
K.K. Sabet
Department of Plant Pathology, Faculty of Agricultural, Cairo University, Giza, Egypt
M.S. Mansour
Department of Plant Pathology, Faculty of Agricultural, Cairo University, Giza, Egypt
S.A. El-Hadad
Plant Pathology Research Institute, A.R.C., Giza, Egypt
Abeer M. Shaltout
Plant Pathology Research Institute, A.R.C., Giza, Egypt
Sozan E. El-Abeid
Plant Pathology Research Institute, A.R.C., Giza, Egypt
Asian Journal of Plant Pathology
Year: 2013 | Volume: 7 | Issue: 2 | Page No.: 60-73







ABSTRACT
The present study was to investigate the variation among Glomus species. Standard method used for Extraction, purification of Vesicular Arbuscular Mycorrhizal (VAM) fungi, during summer and winter season. Study Effect of some factors on number of spores collected from the rhizosphere of various plants in Egypt. Used single spores technique inoculated on maize root to purified and identified isolates of Glomus spp. The highest number of spores was found in summer season while the lowest number was found in winter season. Variation of Glomus spp. isolates among polymorphic DNA markers was studied. Species 2, 3, 4 and 7 identified by VANS1/VALETC primer as Glomus etunicatum. Fatty Acid Methyl Ester (FAME) profiles were analyzed to assess the diversity and quantity of fatty acids in 8 isolates of Glomus species. Spores and endomycorrhizal roots of Sudan grass (Sorghum sudanense) were examined. Analysis of lipid for species found that affinity specific and generic levels. Differentiation on the genus level using lipid profiles and content which was carried out by evaluating the methyl esters of fatty acids, revealed convergence between studied isolates. Palmitic acid (16:0), Oleic acid (cis18:1n9) and Stearic acid (18:0) were the dominant fatty acid of the Glomus spp., while the dominant fatty acids of Glomus etunicatum isolates were myristic acid (14:0) palmitoleic acid (16:1n7 cis), archidic acid (20:0), palmitic acid (16:0), oleic acid (cis18:1n9) and stearic acid (18:0). This study indicates that using lipids as biochemical markers to identify the different genus of AM fungi and differentiate between the species.
PDF Abstract XML References Citation
Received: January 11, 2013;
Accepted: April 04, 2013;
Published: June 29, 2013
How to cite this article
K.K. Sabet, M.S. Mansour, S.A. El-Hadad, Abeer M. Shaltout and Sozan E. El-Abeid, 2013. Differentiation Between Glomus Species in Egyptian Soil Using Fatty
Acid Methyl Ester Profiles. Asian Journal of Plant Pathology, 7: 60-73.
URL: https://scialert.net/abstract/?doi=ajppaj.2013.60.73
URL: https://scialert.net/abstract/?doi=ajppaj.2013.60.73
INTRODUCTION
Soil fungi are not always infectious to plants or causing deleterious effects for them but some fungi could be very beneficial for plants. The majority of plants species on earth can form a symbiotic relationship with some kind of fungi, known as mycorrhizae, this symbiotic relationship is a beneficial association for both of them. In such a relationship mycorrhizae are able to capture and deliver elements, mainly phosphorus and zinc, to the plants and in turn plants supply mycorrhizae with readymade food essential for them. Therefore, plants living in association with mycorrhizae can show vigorous growth which is due to the easy access to the elements from soil (Sanders et al., 1977; Smith and Read, 1997). Hence, the more mycorrhizae hyphae spreading into the soil and attached to the plant roots, the more efficient the association would be and the plant will be amended extensively with phosphorus.
As Eukaryotes, fungi possess phospholipids in their membranes which are considered a core component of the membranes. In addition, lipids are present in eukaryotic cells as source of energy. However, they mostly differ in the types of fatty acids forming the lipid components in the cells which could be used as biochemical markers to be used for identification. Therefore, phospholipid fatty acids are used to identify the different genus of AM fungi and, in some cases, indentifying to the species level. In addition, they also indicate the fungal biomass in association with plant roots which is also an indication for the efficiency of the AM relations with the plants (Tunlid and White, 1992; Sakamoto et al., 2004). Together with the morphological characters, researchers are using the biochemical and molecular features to distinguish between different mychorhizae species. However, the tendency to use lipids as biochemical markers in identification of AM fungi is due accuracy and to avoid the obstacles of handling and staining the AM hayphae and spores associated with plant roots in order to distinguish morphological characters (Walker and Trappe, 1993).
In previous investigations some phospholipid fatty acids (PLFA) were found to be distinguishing for some AM genus. For instance, Glomus species were found to possess 18:w7, 18:1w9, 20:3, 20:4 and 20:5 in the spores and hyphae associated with plant roots. However, some of these fatty acids were common between Glomus spp and other fungi such as Acaulospora laevis (Beiby, 1980, Beilby and Kidby, 1980; Graham et al., 1995; Jansa et al., 1999; Sakamoto et al., 2004).
On the other hand, some phospholipid fatty acids could be used as markers for AM fungi to the species level. It was detected that PLFA 20:1w9 was used as a marker to detect the presence of Gigaspora rosea (Sakamoto et al., 2004).
Class Glomermycota is a major mycorrhiae fungi which consists of genus commonly found in the Egyptian soil (Fares, 1986).
This study aimed to investigate the survey of vesicular arbuscular mycorrhizal fungi in Egypt, study some factors affecting AM fungi sporulation and to differentiate between the isolated Glomus species using lipids as biochemical markers.
