
S.K. Akator
Ecole Superieure d�Agronomie (ESA) Universite de Lome B.P. 1515, Togo
D.K. Adjata
Centre du Riz pour l�Afrique (AfricaRice) 01 B.P. 2031, Cotonou, Benin
S. Drissa
Centre du Riz pour l�Afrique (AfricaRice) 01 B.P. 2031, Cotonou, Benin
S. Awande
Centre du Riz pour l�Afrique (AfricaRice) 01 B.P. 2031, Cotonou, Benin
L. Zadji
Centre du Riz pour l�Afrique (AfricaRice) 01 B.P. 2031, Cotonou, Benin
G. Sangare
Centre du Riz pour l�Afrique (AfricaRice) 01 B.P. 2031, Cotonou, Benin
Y. Sere
Centre du Riz pour l�Afrique (AfricaRice) 01 B.P. 2031, Cotonou, Benin
Y.M.D. Gumedzoe
Ecole Superieure d�Agronomie (ESA) Universite de Lome B.P. 1515, Togo
Asian Journal of Plant Pathology
Year: 2014 | Volume: 8 | Issue: 1 | Page No.: 10-17







ABSTRACT
Study for the genetic diversity of P. oryzae populations, under natural inoculation, trapping of virulence races present in ecosystems, was done during the period of 2008 to 2010 in two countries. One location/country was surveyed to highlight that resistance genes: Pi1; Pi11; Pi12; Pi19; Pib; Pi20; Pi5; Pi7; Pia; Pia+Pi19; Pia+Pish; Pif, Pii, Pik, Pik-p, Pish, Pit, Pit, Pita-2, Piz, Piz-t, Piks at M'be and and those at Ouedeme: Pi1, Pi12, Pi19, Pi1b, Pi3, Pi5, Pi7, Pia, Pii, Pik, Pi-p, Pit, Pita, Piks incidence. It was shown that they were overcome by a large proportion of the virulence gene of P. oryzae population. However, virulence genes that are capable of overcoming resistance genes: Pi33, Pi5 (t), Pi7, Pi9, Pikh+Pi-1+Pita+Pita, Pik-m ,Pish, Pitta-2, Piz-5, were absent or rare. These genes were effective against the pathogen population studied. This study also showed that Pi5 and Pi7, Pikh +Pi-1+Pita+Pita, genes association which were individually inefficient, has conferred a sustainable resistance to blast that was observed in Moroberekan and Tetep. This work will help rice breeders and plant pathologists to select rice varieties with durable resistance to blast disease.
PDF Abstract XML References Citation
Received: October 28, 2013;
Accepted: January 19, 2014;
Published: March 29, 2014
How to cite this article
S.K. Akator, D.K. Adjata, S. Drissa, S. Awande, L. Zadji, G. Sangare, Y. Sere and Y.M.D. Gumedzoe, 2014. Pathological Studies of Pyricularia oryzae at M’be
in Coted’ivoire and Ouedeme in
Benin. Asian Journal of Plant Pathology, 8: 10-17.
URL: https://scialert.net/abstract/?doi=ajppaj.2014.10.17
URL: https://scialert.net/abstract/?doi=ajppaj.2014.10.17
INTRODUCTION
Upland rice represents about 44% of the total rice production in West Africa. It covers 57% of cultivated rice surface and more than 70% of farmers are engaged for their subsistence (Jones et al., 1997). Rice blast fungus is one the most prevalent and devastating disease of rice in West Africa and in the world (Chen et al., 2004). It is caused by a pathogenic fungus which telemorph stage is called Magnaporthe grisea (Pan et al., 1998) and the anamorph stage is Pyricularia oryzae. Rice cultivation is under a constant pressure of blast of the leaf, node and panicle of the grain causing extensive damage to upland rice, estimated at 21-64% in West Africa (Sere et al., 2004). In South Saharan Africa, the use of resistant varieties associated with improved farming techniques is the only method of control available to smallholder farming among which more than half are women. However, if in the fight against P. oryzae, the achievements of agricultural research in the field of chemotherapy are significant in developed countries (Sy et al., 1994), rice blast caused by the fungus M. oryzae was one of the most serious diseases of rice and remains one of the most difficult diseases to manage (Khush and Jena, 2007; Jia and Moldenhauer, 2010).
