
Nabil S. Farag
Plant Pathology Research Institute, ARC, Giza, Egypt
Naglaa M.A. Balabel
Potato Brown Rot Project (PBRP), ARC, Dokki, Egypt
Asian Journal of Plant Pathology
Year: 2014 | Volume: 8 | Issue: 1 | Page No.: 1-9







ABSTRACT
The possible carryover effect of three fungicides and one insecticide on results of PCR and Multiplex-PCR for detection of bacterial pathogens was studied. The potato brown rot caused by phylotype II, sequevar I of Ralstonia solanacearum and the potato ring rot bacteria Clavibacter michiganensis ssp. sepedonicus were considered in this study. Under optimized conditions for PCR activity, the potato extract prepared from the Cara potato cultivar did not interfere with band development of DNA’s extracted from the organisms in concern. Regardless of their mode(s) of action, the studied pesticides exerted different effect(s) on band development according to their chemical grouping. The dimethomorph (Acrobat), systemic fungicide, caused complete inhibition of band development of DNA extracted from either potato brown rot or potato ring rot bacteria which may be considered as false negative reaction(s). The propamocarb carbamate (Previcur N), systemic fungicide, did not inhibit band development of the organisms in concern. The dithiocarbamate plus zinc and manganese (Diathane M45), contact fungicide however, caused a complete band inhibition of ring rot DNA contrary to that being reported for brown rot, indicating a preferential polymerase activity. The organophosphate (Malathion), penetrative non-systemic insecticide, did not inhibit DNA band development for both organisms. Multiplex-PCR of mixed DNA’s of R. solanacearum and C. michiganensis ssp. sepedonicus caused a false negative reaction through inhibition of band development of R. solanacearum but not for C. michiganensis ssp. sepedonicus, under the effect of propamocarb carbamate (Previcur N) fungicide and organophosphate (Malathion) insecticide. It could be concluded that the pesticides carryover effect on PCR results of potato brown rot and ring rot bacteria are being variable according to the pesticide grouping. The dimethomorph (Acrobat) fungicide caused false negative effect with both organisms under study followed by dithiocarbamate (Diathane) fungicide in case of ring rot bacteria. The false negative multiplex-PCR is being evident only in case of R. solanacearum under the stress of both propamocarb carbamate (Previcur N) fungicide and on the organophosphate (Malthion) insecticide. The mode of action of the used pesticide, either contact or systemic, had no effect on PCR or multiplex-PCR results. Further investigations are needed with different groups of pesticide.
PDF Abstract XML References Citation
Received: September 10, 2013;
Accepted: January 22, 2014;
Published: March 29, 2014
How to cite this article
Nabil S. Farag and Naglaa M.A. Balabel, 2014. Effect of Pesticide(s) Carryover on Results of Polymerase Chain Reaction
and Multiplex-PCR for Diagnosis of Potato Bacterial Pathogens. Asian Journal of Plant Pathology, 8: 1-9.
URL: https://scialert.net/abstract/?doi=ajppaj.2014.1.9
URL: https://scialert.net/abstract/?doi=ajppaj.2014.1.9
INTRODUCTION
Potato ring rot caused by Clavibacter michiganensis ssp. sepedonicus and potato brown rot caused by Ralstonia solanacearum are tuber-borne quarantine diseases (Kudela, 2007; Kabeil et al., 2008). The latent infection produced by these bacterial pathogen, under certain circumstances, has complicated the usual quarantine inspection decision, hence the dependency on more sophisticated laboratory methods have been described (Caruso et al., 1998). Among these protocols, polymerase chain reaction, PCR and its variant multiplex-PCR were tried (Seal et al., 1993; Farag et al., 2010). The multiplex-PCR as a modification of the original PCR testing is subject to certain reaction difficulties as PCR drift and PCR selection (Wagner et al., 1994) in addition to other difficulties related to the availability of primers for various plant pathogens (Martin et al., 2000; Schaad and Fredrick, 2002), the formation of primer dimer (Brownie et al., 1997) and the carryover of certain inhibitory factors in the samples (Rossen et al.,1992; Wilson, 1997). The most inhibitory realized factors being plant polysaccharides (Demeke and Adams, 1992), humic acid in plant and soil (Tsai and Olson, 1992; Watson and Blackwell, 2000) and certain groups of antibiotics (Farag et al., 2010). Therefore, the correct sampling and optimization of reaction conditions must be strictly considered to minimize such non-specific reactions and avoid false results (De Lomas et al., 1992; Polz and Cavanaugh, 1998).