MATERIALS AND METHODS
Extraction, purification and preservation of vesicular arbuscular mycorrhizal (VAM) fungi: Vesicular arbuscular mycorrhizal fungi were collected from the rhizosphere of various plants, through summer and winter months grown in four governorates namely; Beni Suef, Ismailia, El-Menofia and El-Behera. Samples of rhizosphere soil (100 g) was carefully collected from roots of healthy plants and placed in polyethylene bag. Extraction of VAM propagules from soil was carried out under laboratory conditions as soon as possible using wet-sieving and decanting technique described by Gerdemann and Nicolson (1963), Gerdemann and Trappe (1974) and Trappe (1982). Sclerotium rolfsii isolates were obtained from Mycology Research and Diseases Survey Dept., Plant Pathology Research Institute, A.R.C.
Mycorrhizal fungi spores will breed as many as single spores inoculated on maize root, where VAM propagation were multiplied and maintained in open pot cultures of Sudan grass as a host on in vivo medium.
Effect of some factors on number of spores collected from the rhizosphere of various plants-Measuring some factors i.e. electrical conductivity (EC), total dissolved solids (TDS) and pH of soil: Soil-water equilibrium was carried out using the procedure of Negrin et al. (1996) as follows: 10 g of air dried soil (<2 mm fraction) was shaken in 25 mL distilled water on reciprocal shaker for 30 min before equilibration at 4°C were used to prevent biological activity (Hedley and Stewart, 1982) and to favor sedimentation of colloidal material (Schoenau and Huang, 1991). When the equilibration time was attained, the suspensions were centrifuged for 15 min at 2000 Xg and then filtered through a 0.45 Mm Millipore (Millipore corp., Milford, MA) cellulose ester filter. Electrical Conductivity (EC), Total Dissolved Solids (TDS) and pH of the filtrate were measured.
Variation of Glomus spp. isolates among polymorphic DNA markers
DNA isolation technique: DNA of spores of each isolate (approximately 200 spores/isolate) of Eight Glomus spp. previously isolated from different governorates ground to fine powder using a pestle and mortar in liquid nitrogen then placed in a lable 1.5 mL tube then mixed with 600 μL preheat extraction buffer (50 mM EDTA, 100 mM Tris pH 8, 500 mM NaCl) to 65°C in a water bath. 45 μL 10% SDS and 7 μL β merecapto ethanol were adding into each tube and mix them thoroughly by inversion. The samples were incubated at 65°C for 30-40 minutes and mix every 15 min. the samples were cooled down to room temperature before adding 220 μL 5 M potassium acetate into each tube mix thoroughly by inversion then incubate sample on ice or in a freezer for 15 min. the plate were centrifuged for 10 minutes at 5700 rpm. The supernatant was Transferred into a labeled new tubes containing 300 μL of cold isopropanol incubate in -20°C for 24 h, the tube was inverted gently 10-15 times to precipitate DNA for 5 min.
The samples were centrifuged for 10 minutes at 5700 rpm in order to pellet the DNA. The supernatant was discarded by quickly and smoothly inverting the tubes. plate was air dried for 5 min, washed in the two volumes of 70% ethanol (room temperature) were added. The DNA was then pelleted by centrifugation for 12 min (room temperature, 15 000 g). The brown-colored pellet containing the DNA was resuspended in 30 mL TE (10 mM Tris pH 8, 1 mM EDTA). (Dellaporta et al., 1983).
Amplification protocol: The following oligonucleotides used as primers review for the three primers used in this study Simon et al. (1993) was obtained commercially. Their location relative to the SSU gene the sequences of used primers were as shown:
| • | VANS1, 5'GTCTAGTATAATCGTTATACAGG |
| • | NS21, 5'AATATACGOCTATTGGAGCTGG |
| • | VALETC, 5'ATCACCAAGGTTTAGTTGGTTGC |
| • | VAGLO, 5'CAAGGGAATCGGTTGCCCGAT |
The amplification reaction include approximately 50 ng of fungal genomic DNA as template, 1 unit of Taq DNA polymerase, o.2 mM each dNTPase, 1x PCR buffer, 3 mM MgCl2 and 10 pmol of primer. The temperature profile was programmed on a DNA Thermal Cycler (Perkin-Elmer) to repeat 25 to 35 times a cycle of denaturation (60 sec at 94°C), annealing (45 sec at 50°C) and polymerization (60 sec at 72°C), followed by a final extension step of 10 min at 72°C. Hold at 4°C 7 μL of 6 X tracking buffer (manufactured by Qiagen Kit) were added to 25 μL of the amplification product.
The amplified product were separated on 1.5% agarose gel to check for product size and purity, using 1X TBE buffer followed by staining in ethidium bromide solution (1 ug mL-1), at 75 constant volt and determine with UV transilluminator.
The total volume was completed to 25 μL using sterile distilled water. The amplification protocol was carried out as follows using PCR unit II biometra.
Gel analysis: DNA gels were scanned for band RF using gel documentation system (AAB Advanced American Biotechnology 1166 E. Valencia Dr. Unit 6 C, Fullerton CA 92631). The different M W of bands was determined against PCR marker promega G 317A by unweighted pair-group method based on arithmetic mean (UPGMA).
