
S.O. Aigbe
Department of Crop Science, Ambrose Alli University, PMB 14 Ekpoma, Nigeria
S.U. Remison
Department of Crop Science, Ambrose Alli University, PMB 14 Ekpoma, Nigeria
Asian Journal of Plant Pathology
Year: 2010 | Volume: 4 | Issue: 2 | Page No.: 82-89







ABSTRACT
A field experiment to determine the influence of root rot on dry matter partition of three cassava cultivars planted in the Sabongidda-Ora humid forests and in the Ibadan derived savanna of Nigeria was carried out from July 2003-November 2005. In each location, healthy cassava stems of three varieties; TME-1 (local), TMS 30572 and TMS 4 (2) 1425 (both improved) were planted in a randomized complete block design with 3 replications. Dry matter partition data were recorded and statistically analysed using the general linear model statistical procedures. Comparisons among treatment means less than six were made with the Least significant square and up to six and above with the Duncan multiple range test (p = 0.05). Results were compared with the previously reported root rot incidences and severities from the same cultivars. Results showed cultivar TME-1; previously reported to have the highest root rot incidence of 52.6% and a severity of 21.3% at 12 months after planting in Sabongidda-Ora humid forest, to have the lowest dry matter partition of 367.4 g to roots in the same location, 12 months after planting. This was significantly lower than the 804.1 g of TMS 4(2)1425 and the 667.6 g of TMS 30572 recorded in the same location; cultivars TMS 4(2)1425 and TMS 30572 have also been reported to have the lowest root rot incidences of 0 and 6.4% and severities of 0-0.5% at the Ibadan derived savanna. The trend was the same during the second year planting. The results show clearly that root rot can have significantly negative effect on the dry matter partition to the storage roots of root rot susceptible cassava cultivar.
PDF Abstract XML References Citation
Received: January 20, 2010;
Accepted: July 09, 2010;
Published: July 27, 2010
How to cite this article
S.O. Aigbe and S.U. Remison, 2010. The Influence of Root Rot on Dry Matter Partition of Three Cassava Cultivars Planted in Different Agro-ecological Environments. Asian Journal of Plant Pathology, 4: 82-89.
URL: https://scialert.net/abstract/?doi=ajppaj.2010.82.89
URL: https://scialert.net/abstract/?doi=ajppaj.2010.82.89
INTRODUCTION
Root yield components of a cassava (Manihot esculenta Crantz.) plant are number of storage roots per plant, average root weight (fresh) and root dry matter content. In cassava, foliage and storage roots develop simultaneously; resulting in competition for assimilates, assimilate supply is thus divided between foliage and storage roots. In cassava, partitioning of dry matter to different parts of the plant varies during the growth cycle. Allocation of dry matter to the storage roots varies from almost zero during the early growth stages to nearly 80% of the daily dry matter production during the late growth stages. The relationship between total dry weight of the plant and dry weight of the storage roots is linear, suggesting that the rate of root growth keeps pace with the rate of crop growth (Ekanayake et al., 1998).
