
Louis K. Prom
USDA-ARS, Southern Plains Agriculture Research Center, 2765 F and B Road, College Station, TX 77845
Asian Journal of Plant Pathology
Year: 2007 | Volume: 1 | Issue: 1 | Page No.: 12-17







ABSTRACT
The effect in vitro of washed and crushed anther/pollen and stigma extracts from six sorghum lines with different levels of ergot susceptibility on ergot conidial germination was determined. Conidia plated on water agar amended with either anther/pollen or stigma extracts generally stimulated higher rates of conidia germination than conidia plated on water agar alone. However, conidia germinations were suppressed on water agar plates amended with washed anther/pollen extracts from Novartis KS310, washed stigma extracts from NC+7W97 and crushed stigma extracts from NC+8R18. Compared to the other plates with stigma extracts, plates containing extracts from male-sterile line, ATx623 stimulated up to 227% higher conidia germination over the control plates. Overall, plates amended with anther/pollen extracts had significantly higher rates of germinated conidia than those containing stigma extracts. Overall, anther/pollen and stigma extracts tended to stimulate ergot conidia germination. However, components in the anther/pollen or stigma extracts from some of the sorghum lines exhibited the capacity to suppress spore germination. The critical factors from these floral components could be identified and used to minimize the impact of the disease.
PDF Abstract XML References Citation
How to cite this article
Louis K. Prom, 2007. Claviceps africana Spore Germination as Influenced by Components of Sorghum Floral Extracts. Asian Journal of Plant Pathology, 1: 12-17.
URL: https://scialert.net/abstract/?doi=ajppaj.2007.12.17
URL: https://scialert.net/abstract/?doi=ajppaj.2007.12.17
INTRODUCTION
Globally, ergot of sorghum (Sorghum bicolor (L.) Moench) is caused by three Claviceps species (anamorph Sphacelia sorghi McRae) (Bandyopadhyay et al., 1998; Frederickson and McLaren, 2000; Tsukiboshi et al., 2001). C. africana Frederickson et al. (1993) found in all regions where the disease occurs, C. sorghi Kulkarni et al. (1976) is found in Asia and Africa, while C. sorghicola Tsukiboshi, Shimanuki, and Uematsu is confined in Japan (Bandyopadhyay et al., 1998; Frederickson and McLaren, 2000, Tsukiboshi et al., 2001). In the USA, sorghum ergot is caused by C. africana (Isakeit et al., 1998; Paoutová et al., 2000). All the ergot pathogens infect the unfertilized ovaries by penetrating primarily through the stigma and sometimes via the style and ovary wall (Futrell and Webster, 1965; Bandyopadhyay et al., 1998). Sorghum lines exhibit different levels of susceptibility to the ergot pathogens (Bandyopadhyay et al., 1998). Male-sterile sorghum lines used for hybrid seed production are most vulnerable to infection because of their dependence on other sorghum plants for their pollen sources and environmental conditions, which may restrict the period when pollen is available for fertilization (Futrell and Webster, 1965; Bandyopadhyay et al., 1998).
Response to ergot by sorghum lines depends on how fast their stigmas, the primary sites for infection are penetrated and ovaries fertilized by pollen grains (Bandyopadhyay et al., 1998). Studies have shown that sorghum lines with stigmas that remain exposed for longer periods of time before flowering are more susceptible to ergot infection than lines whose stigmas are pollinated before the glumes open (Bandyopadhyay et al., 1998).
Thus, management strategies against the ergot pathogens, so far have relied on protecting the exposed stigmas by fungicides, synchronizing of flowering in A- and R-lines in hybrid seed production`fields, stagger planting so that flowering occurs in the dry season and using lines that efficiently pollinate and fertilize the ovaries (Futrell and Webster, 1965; Frederickson and Leuschner, 1997; Bandyopadhyay et al., 1998; Tsukiboshi et al., 2001; Prom and Isakeit, 2003).
Little or no information is available on the role of stigma secretions and extracts from other floral structures of sorghum in stimulating or inhibiting ergot spore germination. This area of research is significant because there are some commonalities between sorghum pollen/stigma and C. africana/stigma interactions (Komolong et al., 2003). Chinnadurai et al. (1970) noted that slide culture medium amended with stigmatic exudates from susceptible lines had significantly higher numbers of germinated spores of S. sorghi than medium containing exudates from resistant lines. Exudates from the susceptible lines were shown to contain large amounts of malic acid, succinic acid, arginine and aspartic acid, whereas exudates from the resistant lines had tartaric acid and tyrosine which were found to be inhibitory to ergot spore germination (Chinnadurai et al., 1970). Also, Bhuiyan et al. (2002) noted greater rates of germinated C. africana conidia on 1.5% water agar amended with extracts from boiled sorghum spikelets than on water agar alone.