The concept of this study stems from the fact that potato raised in the temperate parts of the world is subject to an array of foliar disease and insect attacks that are being routinely controlled with pesticides (Samoucha and Cohen, 1990; Powelson and Inglis, 1999; Gafar and Dagash, 2011).
The present study aimed to determine the possible carryover effect of potato pesticide (s), with different mode (s) of action, on the results of multiplex-PCR for detection of bacterial pathogens.
MATERIALS AND METHODS
Optimization of reaction conditions: The entire adjustment of multiplex-PCR conditions is carried out with consideration of the following:
Bacterial pathogens: Bacterial cultures of the targeted pathogens and their origin are shown in Table 1. The Bacterial Ring Rot (BRR) pathogen was grown on yeast extract glucose mineral salt agar, YGMA, [g:L Bacto-Yeast-Extract (Difco)2.0; D (+) Glucose (monohydrate) 2.5; K2HPO4 0.25; KH2PO4 0.25; MgSO4.7H2O 0.1; MnSO4.H2O 0.015; NaCl 0.05; FeSO4.7H2O 0.005; Bacto-Agar (Difco) 18] at 20°C (Anonymous, 1993). The bacterial brown rot (BBR) was grown on Kelman’s Tetrazolium medium [g:LCasamino acids (Difco) 1.0; Bacto-Peptone (Difco) 10.0; Dextrose 5.0; Bacto-Agar (Difco) 15.0, after sterilization and Cooling to 50°C, a filter-sterilized solution of 2,3,5-triphenyl tetrazolium chloride (Sigma) is added to obtain a final concentration of 50 mg L-1, (Kelman, 1954).
Primers: The sequence of primers used in this study is shown in Table 2. The primers were provided by OPRON, USA.
| Table 1: | Source of bacterial pathogens and disease(s) produced |
 | |
| BRR: Bacterial ring rot, BBR: Bacterial brown rot, *PD: Plant protection service, Wageningen NL., **PBRP: Potato brown rot project, Dokki, Egypt | |
| Table 2: | Sequence of primers used with BRR and BBR (phylotype II, sequevar I, i.e., race 3, biovar 2) |
 | |
| Table 3: | Name of pesticides used with multiplex-PCR |
 | |
Procedure: Multiplex-PCR amplification was run by PTC-200 Peltier (USA) thermal cycler for primers listed in Table 2. The reaction mixture (25.0 μL) consisted of 12.5 μL Qiagen Taq PCR master mix, 2 μL of each of solanacearum primers and 2 μL of sepedonicus primers along with 2 μL of DNA extract. Supplement of pesticides in potato extract was added separately as 2.5 μL solution to the reaction mixture and being replaced by the same quantity of sterile UPW in the check treatment.
The initial denaturation was made at 96°C for 3 min followed by 35 reaction cycles at 94°C for 30 sec, then 52°C for 30 sec and 72°C for 30 sec. The mixture was then kept at 72°C 10 min-1 and stored at 4°C. Aliquots (13 μL) of the reaction mixture were resolved by electrophoresis in agarose gel (2%) and DNA fragments were visualized by ethidium bromide (o.5% μg mL-1).
Potato extract: Potato extracts were made according to the protocols of the European Union Directive No. 98/57 EC and routinely used in Potato Brown Rot Project (PBRP), Egypt. Thin minced slices of the stolon ends of tubers (200 tubers) for processing were extracted in phosphate buffer [4.26 g of Na2HPO4 and 2.72 g of KH2PO4 (50 mM PO4) L-1 of distilled water, pH 7.0]. The sample was shaken (100 rpm) under controlled temperature (15°C) for 4 h. Potato extract was used in different pesticide treatments.
Pesticides: The carryover effect of a group of fungicides and widely used insecticide at 1000 ppm of the commercial products on the results of multiplex-PCR was determined. Table 3 shows the trade name, active component and mode of action.
