
T. Deng-Feng
Maize Research Institute, Sichuan Agricultural University, Ya`an, Sichuan 625014, China
Z. Fan
Maize Research Institute, Sichuan Agricultural University, Ya`an, Sichuan 625014, China
Z. Zhi-Ming
Maize Research Institute, Sichuan Agricultural University, Ya`an, Sichuan 625014, China
W. Yuan-Qi
Maize Research Institute, Sichuan Agricultural University, Ya`an, Sichuan 625014, China
Y. Ke-Cheng
Maize Research Institute, Sichuan Agricultural University, Ya`an, Sichuan 625014, China
R. Ting-Zhao
Maize Research Institute, Sichuan Agricultural University, Ya`an, Sichuan 625014, China
Y. Guang-Sheng
Maize Research Institute, Sichuan Agricultural University, Ya`an, Sichuan 625014, China
P. Guang-Tang
Maize Research Institute, Sichuan Agricultural University, Ya`an, Sichuan 625014, China
Asian Journal of Plant Pathology
Year: 2009 | Volume: 3 | Issue: 1 | Page No.: 1-7







ABSTRACT
The inheritance and gene effects on resistance to F. moniliforme in maize were investigated using 6 relevant generations of a cross (R15xYe 478) including 2 parents, F1, F2, BCP1 and BCP2 by the mixed major gene plus poly-genes in genetic model of quantitative traits. The female parent R15, developed by the Maize Research Institute in the Sichuan Agricultural University is highly resistant to ear rot. Ye 478 is an inbred line with a high combining ability, but it is susceptible to many diseases. The frequency distribution of disease severity in segregating populations showed characteristics of a mixed normal distribution, which indicated the inheritance of resistance followed major genes plus poly-genes model. Twenty-four genetic models were established, which could be classified into five types: one major gene, two major genes, polygene, one major gene plus polygene and two major genes plus polygene. Results showed the genetic model E-3 was the most suitable model for the trait and the resistance was controlled by two additive major genes plus additive-dominance polygene. Finally, the results also revealed that agronomic traits investigated such as spike length, spike width, spike rows and kernel depth, etc. had less correlation to resistance to F. moniliforme maize ear rot, which showed that resistance to maize ear rot was mainly controlled by genetic factors and indirect agronomic traits can not be used as a selection index in breeding maize varieties for resistant to F. moniliforme ear rot.
PDF Abstract XML References Citation
How to cite this article
T. Deng-Feng, Z. Fan, Z. Zhi-Ming, W. Yuan-Qi, Y. Ke-Cheng, R. Ting-Zhao, Y. Guang-Sheng and P. Guang-Tang, 2009. Analysis of Gene Effects and Inheritance of Resistance to Fusarium moniliforme Ear Rot in Maize. Asian Journal of Plant Pathology, 3: 1-7.
URL: https://scialert.net/abstract/?doi=ajppaj.2009.1.7
URL: https://scialert.net/abstract/?doi=ajppaj.2009.1.7
INTRODUCTION
Ear rot, caused by F. moniliforme, is one of the most destructive diseases of maize in the world, especially in the southwest of China. The general incidence was 5-7%, while sometimes it exceeds 50% in susceptible lines (Wu et al., 1999). Development and cultivation of resistant hybrids are the most effective strategy to control F. moniliforme ear rot in maize. In recent years, a high incidence of ear rot has been reported in Sichuan Province of China. Infected maize caused grain yield losses, which seriously restricted the development of maize (Ren et al., 1993; Chen and Wen, 2002).
Several pathogens are known to cause maize ear rot and this complicates plants resistance mechanism. Currently here are few reports on the inheritance of ear rot resistance in maize. Few resistant genetic resources have been identified and the mechanisms of resistance have also not been clearly defined. To identify and efficiently transfer the genes controlling ear rot resistance to susceptible maize genotypes, it is necessary to understand the mode of inheritance of resistance.
The aim of this study was to identify resistance mechanism and inheritance of resistance to F. moniliforme ear rot in maize using parents R15 and Ye478 and their progeny F1, F2, BC1 and BC2 derived from a cross between them. The result is help to establish a robust basis for breeding maize varieties resistant to F. moniliforme ear rot.
MATERIALS AND METHODS
Plant Material
The materials included 6 generations: parents (R15 and Ye 478) and their progenies F1, BC1, BC2 and F2, derived from a cross between R15 and Ye 478. The female line used in the cross was R15, which is resistant to the pathogen. The male parent was the susceptible line Ye 478 with good agronomic characteristics.