Lipid differentiation by evaluation lipid on root of grass: The root samples (75 mg) were dissolved in toluene (1 mL) in a test tube fitted with a condenser, then H2SO4 in methanol (2 mL, 1%) was added. The mixtures were left overnight in a stoppered tube at 50°C then sodium chloride solution (5 mL, 5%) was added and the required esters were extracted with hexane (2x, 5 mL), then the organic layer was separated using Pasteur pipettes for both samples. The hexane layers were washed with potassium bicarbonate solution (4 mL, 2%) and dried over anhydrous Na2SO4 and filtered. The organic solvent was removed under reduced pressure on a rotary evaporator to give FAMEs and other lipids. GC-MS Conditions: GC-MS analysis was performed in a Fisons MD800 mass (quadrupole) -GC8000 series instrument equipped with a flame ionization detector. A silica column (30 mx0.25 mm I.D.) coated with OV1 was utilized. The initial temperature was 40°C for 4 min after injection and then increased to 280°C (8°C min-1) with a final hold at 280 °C for 20 min. The injector and detector temperature were maintained at 270 and 250°C, respectively. Helium was used as the carrier gas at a flow-rate of 0.8 mL min-1 (Yayli et al., 2001).
Statistical analysis: Data were statistically analyzed according to the standard procedure in completely random design as mentioned by Snedecor and Cochran (1982).
RESULTS
Collection, Isolation and Purification of Vesicular Arbuscular Mycorrhizal (VAM) fungi Extraction of VAM spores from some Egyptian governorates during summer season: Data in Table 1 show that the highest numbers i.e., 107, 101, 81 and 60 of VAM fungal spores g-1 soil isolated from maize sown after winter crops i.e., wheat and clover were found at El-Behira, Ismailia, Beni-Suef and El-Menofia governorates respectively.
| Table 1: | Number of VAM fungi spores extracted from some Egyptian governorates during summer season |
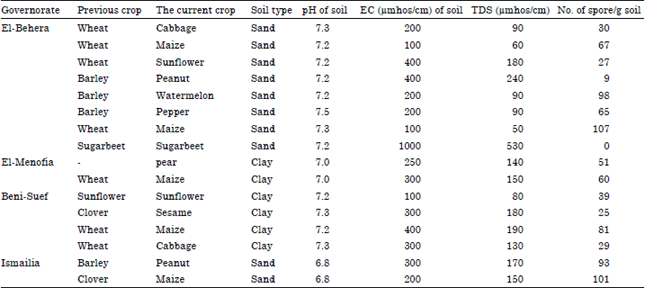 | |
On the other hand, three crops sown after barley i.e. water melon (El-Behira), peanut (Ismailia) and pepper (El-Behira) recorded high rates (98,93 and 65 of VAM spores g-1 soil, respectively).
Type of soil had an effect on survival of VAM fungal spores. It is noticed in Table 1 that sand soil was the best medium for survival of VAM spores. On the other hand pH of Egyptian soil had no effect on the existence of VAM fungal spores. It’s worth to note that there is no VAM fungal spores recorded at EC 1000 μmhos/cm and 530 μmhos/cm of Total Dissolved Solids concentration (TDS) of Egyptian soil.
Extraction of VAM spores from some Egyptian governorates during winter season: Data in Table 2 show that the highest numbers i.e., 45, 26 and 17 VAM fungal spores g-1 soil isolated from onion, lentil and sugar beet respectively sown after maize at Beni-Suef governorate .The average of VAM spores isolated from soil sown with wheat after maize was 23 spores g-1 soil at Elbehera governorate. At El-Menofia, three crops were sown after maize i.e. wheat (20 spores g-1 soil), onion (18 spores g-1 soil) and bean (15 spores g-1 soil); whereas two crops were sown at Ismailia after maize i.e., Barley (16 spore/g soil) and clover (16 spores g-1 soil). Some crops couraged VAM spores existence in the next crops i.e. pear before clover (28 spores g-1 soil) at El-Menofia, soybean before pepper (17 spore/g soil).
It is noticed in that the type of soil had an effect on survival of VAM spores.
Clay soil revealed a good medium for survival of VAM spores.
On the other hand pH of Egyptian soil had no effect on the VAM spores its worth to note that there is no effect at EC 100-400 μmhos/cm and (40-260) TDS of Egyptian soil.
| Table 2: | Number of VAM fungi spores extracted from some Egyptian governorates during winter season |
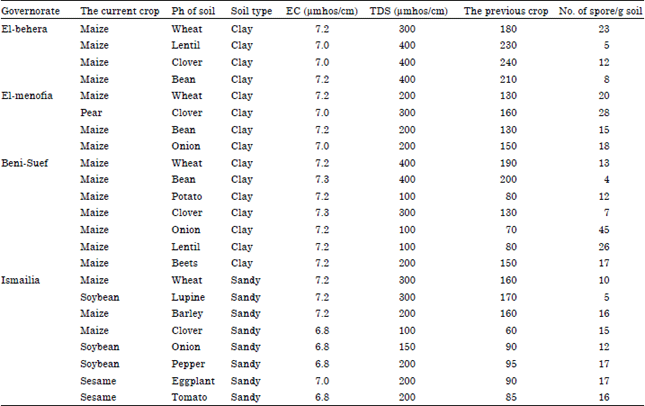 | |
Isolation of VAM spores
Percentage of genera of mycorrhizal fungi in different type soil mixtures: Natural occurrence of VAM fungal species, belonging to four genera such as; Acaulospora, Glomus, Gigaspora and Sclerocystis were observed in the rhizosphere soils of four governorates of Egypt (Behira, El-Menofia, Beni Suef and Ismailia. Frequency percentages of aforementioned genera in 10 g soil were Glomus (57-100%) in aforementioned governorates, Acaulospora (14-22%) in Behira and Ismailia, Gigaspora (21-26%) in Behira and Ismailia and Sclerocystis (32%) in El-menofia. Glomus was the dominant genus in all inspected governments with the highest frequencies as it shown in Table 3.