RESULTS
Multiplex-PCR optimization: Under the afore-mentioned conditions of the experiment the bands of R. solanacearum (phylotype II, sequevar I), previously called potato race 3, biovar 2, were recognized at 400 bp and those for C. michiganensis ssp. sepedonicus at 200 bp, being compatible with the master mix bands (Fig. 1). The figure also shows that potato extract cv. Cara did not interfere with band development for DNA extracted from either R. solanacearum and C. michiganensis ssp. sepedonicus (Fig. 1).
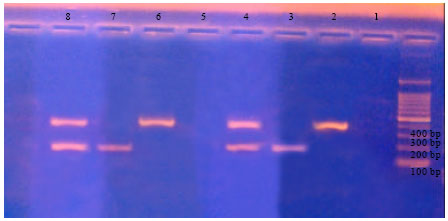 | |
| Fig. 1: | Potato extract did not affect band development; Treatments from 6 to 8 with potato extract; Treatments from 2 to 4 without potato extract; Negative controls (1 and 5); 2 = R. solanacearum without potato extract; 3 = C. michiganensis ssp. sepedonicus without potato extract; 4 = R. solanacearum+C. michiganensis ssp. sepedonicus without potato extract; 6 = R. solanacearum with potato extract; 7 = C. michiganensis ssp. sepedonicus with potato extract; 8 = R. solanacearum+C. michiganensis ssp. sepedonicus with potato extract |
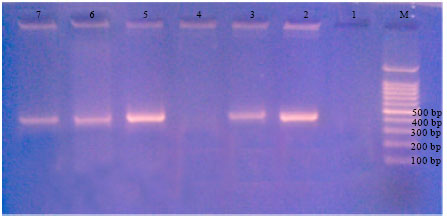 | |
| Fig. 2: | Effect of pesticides on band development for R. solanacearum at 400 bp; M = Molecular size of marker (100 bp DNA); 1 = Negative control, 2 = Positive control, 3 = R. solanacearum+potato extract, 4 = R. solanacearum+potato extract+Acrobat, 5 = R. solanacearum+potato extract+Previcur N, 6 = R. solanacearum+potato extract+Diathane, 7 = R. solanacearum+potato extract+Malathion. Complete inhibition with acrobat pesticide (treatment 4) |
Effect of pesticide(s) carryover on multiplex-PCR results: The possible carryover effect of fungicides used for control of the fungal pathogens, along with an insecticide used for control of potato aphids on multiplex-PCR results, are shown in Fig. 2-4.
Figure 2 shows that the carbamate fungicides, regardless of their mode of action, either contact or systemic, did not affect band development for DNA extracted from R. solanacearum.
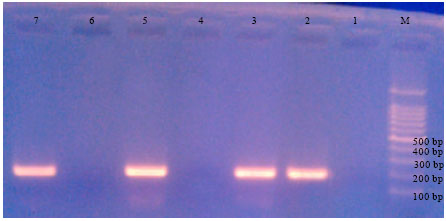 | |
| Fig. 3: | Effect of pesticides on band development for C. michiganensis ssp. sepedonicus at 200 bp; M = molecular size marker (100 bp DNA), 1 = Negative control, 2 = Positive control, 3 = C. michiganensis ssp. sepedonicus +potato extract, 4 = C. michiganensis ssp. sepedonicus+potato extract+Acrobat, 5 = C. michiganensis ssp. sepedonicus+potato extract+Previcur N, 6 = C. michiganensis ssp. sepedonicus+potato extract+Diathane, 7 = C. michiganensis ssp. sepedonicus+potato extract+Malathion. Complete inhibition with Acrobat and Diathane pesticides (treatment 4, 6) |
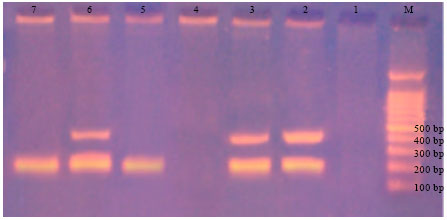 | |
| Fig. 4: | Effect of pesticides on band development for R. solanacearum+C. michiganensis ssp. sepedonicus at 400, 200 bp; M = Molecular size marker (100-bp DNA), 1 = Negative control, 2 = Positive control, 3 = R. solanacearum+C. michiganensis ssp. sepedonicus+potato extract, 4 = R. solanacearum +C. michiganensis ssp. sepedonicus+potato extract+Acrobat, 5 = R. solanacearum+C. michiganensis ssp. sepedonicus+potato extract+Previcur N, 6 = R. solanacearum+C. michiganensis ssp. sepedonicus+potato extract+Diathane, 7 = R. solanacearum+C. michiganensis ssp. sepedonicus+potato extract+Malathion. Complete inhibition with acrobat pesticide (treatment 4) |
The propamocarb carbamate (systemic, previcur N,) and the bisdithiocarbamate (contact, diathane,) did not inhibit band development. The dimethomorph group (acrobat, systemic), however, caused a complete inhibition of band development.