Field Experiments
The field trials were carried out using random block design and individual plants grown during the Spring of 2004 at DuoYing in Ya`an, Sichuan Province. The materials were grown in 10x10 lattice designs with three replications each. The distance between row spacing was 0.8 m long and in-row spacing was 0.22 m. The individual plants for R15, Ye478, F1, BC1, BC2 and F2 were 100, 100, 150, 800, 800 and 1000, respectively.
Plants were artificially inoculated and its effects were assessed by disease severity for the whole generation. The F. moniliforme was donated by the Chinese Academy of Agricultural Sciences. The pathogen was cultured on potato dextrose agar with subsequent sub-culturing for the production of field inoculums. A spore suspension of 2.5~3.0x106 spores mL-1 was prepared at the time of field inoculations. The plants were inoculated using the sponge and nail-punch technique, where, the husk leaves in the middle of the ear were punctured to introduce the inoculum ten days after female flowering (Chen and Wen, 2002; Pan and Zhang, 1987; Huang and Zheng, 1990). At harvest, the ear number was investigated for R15, Ye 478, F1, BC1, BC2 and F2 were 45, 72, 128, 611, 461 and 820, respectively.
Data Scoring and Analysis
When kernels were matured, plant height and ear height were measured in each plot. After harvesting and wind drying, ear length, ear width, grain depth, ear rows, grain numbers and infected grain numbers for single ear were investigated in laboratory.
At harvest, the inoculated ears were evaluated individually based on a severity scale representing the percentage of infection (Chen and Wen, 2002; Huang and Zheng, 1990; Ma et al., 1998). A 1-6 scale was used with 0 = 0-5%, 1 = 5.1-15%, 2 = 15.1-25%, 3 = 25.1-50%, 4 = 50.1-75%, 5 = 75.1-100% infection of the ear.
The infected degree for each generation was evaluated individually based on mean severity scale representing the percentage of infection (Chen and Win, 2002; Ma et al., 1998). A 1-4 scale was used to evaluate the degree. X signed the mean severity scale. High resistance (X≤0.5); Middle resistance (0.5
The number of ears harvested for each generation was recorded (NMT), the number of ears in each division of the classification (i) was noted (Mgi), the highest severity classification was noted (hi), the mean severity degree (MD) for each generation and then the disease severity index (MP) were calculated using the following formula:
(1) |
(2) |
Analysis of phenotypic data were performed using Excel and DPS software and the united variance analysis of six generations method provided by Mather and Jinks (1977, 1982), Gai and Wang (1998a, b), Gai et al. (2000) and Wang et al. (2000).
RESULTS AND DISCUSSION
Numbers of Genes Controlled the Resistance to F. moniliforme
The numbers of infected grains of each ear for P1, P2 and F2 populations were transformed into the reverse sine functions and then the mean index and variance were calculated (Table 1).
Numbers of genes controlling resistance to F. moniliforme were determined based on the method described by Ma (1982). The results revealed that at least 5 pairs genes were correlated with the resistance to F. moniliforme ear rot in maize.
Analysis for Average Severity Degree and Genetic Effect
The percent of uninfected grains were transformed into the reverse sine functions and then the adaptability of additive-dominant model was verified using single scale method.
A, B and C value were calculated based on the average numbers of uninfected grains in 6 generations by ABC model method, the result showed that difference between A, C and zero was significant at the 1% probability level and the genetic model did not accord with additive- dominant model (Table 2).
A = -25.168±2.142, B = -0.887±4.296, C = 24.391±5.188 |
Therefore, the whole average value [m], additive effect [d], dominate effects [h], additivexadditive [i], additivexdominate [j] and dominatexdominate [l] were calculated based on single scale of additive-dominance-epistatic method described by Jinks and Jones (1958). The results revealed that [h], [i] and [l] were not significant at the 1% probability level. We further gradually eliminated [i] with lowest t value and calculated the other five genetic parameters using composite scale of additive-dominance-epistatic method. The results showed that resistance to F. moniliforme was accorded with additive-dominance-epistatic model and [l] was not significant at the 1% probability level. After [l] was eliminated, [l], [M], [d], [h] and [j] were determined by composite scale of additive-dominance-epistatic model (Table 3). The results in Table 7 indicated that [d], [h], [j] were both significant at the 1% probability level. The [d] was larger than [h] and [j] was bigger than [d] or [h] which indicated that addictive effect and additivexdominance-epistatic effect were dominant in the genetic effect of resistance to F. moniliforme maize ear rot.