A typical growth pattern of a cassava plant shows dry matter production and vegetative development. A dry matter production result in an increase of assimilates during the life cycle of the crop. Phasic development (Vegetative plant growth) implies a sequence of four growth phases for a crop harvested after 12 months (Ekanayake et al., 1998). The phases are as follows:
| • | Establishment phase commences when stem cuttings sprout, leaves form and vegetative growth begins |
| • | Storage root initiation phase begins with the initiation and formation of storage roots that start to compete for assimilates |
| • | The storage root-bulking phase is the phase when storage roots enlarge while the leaf area reaches a maximum. When the crop is exposed to a dry period at the latter part of this phase, the leaf area reduces and crop growth slows down |
| • | Recovery phase is the stage when cassava re sprouts and produces a new canopy. Storage root growth and competition for assimilates continue |
Cassava tuberous roots are sometimes left in the ground after plant maturity and harvested as required for consumption (Onyeka et al., 2005). Such underground storage and delayed harvest is problematic where the incidence of root rot is high. Cassava root rot can be caused by fungi, bacteria and some parasitic mushrooms (Moses et al., 2008). In the surveys of 16 states of the humid forest and moist savanna agro-ecologies in Nigeria, tuberous root rots were recorded to be widespread constraints of cassava production (IITA, 2000). Earlier reports highlighted the importance of root rots as constraints to cassava production (Lozano, 1989), but more recent reviews ranked root rots behind mosaic disease and bacterial blight infections (Hillocks and Wydra, 2002; Dixon et al., 2003). Yield losses of up to 25% have been documented in Africa (Messiga et al., 2004). Based on surveys of farmers in the humid forest area of Cameroon, 58% of the sampled population (n = 120) ranked root rot disease as the main constraint to cassava production (Mwangi, M.; Bandyopadhyay, R. and Nolte, C. unpublished data). For over fifteen years in Ghana, a parasitic mushroom (Polyporus sulphureus), originally suspected to be pathogenic on woody plants, has been found attacking cassava plants and causing severe root rots. This parasitic mushroom has been reported as capable of causing 100% yield loss on farms where susceptible cultivars are planted (Moses et al., 2008).
Effective and reliable control measures will remain elusive as long as little is known concerning the disease and its host-pathogen relationships. The objective of the investigation in this study therefore was to investigate the influence of root rot on dry matter partition in three cassava cultivars planted in different ecological environments in Nigeria.
MATERIALS AND METHODS
In July 2003 cassava field trial was set up in IITA experimental fields located in Ibadan (derived savanna). This same trial was repeated in August 2003 at Sabongidda-Ora (humid forest). Healthy cassava stems of three varieties, TME-1 (local), TMS 30572 (improved) and TMS 4 (2) 1425 (improved) were planted in a randomized complete block design with 3 replications. Within each plot were 12 rows of cassava stands and each row was a ridge of 15 stands which were 1 m apart along the row and also 1 m between rows.
Protocol for Harvest and Dry Matter Analysis
Three destructive samplings at 6, 9 and 12 Months after Planting (MAP) and a final harvest were carried out. During each sampling, five cassava plants were harvested on alternate rows (i.e., 4th row at 6 MAP, 6th row at 9 MAP and 8th row at 12 MAP). Plant data recorded were:
| • | Fresh weight of leaves and stems (kg): These were determined after separating the leaves and stems from roots |
| • | Number of tuberous roots:. This was determined after separating tuberous roots from the fibrous roots |
| • | Fresh weight of tuberous roots (kg) |
| • | Total dry weight of leaves, stems and tuberous roots (kg plot-1): All the components of the total shoots or tuberous roots per plot were chopped up into tiny pieces. These were placed into a paper packet and dried in the oven at 65°C for five days. The dry-matter content (%) of shoots and roots were calculated as the ratio between fresh weight and dry weight: |
| • | Number of rotted tuberous roots: The percentage rot incidence was calculated by dividing number of rotted tubers by the total number of tubers and then, multiplying by 100 |
| • | Rot severity on tuberous roots: The percentage-rotted area covered by the pathogen was calculated by visual observation and by making longitudinal and cross sections on tuberous roots with rot |
The dry matter partitioning for each sampling was assessed by comparing the dry weight of tuberous roots with those stem and leaves. Comparing this over the 12 months period gave the trend of dry matter partitioning.
RESULTS
Cassava Root Rot Incidence and Severity in Ibadan and Sabongidda-Ora
The cassava root rot incidence and severity data obtained from a previous experiment preceding this one (Aigbe and Remison, 2009) were used for this study. The first year of the study, generally showed root rot to be significant only at 12 MAP in the two locations. TME-1, at 12 months after planting (MAP) in the Sabongidda-Ora humid forest had the highest root rot incidence of 53% and a 21% severity, while TMS 4(2)1425 and TMS 30572 had the lowest root rot incidences of 0 and 6.4% and severities of 0-0.5% at the Ibadan derived savanna (Table 1, 2).