This present study was undertaken to 1) determine the effects of crude stigma and anther/pollen extracts from six sorghum lines on C. africana conidia germination.
MATERIALS AND METHODS
This study was conducted at the USDA-ARS, Southern Plains Agriculture Research Center and at the Texas A and M Agricultural Research Farm, College Station, Texas in 2004.
Cultivars
Four hybrids: ‘ATx399xRTx 430’, ‘Novartis KS310’, ‘NC+7W97’ and ‘NC+8R18’, a male-sterile line ATx 623 and IS8525 cultivar with different levels of tolerance to C. africana were used in this study. Seeds of the sorghum lines were planted in 20 cm diameter plastic pots containing Metro-Mix 200 growing medium (Scotts-Sierra Horticultural Products Company, Maryland, OH) in the greenhouse at 25°C±2 and at the Texas A andM Agricultural Research Farm near College Station.
Stigma Extract
Fully emerged stigmas or those in flower buds before anthesis or pollen shed were excised using a pair of forceps, put in sterile vials and stored at -7°C. A 0.5 g sample of stigmas of each sorghum line was immersed in separate vials containing 8 mL sterilized distilled water and vortexed for 3-4 min. The supernatants of the washing were filtered through a 0.45 μm Nalgene filter membrane (J.H. Berge, Inc., South Plainfield, NJ). The washed stigmas were then ground in 8 mL sterilized distilled water with a porcelain mortar and pestle (J.H. Berge, Inc., South Plainfield, NJ ). Supernatants from the homogenized stigmas were filtered though a 0.45 μm Nalgene filter membrane.
Anther/Pollen Extract
Anther/pollen was obtained from panicles that were covered with paper bags at flower initiation until the end of anthesis. Panicles were shaken to dislodge the anther and pollen grains. The rest of the protocol was similar to that of the stigma-extract method. Aliquots of the stigma or pollen/anther extracts equivalent to 0.5, 1.0 and 2.0% were mixed with water agar culture medium. Conidia of C. africana (Texas isolate) obtained from newly formed honeydew in panicles of greenhouse-grown ATx623 were subsequently cultured in 150x10 mm Petri dishes containing 10 mL of the culture medium containing the different concentrations. Petri dishes containing water agar without floral extracts act as controls. A drop of conidial suspension was placed on four equidistant spots on the surface of the culture medium within each Petri dish. Petri dishes were subsequently incubated at 22±2°C for 24 h. Each Petri dish represented a replicate and each treatment was replicated three times. The number of germinated conidia (macroconidium producing a short sterigmata bearing a secondary conidium at the end) out of 100, counted within each of the 4 spots in the Petri dishes was recorded with the aid of a light microscope at 100X magnification.
Statistical Analysis
Stigma and anther/pollen extracts data were analyzed using the Analysis of Variance (GLM) procedure of SAS (Version 9.1, SAS Institute Inc., Cary, NC.) to determine the effect of cultivar, extract concentration, treatment (crushed or washed anther/pollen or stigma extracts and controls) and their interactions on ergot conidia germination. Due to heterogeneity of the error variances, data for the two experiments were analyzed separately. Percent stimulation or inhibition of C. africana conidia germination was determined by using this formula.
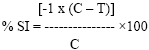 |
where % SI = stimulation or inhibition percentage, C = number of germinated conidia in water agar without floral extract and T = number of germinated conidia in water agar containing stigma, anther/pollen extract.
RESULTS
Anther/Pollen Extract
C. africana conidia germination was highly influenced by treatment (Trt), cultivar (Cul) and in the first experiment by concentration (Conc) of the extracts used (Table 1). The significant Cul xTrt and CulxConc interactions indicate that the amount of inhibition or stimulation of conidia germination on water agar amended with the extracts from the different cultivars was not the same at each treatment and concentration levels.
