
F. Chamandoosti
Department of Plant Pathology, Iranian Research Institute of Plant Protection, No. 1 Yaman Ave, Chamran Exp` Way, Tehran, Iran
Asian Journal of Plant Pathology
Year: 2009 | Volume: 3 | Issue: 1 | Page No.: 8-13







ABSTRACT
In this research, effect of different concentrations of oxalic acid (OAA) (0, 0.02, 0.2, 2 and 20 mM L-1) were studied on callusing and organogenesing of canola in order to possibility producing resistant canola plant to pathogen fungus Sclerotinia sclerotiorum (Lib.) de Bary by this pressure selection. 0-0.2 mM L-1 OAA had a positive effect on above reactions so that the mean diameter of calli and the mean number of regenerated roots and shoots on hypocotyl explants, also the mean number of explants that produced callus and de novo organs were increased. These results showed that the addition of OAA (0.0-20 mM L-1) in canola culture media (in vitro selection technique) lead to produce resistant canola plants to Sclerotinia sclerotiorum (Lib.) de Bary.
PDF Abstract XML References Citation
How to cite this article
F. Chamandoosti, 2009. In vitro Selection: A Novel Source of Resistance to Sclerotinia Stem Rot in Canola (Brassica napus L.). Asian Journal of Plant Pathology, 3: 8-13.
URL: https://scialert.net/abstract/?doi=ajppaj.2009.8.13
URL: https://scialert.net/abstract/?doi=ajppaj.2009.8.13
INTRODUCTION
As the world`s population continues to increase, food supplies must grow to meet nutritional requirements. One means of ensuring the stability and plentitude of the food supply is to mitigate crop losses caused by crop pathogens (Campbell et al., 2002). On the other hand breeding for stress tolerance by the traditional methods is time consuming procedure (Dorffling et al., 1993; Noman et al., 2004). Plant cell culture provides a unique opportunity to manipulate morphogenesis in a controlled environment, thus providing crop improvement a powerful, complementary tool. Since the late 1970s, the process of in vitro selection had been applied to several cell culture systems to generate mutants with useful agronomic traits such as disease resistance (Jayasankar and Gray, 2003). Sclerotinia sclerotiorum (Lib.) de Bary, an ascomycetous fungus causes disease on more than 400 host species (Boland and Hall, 1994; Rejanel and Stotz, 2004). This fungus casuses one of the most important diseases in canola (Brassica napus L.) in Iran. Oxalic acid that produced by Sclerotinia sp. has repeatedly been shown to be important in pathogenesis of this fungus (Rowe, 1993). Oxalate is an essential virulence factor of S. sclerotiorum because mutants, which are deficient in oxalate biosynthesis, are less pathogenic than wild-type fungus (Godoy et al., 1990). In contrast to wild-type fungus, oxalate-deficient S. sclerotiorum is unable to produce oxalate during infection of petals, which are an important source of inoculum in the field and during in vitro cultivation (Godoy et al., 1990; Jamaux et al., 1995; Rejanel and Stotz, 2004). Oil seed crop improvement by conventional breeding is severely restricted by the availability of a rather limited gene pool and by the time scale of most breeding programs, on the other hand there aren`t canola absolutely resistant cultivars in Iran. Plant cell tissue and organ culture provide a power tool for crop improvement by techniques such as in vitro selection, somaclonal variation and genetic engineering. In reality in vitro selection for disease resistance in plants is an alternative to genetic engineering (Jayasankar and Gray, 2003). In this study the possibility of using from OAA for producing resistant canola was studied by this system.
MATERIALS AND METHODS
This study conducted in Iranian Research Institute of Plant Protection, Tehran, Iran in Spring 2007 and 2008.
One canola (Brassica napus L.) cultivar, Pionner was used. In order to establish a stock of plants, seeds that were obtained from Oil seed Research and development Company were sterilized in a commercial sodium hypochlorite solution (with 5% available chlorine) for 8 min and were rinsed 4 times with sterile water. Then sterile seeds were cultured in MS (Murashige and Skoog, 1962) medium without Plant Growth Regulators (PGRs). Seven days later hypocotyls of (4-6 mm) seedlings were used as explant.
Culture Medium, Incubation Conditions, Evaluation Procedure and Statistical Analysis
The basal medium was included the (MS formulation), sucrose 3% (w/v) and agar agar 1% supplemented with PGRs (2,4_D, NAA, IBA, Kinetin, BA) and 0, 0.02, 0.2, 2 and 20 mM L-1 oxalic acid.
The media were adjusted to pH 4 (Thompson et al., 1995) and autoclaved at 121°C for 20 min. Cultures were maintained in 16 h photoperiod at 25±1°C. After establishment, cultures were subcultured at 5-6 weeks intervals on fresh media. And 2 months later the percentage of explants that remained alive and had callus, root and shoot evaluated. Experiment were set up in completely randomized design and repeated 3 times. Twelve treatments had 3 replications. Observation on the diameter of calli and the number of regenerated roots and shoots were recorded. Data were subjected to Mean±SD.