Purification and Identification of Glomus spp.: Examined spores were identified according to the morphological characteristics using keys of Gerdemann and Trappe (1974), Trappe (1982). Data in Table 4 show the main characteristics (color, shape and size) to determine the differentiation between species of genus Glomus the shape was not critical character where all species had globose and subglobose shape but some spores of species G. mosseae and G. coronatum had irregular shape. It is worth to note that the identification was also made according to the size of Glomus spp. G. intraradicesis was the smallest (50-67 μm) compared with other Glomus spp. While G. mosseae and G. etunicatum 3 was large (<100 μm) but they were different in color (G. mosseae Hyaline-dark orange to brown and G. etunicatum 3 was Pale orange brown to dark orange brown).
Variation of Glomus spp. isolates among polymorphic DNA markers: This analysis was carried out on 8 isolates. The eight isolates were isolated from different Egyptian governorates.
| Table 3: | Frequency Percentages of genera of mycorrhizal fungi in 100 g soil of four governorate of Egypt |
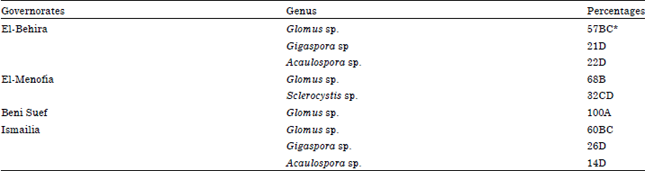 | |
| *Data in the same column followed by the same letter are not significantly different (p<0.05) | |
| Table 4: | Characteristics of Glomus spp., isolated from four governorates in Egypt |
 | |
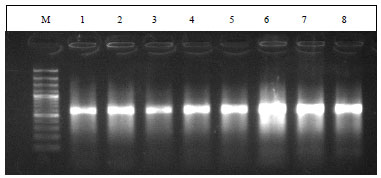 | |
| Fig. 1: | (1%) gel electrophoresis of DNA extracted from 8 strains of VAM fungi to detect the presence of 550-bp ribosomal DNA fragments with VANS1/NS21. M DNA marker, Lanes (1-8) presenting isolates of Glomus G. intraradices, G. etunicatum 1, G. etunicatum 2, G. etunicatum 3, G. mosseae, G. diaphanum, G. etunicatum 4 and G. coronatum respectively, show the presence of 550-bp in all lanes |
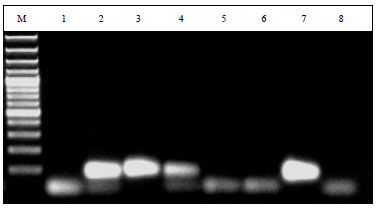 | |
| Fig. 2: | (1%) gel electrophoresis of DNA extracted from 8 strains of VAM fungi to detect the presence of 100-bp ribosomal DNA fragments with VANS1/VALETC primer. M DNA marker, Lanes (1-8) presenting isolates of Glomus G. intraradices, G. etunicatum1, G. etunicatum 2, G. etunicatum 3, G. mosseae, G. diaphanum, G. etunicatum 4 and G. coronatum respectively, show the presence of 100-bp in lanes 2, 3, 4, 7 |
The VANS1/NS21 primer pair was used to amplify 550-bp ribosomal (r) DNA fragment from vesicular arbuscular mycorrhizal fungi, directly from spores and colonized root extracts. A procedure to detect these obligatory biotrophs rapidly, based on competitive PCR, was developed by constructing a suitable internal standard to be used with these primers. DNA fragment appeared identical in all studded Glomus spp. (Fig. 1).
The VANS1/VALETC primer pair was used to amplifying a 100bp ribosomal (r) DNA fragment would be classified as belonging to Glomus etunicatum, directly from spores and colonized root extracts. A procedure to detect these obligatory biotrophs rapidly, based on competitive PCR, was developed by constructing a suitable internal standard to be used with these primers. DNA fragment appeared identical in isolates 2,3,4,7 Glomus etunicatum as shown in Fig. 2.
The lipid differentiation by evaluation lipid on root of grass: Data in Table 5 the total Methyl Esters Fatty Acids in all 8 Glomus isolates on root of grass which illustrated that Palmitic acid (16:0), Oleic acid (cis18:1n9) and Stearic acid (18:0) were the dominant fatty acid of the Glomus spp. Saturated fatty acid, 16:0 was present in all isolates and constituted 16.2 to 41.5% of the total profile.
| Table 5: | Total methyl esters fatty acids in glomus isolates on root of grass |
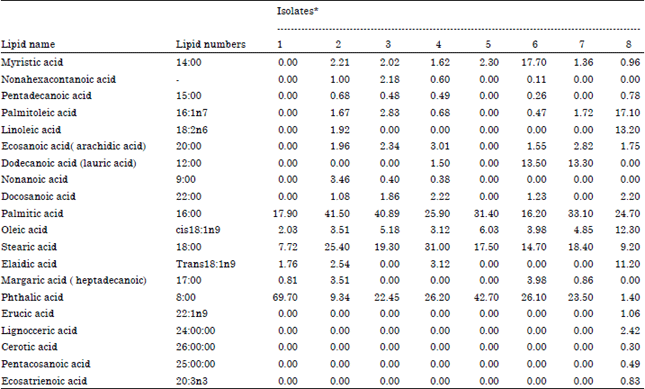 | |
One unsaturated fatty acid, cis18:1n9 was present in all isolates and constituted 2.03-12.3% of the total profile. A high proportion of 18:1n9 cis also occurred in (isolate 8) G. coronatum was unique in possessing most of fatty acid. Conversely, 18:1n9 Trans accounted for 1.76 to 11.2% of the total fatty acids in Glomus isolates 1,2,4,8.