Figure 1 shows increases and reductions in conidia germination on water agar amended with the extracts over the water agar alone. Significant increases in conidia germination on plates containing crushed and washed anther/pollen extracts were noted for all cultivars, except Novartis KS310 in which plates containing washed extracts suppressed conidia germination in the first experiment.
| Table 1: | Analysis of variance for experiments on the in vitro assessments of anther/pollen and stigma extracts on Claviceps africana conidia germination |
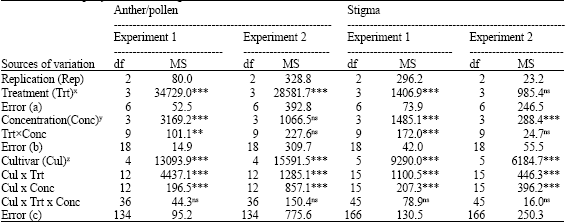 | |
| ns = not significant (p>0.05), ** = Significant (p≤0.05) and *** = Highly significant (p≤0.01), x Treatment = crushed, washed, crushed control and washed control, y Concentration = water agar amended at 0.5, 1, or 2% with either the anther/pollen or stigma extracts, z Cultivar = ATx99xRTx430, IS8525 (D), Novartis KS310, NC+7W97 and NC+8R18 were used for the anther/pollen experiments. These five cultivars and ATx623 were used for the stigma experiments | |
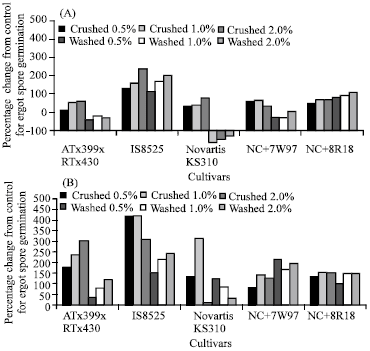 | |
| Fig. 1: | Percent increases or reductions in C. africana spore germination plated on water agar amended with three concentrations of extracts obtained from washed or crushed anther/pollen. Experiment 1 (A) and experiment 2 (B) |
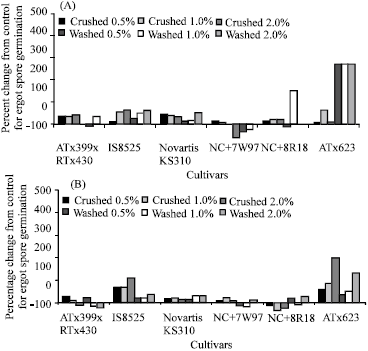 | |
| Fig. 2: | Percent increases or reductions in C. africana spore germination plated on water agar amended with three concentrations of extracts obtained from washed or crushed stigmas. Experiment 1 (A) and experiment 2 (B) |
Extracts from IS8525 stimulated the highest number of conidia germination. Generally, crushed anther/pollen extracts stimulated more conidia to germinate than washed extracts.
Stigma Extract
The main effect of Trt and TrtxConc interaction in the first experiment, Conc and Cul main effects, CulxTrt and CulxConc interactions in both experiments on conidia germination were highly significant (p<0.01) (Table 1). Percent increases or reductions in conidia germination due to stigma extracts are noted in Fig. 2. Water agar amended with stigma extracts from ATx623 and IS8525 stimulated more conidia to germinate than extracts from the other cultivars. Conidia germinations were suppressed on plates containing washed extracts from NC+7W97 and on plates containing crushed extracts from NC+8R18 in the second experiment.
DISCUSSION
Significant increases in C. africana conidia germination were observed when plated on water agar amended with sorghum anther/pollen or stigma extracts. Chinnadurai et al. (1970) noted marked increases in S. sorghi conidia germination when stigma extracts from both susceptible and resistant sorghum lines were added into slide cultures. Similarly, Bhuiyan et al. (2002) recorded higher germination rates of C. africana conidia on 1.5% water agar amended with extracts from boiled sorghum spikelets than on water agar alone.
In this study, higher rates of ergot conidia germinated on medium containing anther/pollen extracts than on medium containing stigma extracts. Leung (1992) documented high levels of chitinase activity in petal and stigma extracts of healthy petunia (Petuniax hybrida) flowers, whereas none was detected on the other floral structures including the stamen (anther and filament). Several studies have shown chitinase to inhibit both fungal spore germination and mycelial growth (Mauch et al., 1988; Darnetty et al., 1993; Seetharaman et al., 1997; Selitrennikoff, 2001). Although in this study the presence of chitinase in sorghum stigma extracts cannot be ascertained, there is the possibility that these extracts may contain elements with antifungal properties and these may have played a role in reducing the number of germinated conidia as compared to anther/pollen extracts.
In both experiments, the percentage of germinated conidia among the cultivars was not the same as indicated by the CulxTrt and CulxConc interactions (Table 1). In the stigma experiments, medium containing extracts from ATx623, the most ergot susceptible sorghum line stimulated higher rates of conidial germination than the other lines that are more tolerant to ergot infection ( Fig. 2). These results concur with that of Chinnadurai et al. (1970), which noted significantly higher rates of germinated ergot conidia in slide cultures amended with stigma exudates from susceptible sorghum lines than in cultures amended with exudates from resistant lines.