RESULTS
As was reported earlier by Chamandoosti et al. (2006) and Chamandoosti (2007) after two weeks of culture hypocotyl explants bigan to induced calli, roots and shoots in media with 6 mg L-1 2, 4-D and 2 mg L-1 BA, 1 mg L-1 NAA and 0.5 mg L-1 Kinetin and 1 mg L-1 IBA and 1 mg L-1 BA, respectively. This media supplemented by 0, 0.02, 0.2, 2 and 20 mM L-1 oxalic acid.
Effect of OAA on Induction and Growth of Calli
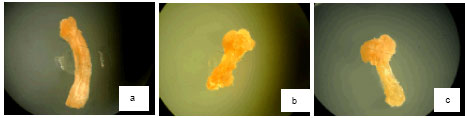
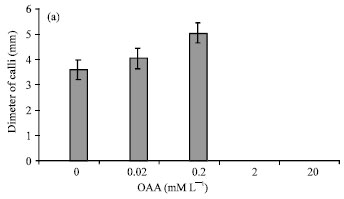
Characteristics of calli that produced after establishment of hypocotyl explants in media for callusing plus OAA was similar to calli that produced in medium without OAA (compact, yellowish and nonorganogene). In this medium with 0-0.2 mM L-1 OAA the growth of calli increased (30564±0.0385) (Fig. 2a). Diameter of calli decreased at presence of 2-20 mM L-1 OAA so that in 20 mM L-1 OAA all of explants died (Fig. 1a-c).
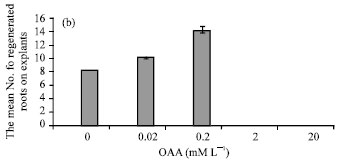
Effect of OAA on the Mean Number of Regenerated Roots per Explants and the Mean Number of Explants that Rooted
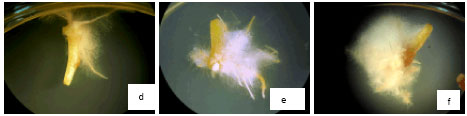
Rooting in media with different concentrations of OAA was alike rooting in medium free from OAA (1 mg L-1 NAA plus 0.5 mg L-1 Kinetin), so that in this media two weeks after establishment of explants at first very light yellow and compact calli were appeared then these calli produced regenerated roots very well. Increment of OAA concentration (0-0.2 mM L-1) caused that either the mean number of regenerated roots per explants or the mean number of explants that rooted were more rather than medium with 0 mM L-1 OAA (14.235±0.050), (Fig. 2b) and (Fig. 1d-f). In rooting medium (1 mg L-1 NAA plus 0.5 mg L-1 Kinetin ) with 2 mM L-1 OAA opposite of callusing and shooting, not only die explants but also produced regenerated few roots.
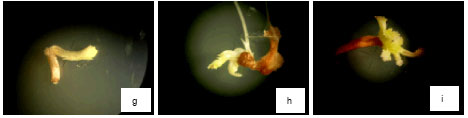
| Fig. 1: | Effect of oxalic acid on callusing, rooting and shooting of Brassica napus L. Effect of OAA on induction and growth of calli in 0 mM L-1 OAA (a), 0.02 mM L-1 OAA (b), 0.2 mM L-1 OAA (c), the means number of regenerated roots in 0 mM L-1 OAA (d), 0.02 mM L-1 OAA (e), 0.2 mM L-1 OAA (f), the means number of regenerated shoots in 0 mM L-1 OAA (g), 0.02 mM L-1 OAA (h) and 0.2 mM L-1 OAA (i) |
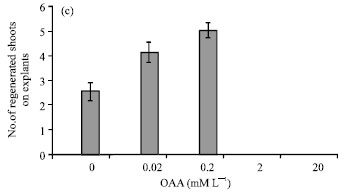
Effect of OAA on the Mean Number of Regenerated Shoots Per Explants and the Mean Number of Explants that Produced Shoot
As mentioned above in 2-20 mM L-1 OAA, (in shooting and callusing medium) explants whited and died therefore produced any regenerated shoots. In 0-0.2 mM L-1 OAA similar callusing and rooting, shooting were increased also. In these medium (either with OAA or without OAA) at first explants produced calli. These calli were white to light yellow and able to regenerating shoots. The increment that were observed in shooting medium with 0-0.2 mM L-1 OAA included of the mean number of regenerated shoots per explants (Fig. 2c) and the mean number of explants that produced shoot (Fig. 1g-i).