The second year’s planting showed similar trend, except, root rot on TME-1 was also now significant at 9 MAP in Ibadan (Table 1, 2).
Cassava Dry Matter Partition to Stem, Leaves and Roots in Ibadan and Sabongidda-Ora Ibadan
At Ibadan and for all cultivars, the dry matter partition to leaves, stem and storage roots followed similar pattern for all the 3 genotypes in the different growth stages. Partitions to the roots were generally significantly more while partitions to leaves and stem were approximately equal with no significant difference except at the 12 months after planting when the partition to the leaves dropped both drastically and significantly behind that of stem.
| Table 1: | Percentage root rot incidence and severity of three cassava varieties at 6, 9 and 12 MAP* in Ibadan, Onne and Sabongidda-Ora** |
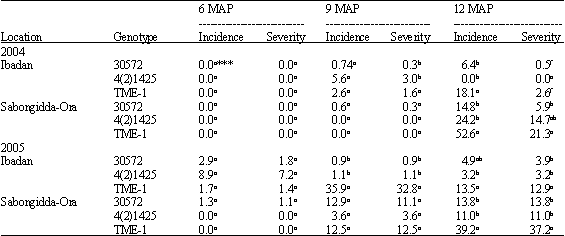 | |
| *MAP: Months after planting, **Data represent a mean of five stands, ***Values with the same letter in the same column under the same year/location are not significantly different at p = 0.05 | |
| Table 2: | Percentage root rot incidence and severity of three cassava varieties at 6, 9 and 12 MAP* across different locations** |
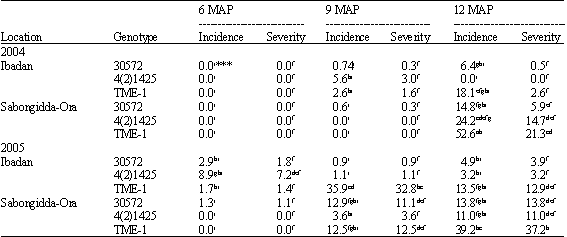 | |
| *MAP: Months after planting, **Data represent a mean of five stands, ***Values with the same letter in the same column are not significantly different at p = 0.05 | |
What was lost by the leaves was gained by the stem and storage roots (Table 3).
During the second year planting, dry matter partition to leaves, stem and storage roots for the 2 genotypes followed the same trend observed in first year (Table 3).
Sabongidda-Ora
Like at Ibadan, the dry matter partition to leaves, stem and storage roots at 6 and 9 months after planting, followed similar pattern for all the 3 genotypes. Partitions to the roots were generally significantly more while partitions to leaves and stem were approximately equal with no significant difference. But at 12 MAP unlike at Ibadan, no significant difference was observed between stem and roots; with an exception for TMS 4(2)1425. TME-1 dry matter partition to roots, 367.4 g at 12 MAP was the lowest and was significantly lower than the 804.1 g of TMS 4(2)1425; the highest recorded on roots at 12 MAP. The 804.1 g dry matter partition to roots of TMS 4(2)1425 was not significantly different from its 684.8 g dry matter at Ibadan (Table 3, 4).