Rice farmers in South Saharan Africa are not able to purchase fungicides; the use of resistant varieties is the only effective weapon they have. The only control method available (Van der Plank, 1968), however, faces several obstacles, including the variability of the pathogenicity of the causal agent of the disease, P. oryzae (Pan et al., 1998). Brown (2002) showed that the resistance genes to blast and the rice yield genes are inversely related, the selection of resistant cultivars to blast would be a penalty for the selection of high-yielding cultivars. The objective of this work is to study the genetic diversity of P. oryzae populations under natural inoculation at M'Be and Ouedeme in Benin and Côte d’Ivoire.
MATERIALS AND METHODS
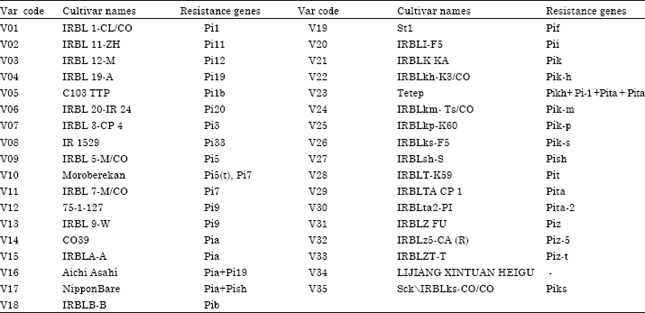
Experimental site and Biological materials: These studies were conducted during the period of 2008 to 2010 in the research stations of AfricaRice at M'be in Côte d'Ivoire and at Ouedeme in Benin, in the zone of southern Guinean savanna of West Africa. Soil characteristics of M'be and Ouèdèmè are cosigned in Table 1. The biological materials are composed of 35 strains and varieties mono or oligo gene (Table 2) increased in the absence of pressure blast and seeds are processed by a non-systemic fungicide.
METHODS
Experimental design: The experimental design is a block of Fischer with 4 repetitions of 3 lines of 50 cm separated from each other by 10 cm and a line between the access to facilitate their identification and evaluation.
| Table 1: | Description of the properties of the soils of Mbé and Ouèdèmè |
 | |
| Table 2: | List of biological material |
 | |
The soil was plowed with a disc plow twice to facilitate the upgrade with a harrow. Fertilization composed fertilizer NPK 10-18-18 was applied at 200 kg ha-1 before sowing. Urea was given to 21 and 42 days after sowing. Sowing was done by putting 3-4 seeds per hole at 10 cm spacing between the holes and 10 cm between rows. The seedlings were reduced to one plant per hole at 14 days later with eventual replacement of missing plants.
Pathological infection: Growing plants were left for natural infection. Pathological symptoms and severity were recorded according to the IRRI standard evaluation scale for rice (IRRI, 1980). Degree of severity was taken in consideration as shown in Table 3. Screening was done for each variety as well as positive and negative control using the same IRRI standard evaluation scale (IRRI, 1980).
Evaluation of host reaction and statistical analysis: Leaf blast was assessed at the 21, 28, 35, 42, 49, 56, 63 and 70 DAS by using the Standard Evaluation Scale (SES) of IRRI (1980) (Table 3). The notes were converted into sick leaf surface and the data were analyzed by using the software XLSTAT 2010 site by site.