DISCUSSION
Results of this research demonstrated that in 20 mM L-1 OAA all of explants died and no callusing, rooting and shooting were observed. Thompson et al. (1995) stressed that 20 mM L-1 OAA have lethal effect for canola. Theses researchers isolated a gene coding for the enzymes oxalate oxidaze from barely roots and introduced into oilseed rape as a means of degrading oxalic acid. This report describes the production of several transgenic plants of oilseed rape and the characterization of these plants by southern, western and enzyme activity assays. Plants were shown to contain an active oxalate oxides enzyme and were tolerant of exogenously supplied 20 mM L-1 oxalic acid.
As were explained earlier oxalic acid that produced by fungus pathogen Sclerotinia sclerotiorum (Lib.) de Bary have a primary role in the pathogenecity of this fungus. Evidence for this comes from studies on mutant strains of this fungus deficient in oxalate production, which are avirulant. For example an oxalate degrading enzyme, oxalate oxidase (OxO), in transgenic soybean (Glycine max L. Merr.) has reduced pathogen growth in indoor seedling studies (Cober et al., 2003). This research showed that susceptibility of canola in in vitro conditions is similar to susceptibility whole plant in in vivo conditions.
| Fig. 2: | Effect of 0.02-20 mM L-1OAA on the (a) Mean (±SD) diameter of calli, (b) Mean (±SD) number of regenerated roots on explants and (c) (±SD) Mean number of regenerated shoots on explants in Brassica napus L. |
One of the interesting results of this research is resistance of hypocotyl explants of 7 days old seedling of canola in 0-0.2 mM L-1 OAA, indeed in these concentrations of OAA the mean diameter of calli and the means number of regenerated roots and shoots on explants was higher than in free from OAA media. It is possible that by gradually adaptation hypocotyl explants of canola in callusing culture media with 0.2, 2 and 20 mM L-1 OAA then transferring of calli to organogene media (1 mg L-1 IBA, 1 mg L-1 BA and 1 mg L-1 IBA for shooting and rooting, respectively) (Chamandoosti et al., 2006; Chamandoosti, 2007), tolerant or resistant whole plants were resulted. Gengenbach and Green (1975) produced also maize callus resistant to Helminthosporium maydis for the first time. This technology (in vitro selection) had been applied to several cell culture systems to generate mutants with useful agronomic traits such as diseases resistance. However, the promise of genetic engineering technology and some early failures among the in vitro selected plants stifled research in this area. Recent advances in molecular characterization of stress-related responses and the emergence of sensitive molecular analytical tools have reinvigorated research on in vitro selection. This technology is easy to use and not encumbered by intellectual property issue and social concerns currently inhibiting development of transgenic crops. Thus it is an attractive complement to existing crop improvement strategies (Jayasankar and Gray, 2003). American chestnut wildtype callus tissue and callus tissue transformed with an oxalate oxidase gene were cultured on media containing various concentrations of oxalic acid. Thermogravimetric analysis (TGA) was employed to test the cellulose, hemicellulose and lignin content of the calli. In the presence of oxalic acid, wildtype tissues demonstrated a significant decrease in lignin content and increase in cellulose content, while transformed tissues did not. Transforming American chestnut with an oxalate oxidase gene may prove to be useful in preventing changes in the cell wall composition caused by the oxalate produced during C. parasitica infection and could possibly enhance resistance (Welcha et al., 2007).
Kumar et al. (2008) described a system for efficient plant regeneration via organogenesis and somatic embryogenesis of safflower (Carthamus tinctorius L.) cv. NARI-6 in Fungal Culture Filtrates (FCF)-treated cultures. FCF was prepared by culturing Alternaria carthami fungal mycelia in selection medium for host-specific toxin production. They produced nine plantlets from organogenesis and 24 plantlets from somatic embryogenesis were selected as FCF-tolerant. Alternaria carthami fungal spores (5x105 spores mL-1) sprayed on the leaves of FCF-tolerant plants showed enhanced survival rate over control plants, which plants were more susceptible to fungal attack.
In vitro selection have been used not only in response to pathogen infection but also in many cases in response to abiotic stresses. Kirti et al. (1991) tried to produce of sodium-chloride-tolerant Brassica juncea plants by in vitro selection at the somatic embryo level. In their experiments embryogenesis was drastically inhibited and the somatic embryos failed to germinate when sodium-chloride was added to the induction medium. They overcame this difficulty by inducing somatic embryos on sodium-chloride free medium and subsequently challenging them to restrictive conditions and they proved this procedure is effective. Mercado et al. (2000) used of growth of apical stem sections and adventitious organogenesis to evaluate salinity tolerance in cultivated tomato (Lycopersicon esculentum L.) and resulted that this approach may not be a reliable tool to evaluate salt tolerance in tomato.