| Table 3: | Dry matter partition to stem, leaves and roots of 3 cassava genotypes, under different agro ecological environments, 6, 9 and 12 months after planting in different years (2004 and 2005) |
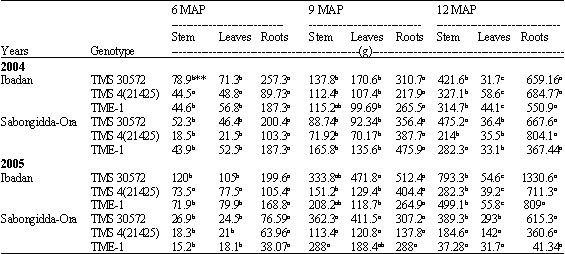 | |
| *Values with the same letter in the same row under the same MAP are not significantly different at p = 0.05 | |
| Table 4: | Dry matter partition to stem, leaves and roots of 3 cassava genotypes, 6, 9 and 12 months after planting across different locations** in different years (2004 and 2005) |
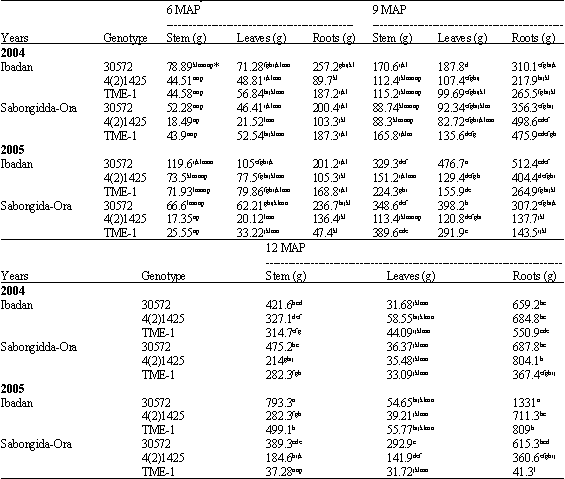 | |
| *Values with the same letter in the same column are not significantly different at p = 0.05, **Plots invaded by Bacteria Blight | |
During the second year planting at Sabongidda-Ora, dry matter partition to leaves, stem and storage roots for the 3 genotypes followed similar trend observed in first year. Dry matter partition to roots in TME-1 at 12 MAP was now reduced to a miserable 41.34 g; the lowest observed at 12 MAP for the two planting seasons (Table 3).
DISCUSSION
The dry matter partition trend observed at Ibadan; were by the partitions to the roots were generally significantly more, while the partitions to the leaves and stem were approximately equal with no significant difference except at the 12 months after planting when the partition to the leaves dropped significantly behind that of stem, can relatively be regarded as normal. This should be so because Ibadan derived savanna had insignificant root rot when compared to Sabongidda-Ora humid forest. With the exception of TMS 30572 and susceptible TME-1 and 12 MAP, the observation that the dry matter partition to roots in both locations and in all varieties at 12 MAP was significantly higher than those of stem and leaves is perfectly in order, since this is the most economical part of the plant at this stage.
The general observation that dry matter partition to leaves at 12 MAP dropped significantly behind those of stem and roots is to be expected since the stem and the roots are the critical portions of the cassava plant at this time rather than vegetative growth. Twelve months after planting is the ideal harvest time for cassava tuberous roots; the harvested roots are consumed or processed while the stems are used as planting materials for the next planting season. This is also in agreement with Ekanayake et al. (1998), who reported that partitioning of dry matter to the storage roots varies from almost zero during the early growth stages to nearly 80% of the daily dry matter production during the late growth stages. They reported the relationship between total dry weight of the plant and dry weight of the storage roots to be linear, suggesting the rate of root growth keeps pace with the rate of crop growth.
The fact that TME-1, with the highest root rot incidence and severity (52.6 and 21.3%, respectively) at 12 MAP in Sabongidda-Ora also had the lowest root dry matter (367.4 g) at 12 MAP, even lower than the 475.9 g recorded at 9 MAP, is an indicator that root rot activities reduces dry matter partition to roots. This result agree with the findings of other workers, who, though working with various crops and different pathogens, similarly observed that an increase in disease severity results in reduction of dry matter production. For instance, McLean (1926) reported a lower dry matter content in potato tubers after infection with secondary leaf-roll disease. Jordan et al. (2007), also reported a great decreased in root and shoot dry matter production after inoculation of winter barley plants with Pyrenophora teres in a glasshouse. During an experiment involving inoculation of radish with nodules inducing bacterium - arctic strain (N44), a 44% reduction in radish dry matter yield was observed (Antoun et al., 1998). The percentage dry weight of the storage roots of sweetpotato were similarly observed to be highest when Fusarium root rot was reduced as result of treatment with UV-C doses from 0 to 3.6 kJ m-2 (Stevens et al., 1999). Simpfendorfer et al. (1999) also observed that a 14-58% reduction in Phytophthora root rot severity of subterranean clover (Trifolium subterraneum L.) resulted into a corresponding 20-73% increase in dry matter production.