| Table 4: | Summary on the effect of pesticides on the two organisms |
 | |
| *Ralstonia solanacearum, **Clavibacter michiganensis ssp. sepedonicus, +: No inhibition, -: Inhibition of bands | |
Figure 2 also shows that the organophosphate insecticide (penetrative but non systemic, malathion) did not inhibit band development for DNA of R. solanacearum.
With regard to the effect of fungicides carryover on C. michiganensis ssp. sepedonicus, Fig. 3 shows that the carbamate fungicide has exerted different effect on band development. The systemic propamocarb carbamate (previcur N) did not inhibit band development, contrary to the contact bisdithiocarbamate (diathane) that caused complete inhibition of banding. The systemic dimethomorph group (acrobat) caused complete inhibition of DNA banding as well. The penetrative non-systemic organophosphate insecticide (malathion) did not inhibit band development of sepedonicus DNA which is similar to that being reported for solanacearum DNA extracted.
The combined interaction of DNA’s extracted from both R. solanacearum and C. michiganensis ssp. sepedonicus in multiplex-PCR trails is shown in Fig. 4. The systemic propamocarb carbamate (previcur N), caused, under the conditions of the experiment, an unexpectedly band inhibition for R. solanacearum at 400 bp which is being different from that reported in the usual PCR testing (Fig. 2). Such inversion in result is not evident in the usual PCR trial with C. michiganensis ssp. sepedonicus at 200 bp (Fig. 3).
The contact bisdithiocarbamate (diathane), however, did not inhibit banding of DNA of the mixed organisms, though the reported inhibitive effect on banding of C. michiganensis ssp. sepedonicus at 200 bp (Fig. 3). The systemic dimethomorph (Acrobat) fungicide has inhibited DNA banding of both organisms under investigation (Fig. 4). The latter effect is similar to that being reported in the usual PCR trials individually for both of them (Fig. 2 and 3).
The non-systemic penetrative organophosphate insecticide (malathion) caused apparent band inhibitions for R. solanacearum and distinct production for C. michiganensis ssp. sepedonicus band at 200 bp was detected (Fig. 4) similar to that being detected Fig. 3.
Table 4 summarizes the effect of pesticides on the both organisms.
DISCUSSION
Early studies on multiplex-PCR have highlighted the obstacles that can jeopardize the production of sensitive and specific multiplex assays but more recent studies have provided systematic protocols and technical improvements for simple test design (Elnifro et al., 2000). Multiplex-PCR as a modification of the original PCR testing is subject to a certain difficulties as PCR drift and PCR selection (Wagner et al., 1994). PCR drift is a bias assumed to be due to fluctuation in the interaction of PCR reagents particularly in the early cycles which could arise in the presence of a very low template concentrations (Dieffenbach et al., 1993; Mutter and Boynton, 1995); variations in the thermal profiles of a thermo cycler, resulting in an unequal ramping temperatures; or simple experimental error. PCR selection on the other hand, is defined as a mechanism which inherently favors the amplification of certain templates, due to the properties of the target, the target flanking sequences, or the entire target genome (Wagner et al., 1994).
The present study aimed to study the effect of the possible carryover of four potato pesticides on the results of PCR and the results of its variant multiplex-PCR for the detection of R. solanacearum and C. michiganensis ssp. sepedonicus.
The results showed that under optimized reaction condition, the potato extract exerted no interference with band development of the brown rot pathogen at 400 bp and the ring rot pathogen at 200 bp, with either PCR or multiplex-PCR. In this regard, Farag et al. (2010) reported that the multiplex-PCR showed a high degree of reliability to detect these bacterial pathogens along with Erwinia carotovora ssp. atroseptica, the causative agent of potato blackleg.
