettuce chlorosis virus: A new whitefly-transmitted closterovirus. Eur. J. Plant Pathol., 102: 591-596.
CrossRef - Duffus, J.E., H.Y. Liu and G.C. Wisler, 1996. Tomato infectious chlorosis virus: A new clostero-like virus transmitted by Trialeurodes vaporariorum. Eur. J. Plant Pathol., 102: 219-226.
CrossRef - Erdogan, C., A.S. Velioglu, M.O. Gurkan, I. Denholm and G.D. Moores, 2012. Chlorpyrifos ethyl-oxon sensitive and insensitive acetylcholinesterase variants of greenhouse whitefly Trialeurodes vaporariorum (Westw.) (Hemiptera: Aleyrodidae) from Turkey. Pesticide Biochem. Physiol., 104: 273-276.
CrossRef - Frohlich, D.R., I. Torres-Jerez, I.D. Bedford, P.G. Markham and J.K. Brown, 1999. A phylogeographical analysis of the Bemisia tabaci species complex based on mitochondrial DNA markers. Mol. Ecol., 8: 1683-1691.
CrossRefPubMedDirect Link - Gawel, N.J. and A.C. Bartlett, 1993. Characterization of differences between whiteflies using RAPD-PCR. Insect Mol. Biol., 2: 33-38.
PubMedDirect Link - Goodwin, P.H., B.G. Xue, C.R. Kuske and M.K. Sears, 1994. Amplification of plasmid DNA to detect plant pathogenic mycoplasmalike organisms. Ann. Applied Biol., 124: 27-36.
CrossRef - Gorman, K., G. Devine, J. Bennison, P. Coussons, N. Punchard and I. Denholm, 2007. Report of resistance to the neonicotinoid insecticide imidacloprid in Trialeurodes vaporariorum (Hemiptera: Aleyrodidae). Pest Manage. Sci., 63: 555-558.
CrossRef - Gorsane, F., I. Fekih-Hassen, M.K. Nakhla, H. Fakhfakh, D.P. Maxwell and M. Marrakchi, 2004. Molecular evidence of Tomato yellow leaf curl virus-Sicily spreading on tomato, pepper and bean in Tunisia. Phytopathol. Mediterr., 43: 177-186.
Direct Link - Hartono, S., T. Natsuaki, Y. Genda and S. Okuda, 2003. Nucleotide sequence and genome organization of Cucumber yellows virus, a member of the genus Crinivirus. J. Gen. Virol., 84: 1007-1012.
CrossRefDirect Link - Horowitz, A.R., S. Kontsedalov, V. Khasdan and I. Ishaaya, 2005. Biotypes B and Q of Bemisia tabaci and their relevance to neonicotinoid and pyriproxyfen resistance. Arch. Insect Biochem. Physiol., 58: 216-225.
CrossRefPubMedDirect Link - Jones, D.R., 2003. Plant viruses transmitted by whiteflies. Eur. J. Plant Pathol., 109: 195-219.
CrossRefDirect Link - Khasdan, V., I. Levin, A. Rosner, S. Morin, S. Kontsedalov, L. Maslenin and A.R. Horowitz, 2005. DNA markers for identifying biotypes B and Q of Bemisia tabaci (Hemiptera: Aleyrodidae) and studying population dynamics. Bull. Entomol. Res., 95: 605-613.
CrossRefDirect Link - Lourencao, A.L., A.C. Alves, C.G. Fugi and E.S. Matos, 2008. Outbreaks of Trialeurodes vaporariorum (West.) (Hemiptera: Aleyrodidae) under field conditions in the State of Sao Paulo, Brazil. Neotrop. Entomol., 37: 89-91.
CrossRef - Ma, D., K. Gorman, G. Devine, W. Luo and I. Denholm, 2007. The biotype and insecticide-resistance status of whiteflies, Bemisia tabaci (Hemiptera: Aleyrodidae), invading cropping systems in Xinjiang Uygur Autonomous Region, Northwestern China. Crop Prot., 26: 612-617.
CrossRef - Martin, J.H., D. Mifsud and C. Rapisarda, 2000. The whiteflies (Hemiptera: Aleyrodidae) of Europe and the Mediterranean Basin. Bull. Entomol. Res., 90: 407-448.
CrossRef - Mugiira, R.B., S.S. Liu and X. Zhou, 2008. Tomato yellow leaf curl virus and Tomato leaf curl Taiwan virus invade South-east coast of China. J. Phytopathol., 156: 217-221.
CrossRef - Pascual, S. and C. Callejas, 2004. Intra-and interspecific competition between biotypes B and Q of Bemisia tabaci (Hemiptera: Aleyrodidae) from Spain. Bull. Entomol. Res., 94: 369-375.
CrossRefDirect Link - Perring, T.M., A.D. Cooper, R.J. Rodriguez, C.A. Farrar and T.S. Bellows Jr., 1993. Identification of a whitefly species by genomic and behavioral studies. Science, 259: 74-77.
CrossRefPubMedDirect Link - Polston, J.E., A. Al-Musa, T.M. Perring and J.A. Dodds, 1990. Association of the nucleic acid of squash leaf curl geminivirus with the whitefly Bemisia tabaci. Phytopathology, 80: 850-856.
Direct Link - Qiu, B., Y. Chen, L.L.W. Peng, X. Li and M.Z. Ahmed et al., 2009. Identification of three major Bemisia tabaci biotypes in China based on morphological and DNA polymorphisms. Prog. Natural Sci., 19: 713-718.
CrossRef - Salazar, L.F., G. Muller, M. Querci, J.L. Zapata and R.A. Owens, 2000. Potato yellow vein virus: Its host range, distribution in South America and identification as a crinivirus transmitted by Trialeurodes vaporariorum. Ann. Applied Biol., 137: 7-19.
CrossRef - Scott, I.A.W., P.J. Workman, G.M. Drayton and G.M. Burnip, 2007. First record of Bemisia tabaci biotype Q in New Zealand. N. Z. Plant Prot., 60: 264-270.
Direct Link - Shadmany, M., D. Omar and R. Muhamad, 2013. First report of Bemisia tabaci (Hemiptera: Aleyrodidae) Biotype Q in Malaysia. Florida Entomol., 96: 280-282.
CrossRefDirect Link - Simon, C., F. Frati, A. Beckenbach, B. Crespi, H. Liu and P. Flook, 1994. Evolution, weighting and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am., 87: 651-701.
Direct Link - Thompson, J.D., D.G. Higgins and T.J. Gibson, 1994. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res., 22: 4673-4680.
CrossRefPubMedDirect Link - Von Dohlen, C.D. and N.A. Moran, 1995. Molecular phylogeny of the homoptera: A paraphyletic taxon. J. Mol. Evol., 41: 211-223.
CrossRef - Wisler, G.C., J.E. Duffus, H.Y. Liu and R.H. Li, 1998. Ecology and epidemiology of whitefly-transmitted closteroviruses. Plant Dis., 82: 270-280.
Direct Link - Hartono, S. and A. Wijonarko, 2007. [Molecular characterization of tomatoe infectious chlorosis virus, a causing agent of tomatoe yellow disease in Indonesia]. Jurnal Akta Agrosia Special Edition, 2: 139-146.
Direct Link - Meliansyah, R., S.H. Hidayat and K.H. Mutaqin, 2011. Geminiviruses associated with the weed species, Ageratum conyzoides, Centipeda minima, Porophyllum ruderale and Spilanthes iabadicensis from Java, Indonesia. Microbiol. Indones., 5: 120-124.
CrossRefDirect Link