Analysis the Mixed Model of Major Genes Plus Polygene and Estimate Parameters
Twenty-four genetic models were established, which could be classified into five types: one major gene (A), two major genes (B), polygene (C), one major gene plus polygene (D) and two major genes plus polygene (E).
| Table 1: | The mean and variance of disease severity in P1, P2, F2 populations |
 | |
| Table 2: | Statistical parameters of 6 generations and genetic effects |
 | |
| Table 3: | Analysis genetic effect by composite scale of additive-dominance-epistatic genetic model |
 | |
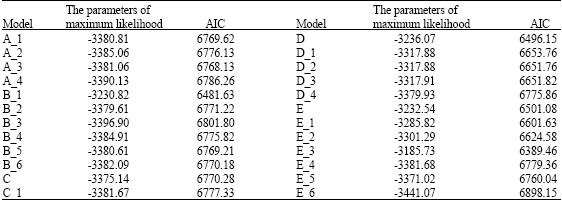
| Table 4: | The parameters of maximum likelihood and AIC by IECM method |
 | |
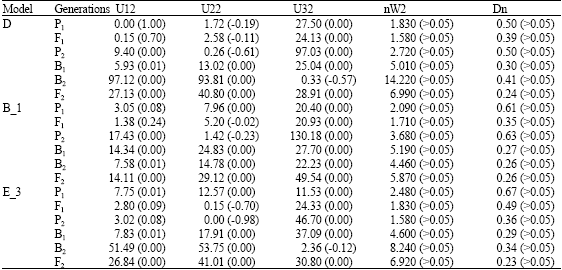
| Table 5: | The estimated adaptability of D, B_1 and E_3 model |
 | |
The maximum likelihood functions (AIC) of each model was estimated based on the reverse sine function in 6 generations according to the mixed major gene plus poly-gene in genetic model of quantitative traits (IECM) (Gai et al., 2003). The results in Table 4 are shown that the maximum likelihood function (AIC) of E_3 model (two additive genes plus additive plus dominant polygene) was smallest, which indicated model E_3 was the most suitable model for the trait and the next optimal models were model B_1 (two pairs genes - dominance-epistatic) and model D (one pairs additive genes-dominance major genes+additive-dominance-epistatic).
The adaptability of three putative models D, B_1 and E_3 were verified (Table 5). 15 parameters in model B_1 showed significant at the 5% probability level, 13 parameters in model E_3 and 12 parameters in model D, thus the putative model E_3 was selected the most suitable model for the trait because of its smallest AIC. Subsequently, the maximum likelihood value and the genetic parameters were estimated in E_3 model (Table 6, 7). The genetic parameters revealed that resistance to F. moniliforme maize ear rot was controlled by two major additive major genes plus additive- dominance polygene (Table 7).
| Table 6: | The maximum likelihood value estimated in E_3 model |
 | |
| Table 7: | The genetic parameters estimated in E_3 model |
 | |
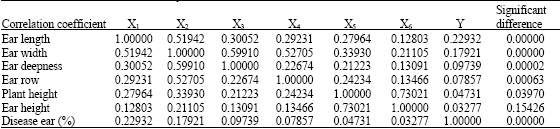
| Table 8: | The result of correlation analysis |
 | |
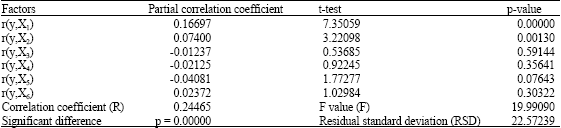
| Table 9: | The result of partial correlation analysis |
 | |
The actions of two additive major genes were reverse and positive effect was bigger than negative effect. The additive effect and dominant effect of polygene were also opposite and positive effect of additive genes was bigger than negative effect of dominant genes.
Analysis Correlation and Path Coefficient
The results indicated that most of these agronomic traits were not correlated with resistance. Only partial correlation coefficients between ear length, ear width and resistance were significant with very low value. This suggest that resistance to F. moniliforme should be directly evaluated by resistance itself rather than other agronomic traits in practical breeding program (Table 8-10).