RESULTS
Evaluation of host reaction and analysis: Statistical data analysis by XLTAT 2010 gives the correlation matrices (Table 4 and 5), the distribution graph lines according to their resistance/susceptibility to blast (Fig. 1 and 3), the dendrograms and classes of resistant and susceptible lines (Fig. 2 and 4).
| Table 3: | Standard evaluation scale of rice blast disease |
 | |
| Source: IRRI (1980) | |
| Table 4: | Matrice of correlation (Pearson (n)) at M’bé |
 | |
| Table 5: | Matrice of correlation (Pearson (n)) at Ouèdèmè |
 | |
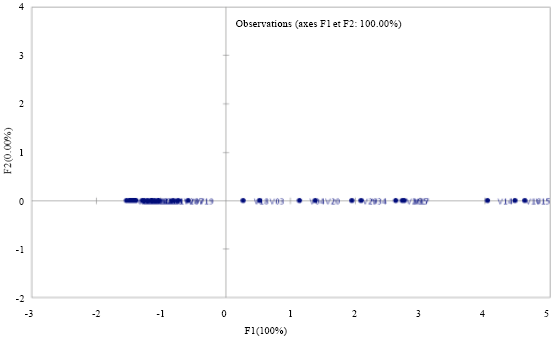 | |
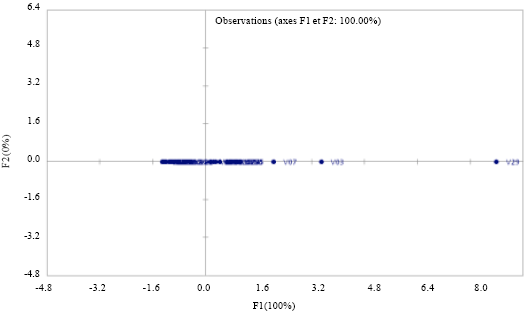
| Fig. 1: | Graph of distribution lines on the first axis of the Principal Component Analysis (PCA) at M’bè |
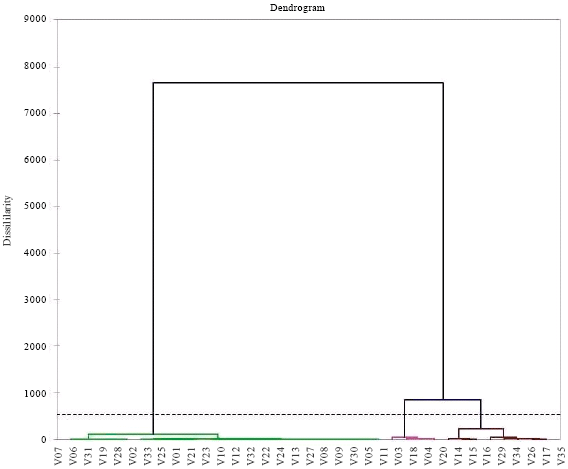 | |
| Fig. 2: | Dendrogram lines according to their reaction to P. oryzae at M’bè |
 | |
| Fig. 3: | Graph of distribution lines on the first axis of the Principal Component Analysis (PCA) at uèdèmè |
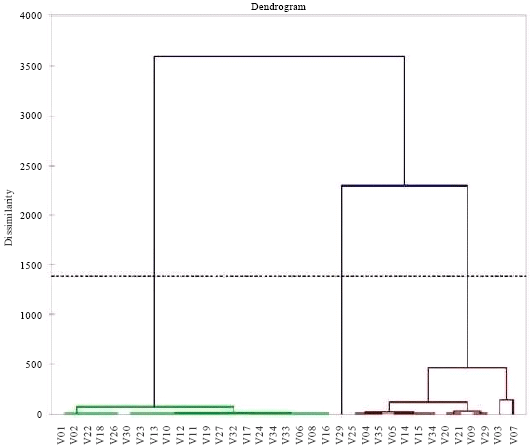 | |
| Fig. 4: | Dendrogram lines according to their reaction to P. oryzae at Ouèdèmè |
This correlation is 75.1 to 87.6% in Ouedeme (Benin), while in M'Be (Côte d’Ivoire) it ranges from 76.6 to 99.3%.
The principal component analysis: The Principal Component Analysis (PCA) and the numerical classification results (Fig. 3) showed that the first two principal components explain at 100% the reaction of the varieties and lines to blast fungus. The lines are distributed as follows: The sensitive lines are at the positive side of the axis and the resistant lines are at the negative side of the axis.