The Myristic acids (14:0), ranging from 0.96 to 17.7%, palmitoleic acid (16:1n7 cis) 0.47 to 17.1, archidic acid (20:0) 1.75 to 3.01, palmitic acid (16:0 )16.2 to 41.5, oleic acid (cis18:1n9) 2.03 to 12.3 and stearic acid (18:0) 7.72 to 25.4 were present in all isolates of Glomus etunicatum isolates 2,3,4,7, also Linoleic acid (18:2n6) was present only in Glomus etunicatum isolates (2) and G. coronatum (isolate 8).
DISCUSSION
In summer season it was found that the highest numbers of vesicular arbuscular mycorrhizal (VAM) fung l spores g-1 soil were recorded on maize grown after winter crops i.e. wheat and clover at El-Behira, Ismailia, Beni-Suef and El-Menofia governorates respectively. On the other hand, three crops grown after barley, i.e., water melon (El-Behira), peanut (Ismailia) and pepper (El-Behira) recorded high numbers of spores. In winter season the highest VAM fungi spores numbers were extracted from onion, lentil and sugar beet rhizosphere respectively grown after maize at Beni-Suef governorate. Sand soil was the best medium for survival of VAM spores in summer season but Clay soil revealed a good medium for survival of VAM spores in winter season. On the other hand pH of Egyptian soil had no effect on the VAM spores. Fares (1986) examined soil samples taken from around the roots of 23 different hosts grown in 5 sites in Egypt, i.e., El- Kaliobia, Ismailia, Alexandria, El-Minia and Sinai desert.
Schultz et al. (1999) and Bever et al. (2001) also found evidence that AM fungi differ in their seasonality, with some fungi sporulating in late spring and others sporulating at the end of summer. As the spores represent the dormant state of the fungus, the physiologically active state is most likely the mirror image of the seasonal spore counts. Therefore, Gigaspora gigantea which sporulates most abundantly in the fall and appears to overwinter as spores is likely to be physiologically active during the warm season. Similar patterns have been seen for Gi. gigantea in a sand dune on the coast of Rhode Island (Gemma et al., 1989; Lee and Koske 1994). On the other hand, Acaulospora colossica which sporulates most profusely at the beginning of summer and over summers as spores, is physiologically active with the cool season plant community (e.g., Allium vineale). Hayman (1982) indicated that variation in the populations of the fungi and their symbiosis with plant roots is related to both soil properties and host plants. In addition, species and isolates of AMF differ in their tolerance to adverse physical and chemical conditions in soil (Juniper and Abbott 1993).
Arbuscular mycorrhizal fungi are also known to vary in their response to the mineral environment of the soil. About 30% of the variation in the spatial distribution of AM fungi could be explained by variation in aspects of the mineral soil (Schultz 1996; Bever et al., 2001). Individual fungi showed opposite associations with certain soil parameters. For example, the distribution and abundance of A. colossica was negatively associated with soil phosphorus concentration, while the reverse was true for G. gigantea (Schultz, 1996). This dependence of fungal spatial distributions on edaphic factors is consistent with observations in other communities, including tall grass prairie (Johnson et al. 1992) and sand dunes (Koske 1981). However, previous study on AM fungi sporulation in different seasons showed that heat can reduce AM fungal spore production in arid and semiarid areas (Camargo-Ricalde and Esperon-Rodriguez 2005). Lugo et al., 2005 found plant-specific and AM fungi-specific variations in the formation of intraradical AM fungal structures and spores in the moist and dry part of the year in an Argentinean dry climate with no seasonal variation in temperature.
In this study natural occurrence of VAM fung, belonged to four genera such as; Acaulospora, Glomus, Gigaspora and Sclerocystis were observed in the rhizosphere soils of four governorates in Egypt Behira, El-Menofia, Beni Suef and Ismailia . Percentages of aforementioned genera in 10 g soil were Glomus (57-100%) in all governorates, Acaulospora (14-22%) in Behira and Ismailia, Gigaspora (21-26%) in Behira and Ismailia and Sclerocystis (32%) in El-menofia. Glomus was the dominant genus in all inspected governments with the highest frequencies. This result agrees with that of Fares (1986) showed that morphological characteristics revealed that the tested spores belonged to the genera Glomus, Gigaspora and Sclerocystis. Spores belonged to genus Glomus were the most dominant. These spores were identified as G. mosseae, G. clarum and G. fasciculatum. A good number of spores was recorded in the rhizosphere soil of non-colonized plant species in saline depression habitat. Tressner and Hayes (1971), Hirrel (1981) suggested that AMF sporulation is stimulated under salt stress conditions, but In this study arbuscular mycorrhizal fungi were found to be present in almost all samples and show that AMF and their symbiosis with plants are widely spread in Egypt.