Since the dry matter partition to the stem of root rot susceptible TME-1 at 12 MAP at Sabongidda-Ora (282.3 g) and Ibadan (314.7 g) were significantly lower than those of root rot tolerance TMS 30572, both at Sabongidda-Ora (475.2 g) and Ibadan (421.6 g), it shows therefore that root rot also affect dry matter partition to stems. The reduction in dry matter partition to the roots of TME-1 (41.3 g) at 12 MAP in Sabongidda-Ora as result of root rot was responsible for it not differing significantly from the 37.3 g partition to stem, as would have been expected. Even though the root tissue selected for dry matter analysis was healthy looking, this however did not rule out the possibility of latent infection. This is because five distinct phases are involved in root rot diseases development; introduction, incubation, penetration, infection and root rot disease (Keinath et al., 2007). The last phase alone results in symptoms expression, the process of root rot disease development most likely has already commenced in the healthy looking root tissue of rot susceptible TME-1 processed for dry matter analysis.
CONCLUSION
In conclusion, this research has shown for the first time that high root rot incidence / severity reduce dry matter production and partitioning in cassava plants. There is a great potential for cassava for food particularly with increased population, market opportunities and recent policies aimed at reducing cereal imports. Effective control of cassava root rot will contribute towards the constancy of not only supply but of quality cassava to urban markets throughout the year and strengthen food security of rural communities thereby leaving the farmers rich.
There is need for more detailed study on the effect of different regimes of temperature, solar radiation and soil moisture on root rot and dry matter partitioning in a controlled environment. The molecular analysis of the pathogens / cassava cultivars involved should also be studied. Studies that will lead to increased understanding of the effect of other diseases on the dry matter of cassava roots are also desirable.
ACKNOWLEDGMENT
This research was supported by the International Institute of Tropical Agriculture.
REFERENCES
- Antoun, H., C.J. Beachamp, N. Goussard, R. Chabot and R. Lalande, 1998. Potential of Rhizobium and Bradyrhizobium species as plant growth promoting rhizobacteria on non-legumes: Effects on radishes (Rhaphanus sativus L.). Plant Soil, 204: 56-67.
Direct Link - Jordan, V.W.L., G.R. Best and E.C. Allen, 2007. Effects of Pyrenophora teres on dry matter production and yield components of winter barley. Plant Pathol., 34: 200-206.
CrossRef - McLean, W., 1926. Effect of leaf-roll disease in potatoes on the composition of the tuber and mother tuber. J. Agric. Sci., 16: 318-324.
CrossRef - Messiga, A.J.N.A., M. Mwangi, R. Bandyopadhyay and C. Nolte, 2004. The status of fungal tuber rots as a constraint to cassava production phytopathology in the Pouma district of Cameroon. Proceedings of the 9th Triennial Symposium of the International Society for Root and Tuber Crops, (TSISRTC`04), Mombasa, Kenya, pp: 42-42.
- Onyeka, T.J., A.G.O. Dixon and E.J.A. Ekpo, 2005. Field evaluation of root rot disease and relationship between disease severity and yield in cassava. Exp. Agric., 41: 357-363.
CrossRef - Stevens, C., V.A. Khan, J.Y. Lu, O. Adeyeye, L.P. Pusey and A.Y.A. Tang, 1999. Induced resistance of sweetpotato to Fusarium root rot by UV-C hormesis. Crop Protection, 18: 463-470.
CrossRef