Secondary conidia of C. africana are the primary propagules for spreading the disease (Frederickson et al., 1993). This communication has demonstrated that crude extracts of anther/pollen or stigma extracts can increase the number of germinated conidia. Although, anther /pollen are not the sites of ergot infection, based on the results of this study these floral structures can act as substrate for C. africana growth and that the resulting secondary conidia can be dispersed and infect unfertilized ovaries. On the other hand, stigmas are the primary sites of infection for the sorghum ergot pathogens (Futrell and Webster, 1965; Bandyopadhyay et al., 1998). Currently, all types of sorghum, even among male-sterile plants show different levels of susceptibility to C. africana (Bandyopadhyay et al., 1998; Komolong et al., 2003). It is most likely that the stigma secretions may play a role in the different ergot reactions exhibited by the different sorghum lines. Further investigation of the chemical components of these extracts that are most inhibitory to ergot spore germination should be pursued. The elucidation of the mechanisms of action and appropriate concentrations required would be useful to more effectively control this important sorghum pathogen.
ACKNOWLEDGMENTS
I wish to thank Jonathan Egilla and Gurudev Mayalagu for their technical assistance.
REFERENCES
- Bandyopadhyay, R., D.E. Frederickson, N. McLaren, G.N. Odvody and M.J. Ryley, 1998. Ergot: A new disease threat to sorghum in the Americas and Australia. Plant Dis., 82: 356-367.
Direct Link - Bhuiyan, S.A., V.J. Galea, M.J., Ryley, D. Tay and A.T. Lisle, 2002. Factors influencing the germination of macroconidia and secondary conidia of Claviceps africana. Aust. J. Agric. Res., 53: 1087-1094.
Direct Link - Darnetty, J.F.L., J.F. Leslie, S. Muthukrishnan, M. Swegle, A.J. Vigers and C.P. Selitrennikoff, 1993. Variability in antifungal proteins in the grains of maize, sorghum and wheat. Physiol. Plant, 88: 339-349.
Direct Link - Frederickson, D.E., P.G. Mantle and W.A.J. de Milliano, 1993. Windborne spread of ergot disease (Claviceps africana) in sorghum A-lines in Zimbabwe. Plant Pathol., 42: 368--377.
Direct Link - Frederickson, D.E. and K. Leuschner, 1997. Potential use of benomyl for control of ergot (Claviceps africana) in sorghum A-lines in Zimbabwe. Plant Dis., 81: 761-765.
Direct Link - Isakeit, T., G.N. Odvody and R.A. Shelby, 1998. First report of sorghum ergot caused by Claviceps africana in the United States. Plant Dis., 82: 592-592.
Direct Link - Komolong, B., S. Chakraborty, M. Ryley and D. Yates, 2003. Ovary colonization by Claviceps africana is related to ergot resistance in male-sterile sorghum lines. Plant Pathol., 52: 620-627.
Direct Link - Leung, D.W.M., 1992. Involvement of plant chitinase in sexual reproduction of higher plants. Phytochemistry, 31: 1899-1900.
Direct Link - Mauch, F., B. Mauch-Mani and T. Boller, 1988. Antifungal hydrolases in pea tissue: II. Inhibition of fungal growth by combinations of chitinase and β-1,3-glucanase. Plant Physiol., 88: 936-942.
CrossRefPubMedDirect Link - Paoutova, S., R. Bandyopadhyay, D. Frederickson, P.G. Mantle and R.A. Frederiksen, 2000. Relations among sorghum ergot isolates from the Americas, Africa, India and Australia. Plant Dis., 84: 437-442.
Direct Link - Prom, L.K. and T. Isakeit, 2003. Laboratory, greenhouse and filed assessment of fourteen fungicides for activity against Claviceps africana, causal agent of sorghum ergot. Plant Dis., 87: 252-258.
Direct Link - Seetharaman, K., E. Whitehead, N.P. Keller, R.D. Waniska and L.W. Rooney, 1997. In vitro activity of sorgum seed antifungal proteins against grain mold pathogens. J. Agric. Food Chem., 45: 3666-3671.
Direct Link - Selitrennikoff, C.P., 2001. Antifungal proteins. Applied Environ. Microbiol., 67: 2883-2894.
CrossRefPubMedDirect Link - Tsukiboshi, T., T. Shimanuki and H. Koga, 2001. Claviceps sorghicola and C. africana, the ergot pathogens of sorghum and their cultural control in Japan. JARQ, 35: 221-226.
Direct Link