Simon et al. (1993) designed primers to identify AMF in colonized roots by PCR fragment amplification of the 18S rDNA combined with the SSCP analysis. The VALETC, VAGLO, VAACAU and VAGIGA primers were designed to discriminate among four distinct groups of endomycorrhizal species. Furthermore they designed the VANS22 and VANS32 which were able to amplify a 150-bp informative fragment from any endomycorrhizal fungi. These primers were not designed specifically for Glomales and Simon et al. (1993) mentioned the possibility that they could also be useful for other fungi or eukaryotes. Later, Simon and Lalonde (1995) designed and patented the VANS1 primer which amplified part of the 18S rDNA from AMF (Glomus intraradices and Gigaspora margarita) directly from colonized roots. However, Simon (1996) concluded that primers pairs VANS22/VANS32 and NS71/SSU1492 can only detect AMF genus differences. Furthermore, he reported that those primers are not AMF specific and samples must be treated with a nested PCR amplified firstly by the primers VANS1 (Glomalean specific) and VANS22 and then the amplicons amplified by the VANS22/VANS32 primers.
The results of the present study show that VANS1/NS21 primer pair is useful for specifically amplifying use as review for the three primers used in this study Simon et al. (1993) 550-bp ribosomal (r) DNA fragment from arbuscular endomycorrhizal fungi, directly from spores and colonized root extracts. A procedure to these obligatory biotrophs rapidly, based on competitive PCR, was developed by constructing a suitable internal standard to be used with these primers. The taxon-specific primers, VANS1 and NS21. DNA fragment showed identical in all arbuscular endomycorrhizal fungi. These results are similar to those obtained by Simon et al (1992) who indicated that a portion of the small subunit rRNA gene (rDNA) specific for AM fungi can be amplified when a taxon-specific primer (VANS1) is paired with a universal primer (NS21). And examined the specificity of the VANS1 primer, when paired it with a downstream universal primer (NS21) and tested its ability to amplify a 550-bp portion of the SSU sequences from DNAs of other arbuscular endomycorrhizal fungi as well as a variety of other fungi and plants. The fungi selected for testing spanned a broad phylogenetic range and included putative relatives of arbuscular endomycorrhizal fungi as well as other rhizosphere associates such as ectomycorrhizal species and root pathogens. Also Di Bonito et al. (1995) concluded that PCR was used with the primer pair VANS1-NS21 to detect the arbuscular mycorrhizal fungus Glomus intraradices (commercial inoculum source) on roots of lettuce, zinnia, leek, pepper and endive plants and during the application of these methods a number of problems have been identified. Specificity is limited to the level of genus and may not be absolute. The studies of PCR application for AM fungi have been carried out with spores or purified DNA from colonized leek roots (Wyss and Bonfante, 1993) the AM taxon-specific primer VANS1 was used for detection of the AM fungus Glomus intraradices on roots of five host species colonized by this fungus. This was accomplished without extraction of DNA from colonized roots by simply boiling root tissue in a buffer (Henson et al., 1993).
The VANS1/VALETC primer pair is useful for specifically amplifying use as review for the three primers used in this study Simon et al. (1993) 100bp ribosomal (r) DNA fragment would be classified as belonging to Glomus etunicatum, directly from spores and colonized root extracts. A procedure to these obligatory biotrophs rapidly, based on competitive PCR, was developed by constructing a suitable internal standard to be used with these primers. The taxon-specific primers, VANS1 and VALETC. DNA fragment showed identical in isolates 2, 3, 4, 7 Glomus etunicatum. SSCP analysis is a technique that could detect sequence variation, sometimes as small as a single-base substitution, between DNA fragments ranging from 100 to 450 bp Simon et al. (1992). By using SSCP analysis, fragments amplified using a single primer pair could be further characterized without the need for other specific primers or probes. This was illustrated by the analysis of fragments amplified with primer pair VANS1-VAACAU, with which discrimination between related species was achieved.
In this study Palmitic acid (16:0), Oleic acid (cis18:1w9) and Stearic acid (18:0) were the dominant fatty acid of the Glomus spp. Saturated fatty acid, 16:0 was present in all isolates and constituted 2.65 to 20% of the total profile. As Graham et al. (1995) reported that in spores, 16:0 was present in all isolates of Glomus spp. and constituted 10 to 38% of the total profile. Also some fatty acids have been known as specific markers for AM fungi. In the case of Glomus species, fatty acids 16:0, 16:1n5, 18:1n7, 18:1n9, 20:3, 20:4 and 20:5 have been detected in Glomus spores and the roots of plants colonized by Glomus spp. (Beilby and Kidby, 1980; Jabaji-Hare, 1988; Pacovsky and Fuller, 1988; Pacovsky, 1989; Graham et al., 1995; Jansa et al., 1999; Madan et al., 2002). On the other hand (Sakamoto et al., 2004) conclude that 18:1n9 and 20:1n9 can be used as a specific marker for identifying and quantifying the external hyphae of Gigaspora species in soil (Bentivenga and Morton, 1994, 1996; Graham et al., 1995; Madan et al., 2002).
One unsaturated fatty acid, cis18:1n9 was present in all isolates and constituted 0.30 to 6.84% of the total profile. A high proportion of 18:1n9 cis also occurred in (isolate 8) G. coronatum was unique in possessing most of fatty acid. Conversely, 18:1n9 Trans accounted for 0.26 to 6.18% of the total fatty acids in Glomus isolates 1,2,4,8. As Graham et al. (1995) reported that 18:1n9 cis accounted for 38 to 48% of the total fatty acids in all Gigaspora isolates tested. A high proportion of 18:1n9 cis also occurred in G. leptotichum and Scutellospora persica. Isolates of G. leptotichum and G. occultum were unique in possessing 16:1n7 cis. These results are in a harmony with those reported by Peng et al. (1993) where he stated that 16:1n7 cis was erroneously reported to be the principal fatty acid present in citrus roots infected by G. intraradices . Many authors have reported the presence of 18:1n9 in the spore of Glomus,Gigaspora, Scutellospora and Acaulospora species (Graham et al., 1995; Bentivenga and Morton, 1996; Madan et al., 2002) and in the soybean root colonized by Glomus fasciculatum (Pacovsky and Fuller, 1988).