Reaction of the different cultivars and lines in relation to blast: The two dendrograms (that of M'Bé (Côte d’Ivoire) and of Ouedeme (Benin)) share out the lines and varieties into two classes. Resistant varieties with Sick Foliar Surface is equal to or less than 1.5% are: At M'be (Côte d’Ivoire): V10, V12, V13, V22, V23, V24, V27, V30, V32 and at Ouèdèmè (Benin): V02, V06, V08, V10, V12, V13, V17, V18, V19, V22, V23, V24, V30, V32. These lines are followed by the susceptible varieties. Thus, at M'be, there are: V01, V02, V03, V04, V05, V06, V07, V09, V11, V14, V15, V16, V17, V18, V19, V20, V21, V25, V26, V28, V29, V31, V33, V34, V35 and at Ouèdèmè, there are: V01 V03 V04 V05 V07 V09 V11 V14 V15 V20 V21 V25 V28 V29 V34 V35.
DISCUSSION
At the both sites, the Principal Component Analysis (PCA) and numerical classification notes collected 49, 56, 63, 70 days after sowing and transformed in Foliar Surface Sick, categorizes the 35 varieties in two distinct classes: the class of the resistant varieties and that of the susceptible ones Fig. 3 and 4. The resistant varieties have been able to react against virulence genes in populationsof P. Oryzae at both sites (Table 4 and 5). PCA analysis and numerical classification show that there is great variability between different varieties and lines at the both sites. These genes at the both sites are: Pi5 (t) Pi7 Moroberekan, Pi9 of 75-1-127, Pik-h of IRBLKH-K3, Pikh + Pi-1+Pita+Pita of Tetep, Pik-m of IRBLkm-TS 5, Piz-5 of IRBLZ-CA (R), Pi-9 of IRBL 9-W, Pita-2 of IRBL TA 2-PI, Pi33 of IR 1529 and Pish of IRBLsh-S which are resistant on the two sites. The observation of the behavior of varieties and lines has allowed to show that genes Pikh, Pi-1, Pita, Pi5 (t) are individually Pi7 ineffective whereas their association within Moroberekan (Pi5 (t) Pi7) and Tetep (pikh Pi-1+Pita+Pita) provides high resistance. That means the varieties evaluated exhibited differential but characteristic responses to rice blast fungus as it has been reported earlier (Odjo et al., 2011).
The contrasts of the two locations were unexpected at least to the extent and magnitude observed, because the two sites were in the same agro-ecological zones and is expected to share similar macroclimatic conditions. Nevertheless, the differences suggest that site specific microclimatic factors and the variability associated with them are more in blast screening than apparent macroclimatic factors in the choice of site for blast screening. Furthermore, there may be possibility of the presence of different strains of blast fungus at the two sites since the genotypes used in this study were special varieties, each carrying specific known resistant genes. According to Sere et al. (2004) it is important that the performance of a variety be viewed in relation to the biotic and abiotic parameters of the environment (or precisely of the site and ecological niches) that have been targeted. Such an effect of several genes in a variety explains the interest of pyramiding resistance genes to confer a high level of resistance to blast. The usefulness of pyramiding resistance genes to blast has been demonstrated experimentally (Hittalmani et al., 2000). Such associations also provide the benefit of a durable type of resistance difficult to overcome in time and space by the virulence gene; case of Pi5 (t) and Pi7 Pikh Pi-1 + Pita + Pita and it is established that they are easily circumvented by the pathogen (Conaway-Bormans et al., 2003). This work also confirms the work of Roder et al. (1998) in which he showed that resistance genes Pi2 and Pi1 had a spectra of additional resistance and the accumulation of these genes in a variety could confer resistance to broad-spectrum so potentially sustainable.
In the statistical model used for this study, although, there were significant effects for all main factors and interaction effects; it is interesting to note that among the known sources of variation, Genotype by Environment Interaction (GEI) consistently had highest contribution to the total sum of squares for all levels of disease development and severity scores except at 70 days after sowing. For growth traits, however, variety had the highest contribution to total sum of squares. This is an indication that blast disease development and severity scores were greatly conditioned by environmental factors while growth traits were controlled by the genotypic effect of the variety (Van der Plank, 1968). This result is expected because the evaluation was carried out under a natural infection conditions on the field (Jia and Moldenhauer, 2003). Different trends may be observed if evaluation has been done in the screen house conditions or under artificial inoculation on the field. The pattern of disease severity differed at the two locations. At M’Be (Cote d’Ivoire), the difference between resistant and susceptible varieties were not significant until about 70 days after planting while difference became significant by around 78 days at Ouedeme (Benin). This could be explained by the fact that rice was cultivated at M'Be since the 1970s while at Ouèdèmè it just started in 2002.