The selected pesticides are of common use on potato worldwide (Samoucha and Cohen, 1990; Powelson and Inglis, 1999; Chaurasia, 2005; Gafar and Dagash, 2011). Regardless, of their mode(s) of action, either contact or systemic, the propamocarb carbamate (previcur N) and bisdithiocarbamate (diathane) did not inhibit the band development of the DNA target of the brown rot pathogen. Under similar conditions, however, the bisdithiocarbamate caused a complete inhibition of DNA band of the other target pathogen i.e., the bacterial ring rot. Therefore, it could be concluded that the carbamate fungicides, according to their chemical nature and the attached side groups, were reacting differently. Such reported differences may be attributed as well to certain factors preventing optimal annealing and/or the extension rates of the polymer in case of ring rot pathogen (Ruano et al., 1991). Therefore, the bisdithiocarbamate (diathane) carryover may interfere with PCR reaction, thus producing false negative results and producing considerable difficulties during ring rot diagnosis. The non-systemic penetrative organophosphate insecticide (malathion) did not inhibit the band development of the targets in concern, indicating no interference with enzyme activity under both circumstances. The systemic dimethomorph fungicide (acrobat), under the conditions of the experiment, caused a complete inhibition of band development of DNAs extracted from either R. solanacearum or C. michiganensis ssp. sepedonicus which may be considered as false negative reaction. The effect may be attributed to direct interference of the fungicide with enzyme activity, due to possible changes in the optimal reaction conditions (Stephenson and Abilock, 2012). In this regard the multiplex-PCR with certain medical bacteria showed 2 false-negative in many positive samples but no false-positive was detected (Kim et al., 2010). It is well established recently that the rate at which primers anneal to their target and the rate at which annealed primers are extended along the desired sequence, during different cycles of the amplification, are among the most important factors controlling the successful detection. Factors preventing optimal annealing rates include poorly designed primers and suboptimal buffer constituents and annealing temperature. Such conditions are not apparently evident in the present study as shown by the positive controls for both organisms (Fig. 1-3). Moreover, the bias in template-to-product ratios may be considered in multiplex-PCRs that are designed to amplify more than one target simultaneously (Walsh et al., 1992; Mutter and Boynton, 1995; Polz and Cavanough, 1998).
In retrospect, it could be concluded that potato extract did not inhibit band development of DNA extracted from R. solanacearum and C. michiganensis ssp. sepedonicus either in PCR or multiplex-PCR. The dimethomorph (Acrobat) fungicide caused a false negative PCR and multiplex-PCR results, for both organisms under investigation. Multiplex-PCR, for mixed DNA’s of R. solanacearum and C. michiganensis ssp. sepedonicus, caused false negative results in propamocarb carbamate (Previcur N) and organophosphate (Malathion) fungicides treatments.
Further investigations are needed to study in greater depth an additional numbers of pesticides practically used with potatoes to overcome the possible carryover effect on PCR and multiplex-PCR results during bacterial disease diagnosis.
REFERENCES
- Caruso, P., P. Llop, J.L. Palomo, P. Garcia, C. Morente and M.M. Lopez, 1998. Evaluation of Methods for Detection of Potato Seed Contamination by Ralstonia Solanacearum. In: Bacterial Wilt Disease: Molecular and Ecological Aspects, Prior, P., C. Allen and J. Elphinstone (Eds.). Springer, Berlin, Germany, pp: 128-132.
- Chaurasia, P.C.P., 2005. Economic management of late blight (Phytophthora infestans L.) of potato in Eastern Tarai of Nepal. Nepal Agric. Res. J., 6: 57-61.
CrossRefDirect Link - De Lomas, J.G., F.J. Sunzeri and M.P Busch, 1992. False-negative results by polymerase chain reaction due to contamination by glove powder. Transfusion, 32: 83-85.
CrossRefPubMedDirect Link - Demeke, T. and R.P. Adams, 1992. The effects of plant polysaccharides and buffer additives on PCR. Biotechniques, 12: 332-334.
PubMed - Dieffenbach, C.W., T.M.J. Lowe and G.S. Dveksler, 1993. General concepts for PCR primer design. Genome Res., 3: S30-S37.