The Myristic acids (14:0), ranging from 0.4 to 1.06%, palmitoleic acid (16:1n7 cis) 0.18 to 1.05, archidic acid (20:0) 0.8 to 1.08, palmitic acid (16:0 )6.9 to 12.65, oleic acid (cis18:1n9) 0.83 to 1.68 and stearic acid (18:0) 7.06 to 12.15 were present in all isolates of Glomus etunicatum isolates 2, 3, 4, 7 also Linoleic acid (18:2n6) was present only in Glomus etunicatum isolates (2, 3, 4, 7) and G. coronatum (isolate 8).
Morton (1985) stated that Profiles of G. etunicatum were distinct in that 20:0 iso was unique to these isolates and the one isolate of G. claroideum studied. G. leptotichum and G. occultum isolates were distinguished from all other glomalean fungi by the high levels of 16:1n7 cis (almost 50% in two G. occultum isolates).
The end product of fungal Fatty Acid Synthesis (FAS) is palmitic acid (16:0) which can then be elongated and desaturated to produce all of the other fatty acids (Williams et al., 1987). Trepanier et al. (2005) reported that in germinating spores and the extraradical hyphae of G. intraradices supplied with [14C] acetate, only fatty acids longer than 16 carbons were labeled which suggests that no de novo fatty acid synthesis by FAS was occurring in these two fungal cell domains. Previous work (Phillips and Hayman, 1970) also found no labeling of 16:1n5 when extraradical hyphae were supplied with 13C-acetate. In other study, germinating spores and the extraradical mycelium of G. intraradices could elongate pre-existing 16-carbon fatty acids (16:0 or 16:1) with labeled acetate to form 18- and 20-carbon fatty acids. An equivalent result was obtained with G. rosea, demonstrating that this fungus also lacks FAS activity in germinating spores. These two phylogenetically distant AM fungi share the same lack of enzymatic activity suggests that the lack of FAS syntheses might be a general feature of the early development of AM fungi. 16-Carbon fatty acids of G. intraradices were labeled only when [14C] acetate or [14C] sucrose was added to colonized roots. In the root compartment, plant fatty acids were highly labeled and mixed with the fungal fatty acids. Carrot roots contain only minute levels of 16:1, so this strongly labeled fatty acid was almost exclusively of fungal origin. Both organisms synthesize the other fatty acids and their origins cannot be distinguished. When radioactive precursors were included in the root compartment, all fungal compartment fatty acids also became highly labeled, especially the most abundant one: 16:1.
CONCLUSION
The previous study showed that there were many factors affecting the activity of mycorrhizal fungi such as season. Palmitic acid (16:0), Oleic acid (cis18: 1w9) and Stearic acid (18:0) were found in all isolates that were under investigation. However, other fatty acid was found to be specific for the species.
REFERENCES
- Beilby, J.P., 1980. Fatty acid and sterol composition of ungerminated spores of the vesicular-arbuscular mycorrhizal fungus, Acaulospora laevis. Lipids, 15: 949-952.
CrossRef - Beilby, J.P. and D.K. Kidby, 1980. Biochemistry of ungerminated and germinated spores of the vesicular-arbuscular mycorrhizal fungus, Glomus caledonius: Changes in neutral and polar lipids. J. Lipid Res., 21: 739-750.
Direct Link - Bentivenga, S.P. and J.B. Morton, 1994. Stability and heritability of fatty acid methyl ester profiles of glomalean endomycorrhizal fungi. Mycol. Res., 98: 1419-1426.
CrossRef - Bentivenga, P.S. and J.B. Morton, 1996. Congruence of fatty acid methyl ester profiles and morphological characters of arbuscular mycorrhizal fungi in Gigasporaceae. Proc. Natl. Acad. Sci. USA., 93: 5659-5662.
Direct Link - Bever, J.D., P.A. Schultz, A. Pringle and J.B. Morton, 2001. Arbuscular mycorrhizal fungi: More diverse than meets the eye and the ecological tale of why. BioScience, 51: 923-931.
Direct Link - Camargo-Ricalde, S.L. and M. Esperon-Rodriguez, 2005. Effect of the spatial and seasonal soil heterogeneity over arbuscular mycorrhizal fungal spore abundance in the semi-arid valley of Tehuacan-Cuicatlan, Mexico. Rev. Biol. Trop., 53: 339-352.
PubMedDirect Link - Dellaporta, S.L., J. Wood and J.B. Hicks, 1983. A plant DNA minipreparation: Version II. Plant Mol. Biol. Rep., 1: 19-21.
CrossRefDirect Link - Di Bonito, R., M. Elliot and E. Jarclin, 1995. Detection of an arbuscular mycorrhizal fungus in roots of different plant species with the PCR. Applied Environ. Microbiol., 61: 2809-2810.
Direct Link - Gemma, J.N., R.E. Koske and M. Carreiro, 1989. Seasonal dynamics of selected species of V-A mycorrhizal fungi in a sand dune. Mycol. Res., 92: 317-321.
CrossRef - Graham, J.H., N.C. Hodge and J.B. Morton, 1995. Fatty acid methyl ester profiles for characterization of glomalean fungi and their endomycorrhizae. Applied Environ. Microbiol., 61: 58-64.
Direct Link - Hedley, M.J. and J.W.B. Stewart, 1982. Method to measure microbial phosphate in soils. Soil Biol. Biochem., 14: 377-385.