Genetics of Resistance to F. moniliforme Ear Rot
Based on the results derived from scale method and IECM method (Gai et al., 2003), we can draw a reasonable conclusion that resistance to F. moniliforme ear rot is a quantitative trait and controlled by major genes plus polygene.
| Table 10: | Path coefficient analysis |
 | |
Genetic effects were main the additive of major genes, additive-dominance effect of polygene and additivexdominance-epistatic of polygene. The additive effects e both negative (da = -11.21) and s positive (db = 12.63). Positive effect was larger than negative effect, so resistance to ear rot exhibited positive additive effects ([d] = 16.33). Dominant effect exhibited negative effects ([h] = -14.09), which was verified by additivexdominance-epistatic model. The results of this study are consistent with the multigenic, quantitative nature of F. moniliforme ear rot resistance in corn (Reid et al., 1994). Therefore, the resistance materials may be utilized in breeding for the resistant varieties and for the development of resistant hybrids which are effective strategies to control F. moniliforme ear rot. Resistance gene was readily extracted from resistant germplasm because genetic effects were main additive effects for maize ear rot. At the same time, resistance to F. moniliforme ear rot also exhibited dominance and additivexdominance-epistatic, thus parents should be resistant to F. moniliforme when used to hybrid.
Strategies for Breeding the Resistance Varieties
Correlation analysis showed that these agronomic natures were not correlated with resistance. Therefore, selecting based on phenotypic trait was not suitable for breeding the resistance varieties. Inoculation of pathogen and selecting a close linkage marker were the effective strategy. The results provide excellent staring points for building an understanding of resistance to F. moniliforme infection in corn at genetic level and lay a basis for Marker-Assisted Selection (MAS) for the resistance species.
Application of Resistant Material R15 in Breeding Maize in Southwest of China
In the study, the female parent R15 is an inbred line with suitable plant height, narrow leaf, robust stem and highly resistant to ear rot, developed by the Maize Research Institute in the Sichuan Agricultural University (data not shown). Ye478 is an inbred line with excellent agronomic characters and a high combining ability, but it is susceptible to many diseases. Therefore, resistant gene could be utilized from R15 and transferred into elite corn lines from Ye478. In a word, successful development and application of R15 with highly resistant to ear rot would further contribute to breeding the resistant varieties in Sichuan Province and even Southwest in China.
ACKNOWLEDGMENTS
This study was supported by the National Natural Science Foundation of China (No. 30571173), the Beijing Natural Science Foundation (No. YZPT02-06) and the National 863 Foundation Project (No. 2006AA100103) and the Program for Changjiang Scholars and Innovative Research Teams in University of China (IRT0453).
REFERENCES
- Chen, X.J. and C.J. Wen, 2000. Preliminary study of maize ear rot in Sichuan. J. Southwest Agric. Univ., 24: 21-25.
Direct Link - Gai, J.Y. and J.K. Wang, 1998. Identification of major gene and polygene mixed inheritance model from backcrosses of F2: 3 families. Acta Agronomica Sinica, 24: 402-409.
Direct Link - Gai, J.Y., Y.M. Zhang and J.K. Wang, 2000. A joint analysis of multiple generations for QTL models extended to mixed two major genes plus polygene. Acta Agron. Sinica, 26: 385-391.
Direct Link - Gai, J.Y. and J.K.Wang, 1998. Identification and estimation of a QTL model and its effects. J. Theor Applied Genet., 97: 1162-1168.
CrossRef - Huang, C.L. and C.G. Zheng, 1990. Studies on some problems of identifying of opaque-2 maize to Fusarium moniliforme ear rot. Sci. Agric. Sinica, 23: 12-20.
Direct Link - Jinks, J.L. and R.M. Jones, 1958. Estimation of components of heterosis. Genetic, 43: 223-234.
PubMed - Ma, B.Y., S.S. Long, Y.L. Li and D.C. Li, 1998. Identification and pathogenicity of the pathogens of corn ear or kernel rot. J. Plant Prot., 25: 300-304.
Direct Link - Reid, L.M., D.E. Mather, A.T. Bolton and R. I. Hamilton, 1994. Evidence for a gene for silk resistance to Fusarium graminearum Schw. ear rot of maize. J. Heredity, 85: 118-121.
Direct Link - Ren, J.P., X.L. Wu, Z.C. Pang, X.W. Zhang and X.C. Liu, 1993. Preliminary study of maize ear rot. J. Maize Sci., 1: 75-79.
Direct Link - Wu, J.Y., Z.Y. Xi and J.Y. Gai, 1999. Advance in genetics and breeding of resistance of maize to disease. J. Maize Sci., 7: 6-11.
Direct Link - Wang, J.K., J.Y. Gai and Y.M. Zhang, 2000. Identification of two major genes plus polygenes mixed inheritance model of quantitative traits in B1 and B2 and F2. J. Biomathematics, 15: 358-366.
Direct Link