Direct Link - Elnifro, E.M., A.M. Ashshi, R.J. Cooper and P.E. Klapper, 2000. Multiplex PCR: Optimization and application in diagnostic virology. Clin. Microbiol. Rev., 13: 559-570.
CrossRefDirect Link - Farag, N.S., A.A. Gomah and N.M.A. Balabel, 2010. False negative multiplex PCR results with certain groups of antibiotics. Plant Pathol. J., 9: 73-78.
CrossRefDirect Link - Brownie, J., S. Shawcross, J. Theaker, D. Whitcombe, R. Ferrie, C. Newton and S. Little, 1997. The elimination of primer-dimer accumulation in PCR. Nucl. Acids Res., 25: 3235-3241.
Direct Link - Kabeil, S.S., S.M. Lashin, M.H. El-Masry, M.A. El-Saadani, M.M. Abd-Elgawad and A.M. Aboul-Einean, 2008. Potato brown rot disease in Egypt: Current status and prospects. Am. Eurasian J. Agric. Environ. Sci., 4: 44-54.
Direct Link - Kelman, A., 1954. The relationship of pathogenicity of Pseudomonas solanacearum to colony appearance in a tetrazolium medium. Phytopathology, 44: 693-695.
Direct Link - Kim, H., J. Kim and C. Ihm, 2010. The usefulness of multiplex PCR for the identification of bacteria in joint infection. J. Clin. Lab. Anal., 24: 175-181.
CrossRefPubMedDirect Link - Martin, R.R., D. James and C.A. Levesque, 2000. Impacts of molecular diagnostic technologies on plant disease management. Ann. Rev. Phytopathol., 38: 207-239.
CrossRefDirect Link - Mutter, G.L. and K.A. Boynton, 1995. PCR bias in amplification of androgen receptor alleles, a trinucleotide repeat marker used in clonality studies. Nucleic Acids Res., 23: 1411-1418.
PubMed - Powelson, M.L. and D.A. Inglis, 1999. Foliar fungicides as protective seed piece treatments for management of late blight of potatoes. Plant Dis., 83: 265-268.
Direct Link - Polz, M.F. and C.M. Cavanaugh, 1998. Bias in template-to-product ratios in multitemplate PCR. Applied Environ. Microbiol., 64: 3724-3730.
PubMedDirect Link - Rossen, L., P. Norskov, K. Holmstrom and O.F. Rasmussen, 1992. Inhibition of PCR by components of food samples, microbial diagnostic assays and DNA-extraction solutions. Int. J. Food Microbiol., 17: 37-45.
CrossRefPubMedDirect Link - Samoucha, Y. and Y. Cohen, 1990. Toxicity of propamocarb to the late blight fungus on potato. Phytoparasitica, 18: 27-40.
CrossRefDirect Link - Seal, S.E., L.A. Jackson, J.P.W. Young and M.J. Daniels, 1993. Differentiation of Pseudomonas solanacearum, Pseudomonas syzygii, Pseudomonas pickettii and the blood disease bacterium by partial 16S rRNA sequencing: Construction of oligonucleotide primers for sensitive detection by polymerase chain reaction. J. General Microbiol., 139: 1587-1594.
PubMed - Tsai, Y.L. and B.H. Olson, 1992. Rapid method for separation of bacterial DNA from humic substances in sediments for polymerase chain reaction. Applied Environ. Microbiol., 58: 2292-2295.
Direct Link - Walsh, P.S., H.A. Erlich and R. Higuchi, 1992. Preferential PCR amplification of alleles: mechanisms and solutions. PCR Methods Appl., 1: 241-250.
PubMed - Wagner, A., N. Blackstone, P. Cartwright, M. Dick and B. Misof et al., 1994. Surveys of gene families using polymerase chain reaction: PCR selection and PCR drift. Syst. Biol., 43: 250-261.
Direct Link - Watson, R.J. and B. Blackwell, 2000. Purification and characterization of a common soil component which inhibits the polymerase chain reaction. Can. J. Microbiol., 46: 633-642.
PubMedDirect Link - Wilson, I.G., 1997. Inhibition and facilitation of nucleic acid amplification. Applied Environ. Microbiol., 63: 3741-3751.
PubMedDirect Link