CrossRef - Hirrel, M.C., 1981. The effect of sodium and chloride salts on the germination of Gigaspora margaria. Mycology, 43: 610-617.
Direct Link - Jabaji-Hare, S., 1988. Lipid and fatty acid profiles of some vesicular-arbuscular mycorrhizal fungi: Contribution to taxonomy. Mycologia, 80: 622-629.
Direct Link - Johnson, N.C., P.J. Copeland, R.K. Crookston and F.L. Pfleger, 1992. Mycorrhizae: Possible explanation for yield decline with continuous corn and soybean. Agron, J., 84: 387-390.
Direct Link - Juniper, S. and L. Abbott, 1993. Vesicular-arbuscular mycorrhizas and soil salinity. Mycorrhiza, 4: 45-57.
CrossRefDirect Link - Koske, R.E., 1981. A preliminary study of interactions between species of vesicular-arbuscular fungi in a sand dune. Trans. Br. Mycol. Soc., 76: 411-416.
CrossRef - Lee, P.J. and R.E. Koske, 1994. Gigaspora gigantea: Seasonal abundance and ageing of spores in a sand dune. Mycol. Res., 98: 453-457.
CrossRef - Lugo, M.A., A.M. Anton and M.N. Cabello, 2005. Arbuscular mycorrhizas in the Larrea divaricata scrubland of the arid Chaco, Central Argentina. J. Agric. Technol., 1: 163-178.
Direct Link - Madan, R., C. Parkhurst, B. Hawke and S. Smith, 2002. Use of fatty acids for identification of AM fungi and estimation of the biomass of AM spores in soil. Soil Biol. Biochem., 34: 125-128.
CrossRef - Morton, J.B., 1985. Variation in mycorrhizal and spore morphology of Glomus occultum and Glomus diaphanum as influenced by plant host and soil environment. Mycologia, 77: 192-200.
Direct Link - Negrin, M.A., M. Espino-Mesa and J.M. Hernandez-Moreno, 1996. Effect of water: Soil ratio on phosphate release: P, aluminium and fulvic acid associations in water extracts from Andisols and Andic soils. Eur. J. Soil Sci., 47: 385-393.
CrossRef - Pacovsky, R.S., 1989. Metabolic differences in Zea-glomus-azospirillum symbioses. Soil Biol. Biochem., 21: 953-960.
CrossRef - Pacovsky, R.S. and G. Fuller, 1988. Mineral and lipid composition of Glycine-glomus-bradyrhizobium symbioses. Physiol. Plant., 72: 733-746.
CrossRef - Phillips, J.M. and D.S. Hayman, 1970. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc., 55: 158-161.
CrossRefDirect Link - Sakamoto, K., T. Iijima and R. Higuchi, 2004. Use of specific phospholipid fatty acids for identifying and quantifying the external hyphae of the arbuscular mycorrhizal fungus Gigaspora rosea. Soil Biol. Biochem., 36: 1827-1834.
CrossRef - Sanders, F.E., P.B. Tinker, R.L.B. Blacfk and S.M. Palmerley, 1977. The development of endomycorrhizal root systems. I. Spread of infection and growth-promoting effects with four species of vescular-arbuscular endophyt. New Phytol., 78: 257-268.
CrossRef - Schoenau, J.J. and W.Z. Huang, 1991. Anion-exchange membrane, water and sodium bicarbonate extractions as soil tests for phosphorus. Commun. Soil Sci. Plant Anal., 22: 465-492.
CrossRef - Schultz, P.A., J.D. Bever and J. Morton, 1999. Acaulospora colossica sp. nov. from an old field in North Carolina and morphological comparisons with similar species, A. laevis and A. koskei. Mycologia, 91: 676-683.
Direct Link - Simon, L., 1996. Phylogeny of the glomales: Deciphering the past to understand the present. New Phytol., 133: 95-101.
CrossRefDirect Link - Simon, L., R.C. Levesque and M. Lalonde, 1993. Identification of endomycorrhizal fungi colonizing roots by fluorescent single-strand conformation polymorphism polymerase chain reaction. Applied Environ. Microbiol., 59: 4211-4215.
Direct Link - Trepanier, M., G. Becard, P. Moutoglis, C. Willemot, S. Gagne, T.J. Avis and J.A. Rioux, 2005. Dependence of arbuscular-mycorrhizal fungi on their plant host for palmitic acid synthesis. Applied Environ. Microbiol., 71: 5341-5347.
CrossRef - Tresner, H.D. and J.A. Hayes, 1971. Sodium chloride tolerance of terrestrial fungi. Applied Environ. Microbiol., 22: 210-213.
Direct Link - Walker, C. and J.M. Trappe, 1993. Name and epithets in the Glomales and endogonales. Mycol. Res., 97: 339-344.
CrossRef - Williams, K., F. Percival, J. Merino and H.A. Mooney, 1987. Estimation of tissue construction cost from heat of combustion and organic nitrogen content. Plant Cell Environ., 10: 725-734.
CrossRef - Wyss, P. and P. Bonfante, 1993. Amplification of genomic DNA of Arbuscular-Mycorrhizal (AM) fungi by PCR using short arbitrary primers. Mycol. Res., 97: 1351-1357.
CrossRef - Yayli, N., Z. Kiran, H. Seymen, H. Genc and M. Kucukislamoglu, 2001. Characterization of lipids and fatty acid methyl ester contents in leaves and roots of Crocus vallicola. Turk. J. Chem., 25: 391-396.
Direct Link