In conclusion, the study the genetic diversity of P. oryzae populations under natural inoculation by using trapping design of virulence races of fungus in ecosystems of M'bé and Ouèdèmè led to the study of the genetic diversity of populations of P. oryzae. It has been proven that any pathogen resistance gene is a virulence gene of the host plant. It is clear from this work that the resistance genes of the cultivars are overcome by a significant proportion of virulence genes of the explored experimental sites. Nevertheless, the virulence genes capable of overcoming the resistance genes Pi9 of the isogenic lines 75-1-127, Pi5 (t) of Pi7 Moroberekan, Pikh Pi-1+Pita+Pita of Tetep, Pik-m of IRBLKM TS and Piz-5 of IRBLZ 5-CA are absent or rare in the two sites. These are effective genes from this experiment. These genes could be associated two or three in the same variety to ensure the sustainability of rice resistance to blast. In fact, such associations are effective in the case of Pi5 and Pi7 and it is established that they are difficult to overcome by pathogens.
ACKNOWLEDGMENTS
The authors wish through the Head Unit Pathology of AfricaRice Center, Dr Yacouba Sere, to express their gratitude to IRRI and CIRAD for the isogenic lines.
REFERENCES
- Brown, J.K., 2002. Yield penalties of disease resistance in crops. Curr. Opin. Plant Biol., 5: 339-344.
PubMed - Chen, X.W., S.G. Li, J.C. Xu, W.X. Zhai and Z.Z. Ling et al., 2004. Identification of two blast resistance genes in a rice variety, digu. J. Phytopathol., 152: 77-85.
CrossRef - Conaway-Bormans, C.A., M.A. Marchetti, C.W. Johnson, A.M. McClung and W.D. Park, 2003. Molecular markers linked to the blast resistance gene Pi-z in rice for use in marker-assisted selection. Theor. Applied Genet., 107: 1014-1020.
CrossRef - Roder, M.S., V. Korzun, K. Wendehake, J. Plaschke, M.H. Tixier, P. Leroy and M.W. Ganal, 1998. A microsatellite map of wheat. Genetics, 149: 2007-2023.
PubMedDirect Link - Hittalmani, S., A. Parco, T.V. Mew, R.S. Zeigler and N. Huang, 2000. Fine mapping and DNA marker-assisted pyramiding of the three major genes for blast resistance in rice. Theor. Applied Genet., 100: 1121-1128.
CrossRefDirect Link - Jia, Y. and K. Moldenhauer, 2010. Development of monogenic and digenic rice lines for blast resistance genes Pi-ta, Pi-kh/Pi-ks. J. Plant Registrations, 4: 163-166.
Direct Link - Jones, M.P., M. Dingkuhn, G.K. Aluko and M. Semon, 1997. Interspecific Oryza sativa L. X O. Glaberrima steud. Progenies in upland rice improvement. Euphytica, 94: 237-246.
CrossRefDirect Link - Pan, Q., L. Wang, T. Tanisaka and H. Ikehashi, 1998. Allelism of rice blast resistance genes in two Chinese rice cultivars and identification of two new resistance genes. Plant Pathol., 47: 165-170.
CrossRef - Sere, Y., A.A. Sy, S.K. Akator, A. Onasanya and K. Zai, 2004. Screening Strategy for Durable Resistance to Rice Blast Disease at WARDA. In: Rice Blast in West Africa: Characterisation of Pathogen Diversity, Key Screening Sites and Host Resistance, Sere, Y., S. Sreenivasaprasad and S.K. Nutsugah (Eds.). Vol. 4. Africa Rice Center (WARDA), Benin, Nigeria, pp: 38-43.
Direct Link