Results of this research clearly demonstrated that we can use from the gradually increment of oxalic acid concentration in canola callus culture media as a selection pressure to producing tolerant lines and shows that this procedure is effective and also 0.2 mM L-1 OAA is the suitable concentration for beginning (gradually adaptation) for the first time.
ACKNOWLEDGMENT
I express my thanks to Prof. Afshari Azad for his valuable help in establishment a tissue culture laboratory.
REFERENCES
- Boland, G.J. and R. Hall, 1994. Index of plant hosts of Sclerotinia sclerotiorum. Can. J. Plant Pathol., 16: 93-108.
Direct Link - Campbell, M.A., H.A. Fitzgerald and P.C. Ronald, 2002. Engineering pathogen resistance in crop plants. Transgen. Res., 11: 599-613.
CrossRefDirect Link - Chamandosti, F., A. Majd and S. Mehrabian, 2006. In vitro plant regeneration from callus of cotyledons in canola (Brassica napus L.). Pak. J. Biol. Sci., 9: 302-306.
CrossRefDirect Link - Chamandoosti, F., 2007. Effect of sodium chloride on establishment of callus and organogenesis in Brassica napus L. Pak. J. Biol. Sci., 10: 3880-3884.
CrossRefPubMedDirect Link - Cober, E.R., S. Riouxb, I. Rajcan, P.A. Donaldson and D.H. Simmonds, 2003. Partial resistance to white mold in a transgenic soybean line. Crop Sci., 43: 92-95.
Direct Link - Dorffling, K., H. Dorffling and G. Lesselich, 1993. In vitro selection and regeneration of hydroxyprolin-resistant lines of winter wheat with increased prolin content and increased frost tolerance. J. Plant Physiol., 142: 222-225.
Direct Link - Gengenbach, B.G. and C.E. Green, 1975. Selection of T-cytoplasm maize callus cultures resistant to Helminthosporium maydis race T pathotoxin. Crop Sci., 15: 645-649.
Direct Link - Godoy, G., J.R. Steadman, M.B. Dickman and R. Dam, 1990. Use of mutants to demonstrate the role of oxalic acid in pathogenicity of Sclerotinia sclerotiorum on Phaseolus vulgaris. Physiol. Mol. Plant Pathol., 37: 179-191.
CrossRef - Jamaux, I., B. Gelie and C. Lamarque, 1995. Early stages of infection of rapeseed petals and leaves by Sclerotinia sclerotiorum revealed by scanning electron microscopy. Plant Pathol., 44: 22-30.
CrossRef - Jayasankar, S. and D.J. Gray, 2003. In vitro selection for diseases resistance in plants-an alternative to genetic engineering. AgBiotechNet, 5: 1-5.
Direct Link - Kirti, P.B., S. Hadi, P.A. Kumar and V.L. Chopra, 1991. Production of sodium-chloride-tolerant Brassica juncea plants by in vitro selsction at the somatic embryo level. Theor. Applied Genet., 83: 233-237.
CrossRefDirect Link - Kumar, J.V., B.D. Ranjitha Kumari, G. Sajatha and Enrique Castano, 2008. Production of plants resistant to Alternaria carthami via organogenesis and somatic embryogenesis of safflower cv. NARI-6 treated with fungal culture filtrates. Plant Cell Tissue Organ Culture, 93: 85-96.
CrossRefDirect Link - Mercado, J.A., M.A. Sancho-Carrascosa, S. Jimenez-Bermudez, R. Peran-Quesada, F. Pliego-Alfaro and M.A. Quessada, 2000. Assessment of in vitro growth of apical stem section and adventitious organogenesis to evaluate salinity tolerance in cultivared tomato. Plant Cell Tissue Organ Cult., 62: 101-106.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Noman, S.H., L.D. Shaaban, E.S.A. Hashem and E.E. Seleem, 2004. In vitro selection for water strees tolerant callus line of Helianthus annus L. cv. Myak. Int. J. Agric. Biol., 1: 13-18.
Direct Link - Rejanel, G. and H. stotz, 2004. Oxalate production by Sclerotinia sclerotiorum deregulates Gurd cells dyring infection. Plant Physiol., 136: 3703-3711.
CrossRefDirect Link - Rowe, D.E, 1993. Oxalic acid effects in exudates of Sclerotinia trifoliorum and S. sclerotiorum and potential use in selection. Crop Sci., 33: 1146-1149.
Direct Link - Thompson, C., J.M. Dunwell, C.E. Johnstone, V. Lay and J. Ray et al., 1995. Degradation of oxalic acid by transgenic oilseed rape plants expressing oxalate oxidase. Euphytica, 85: 169-172.
CrossRef - Welcha, A., A.J. Stipanovicb, C.A. Maynardc and W.A. Powella, 2007. The effects of oxalic acid on transgenic Castanea dentata callus tissue expressing oxalate oxidase. Plant Sci., 172: 488-496.
CrossRef