
Mohammad Ibrahim Al-Daej
College of Agriculture and Food Sciences, King Faisal University, P.O. Box 55065, 31982 Al-Ahsa, Kingdom of Saudi Arabia
LiveDNA: 966.24176
American Journal of Plant Physiology
Year: 2018 | Volume: 13 | Issue: 2 | Page No.: 58-64







ABSTRACT
Background and Objectives: Salinity stress is one of the main factors affecting the optimal yield of many vegetable crops considered as salt sensitive at early growth stages. The objective of this study was to study the salt tolerance of some tomato (Solanum lycoversicum L.) cultivars under laboratory conditions using different levels of NaCl. Materials and Methods: The experiment was carried out in vitro under laboratory conditions. Out of 10 varieties, only two cultivars namely “Rams” and “C10” were selected for investigation based on their physical characteristics. Both cultivars were subjected to 0, 20, 40, 60, 80 and 100 mM NaCl concentrations. The physiological characters such as seed germination, plant length, fresh weight, dry weight and number of leaves were studied against the salt stress. Also, the concentration of K, N, Na and Ca was determined in plant leaves. Results: The results showed that the cultivar Rams performed better than C10 for all the physical properties i.e., germination (%), fresh and dry matter yield etc. Also, the cultivar Rams accumulated less Na and K ions compared to cultivar C10. The cultivar Rams proved more salt tolerant even at high levels of salinity. Conclusion: Based on the studying findings, cultivar Rams proved more tolerant to salinity compared to other cultivars. Therefore, it can successfully be used in future breeding programs for its better salt tolerance capabilities to achieve optimal crop yield.
PDF Abstract XML References Citation
How to cite this article
Mohammad Ibrahim Al-Daej, 2018. Salt Tolerance of Some Tomato (Solanum lycoversicum L.) Cultivars for Salinity under Controlled Conditions. American Journal of Plant Physiology, 13: 58-64.
DOI: 10.3923/ajpp.2018.58.64
URL: https://scialert.net/abstract/?doi=ajpp.2018.58.64
DOI: 10.3923/ajpp.2018.58.64
URL: https://scialert.net/abstract/?doi=ajpp.2018.58.64
INTRODUCTION
High soil salinity coupled with heat stress are two of the most important constraints for crop production among various a biotic stresses1. Although the losses due to salinity are difficult to estimate, but according to a study around 950 Mha of surface land and 230 Mha of agricultural land is affected by salt2,3. Primarily, the high salt concentrations in the plant cells are due to osmotic and ionic stress combination4,5. Soil salinity is a global menace to plant growth and development causing significant economic losses to tomato and other crop plants6,7. Plants usually suffer from adverse effects due to osmotic stress during salinity. Because salinity causes a significant deficit in water uptake from soil that ultimately disturbs the osmotic balance in the affected plants8. Consequently, the water deficiency leads to secondary effects of salinity such as oxidative stress and ionic imbalance. Various plants develop diverse mechanisms to withstand high concentrations of salts in their surrounding environment9,10. The ability of plants to detect these ionic changes and their appropriate response to these changes is a pre-requisite for long-term survival under saline environment11. A counter-defense strategy is the accumulation of proline contents in their cells that helps in osmotic adjustment, scavenging free radicals and stabilizing sub-cellular structures12. Moreover, accumulation of proline also induces salt stress-responsive genes. The plants under high soil salinity stress can also synthesize various antioxidant enzymes to cope with oxidative stress13. Due to extreme aridity and continuous loss of water from soil surface, salts are accumulated in the upper soil layers that create water stress and imparts nutrient imbalances that trigger metabolic damage and cell death14. Salt stress triggers and provokes several signaling pathways to maintain ionic homeostasis and osmotic adjustment. Salt stress reduces plant growth and influence a series of physiological processes and finally suppress photosynthesis resulting in yield reduction4,1.
The crop production can be enhanced by improving the salt tolerance capacity of crop plants15. Micropropagation using tissue culture technology is very useful and efficient tool for mass production. Other modern biotechnological tools and contemporary approaches in molecular biology has made it entirely possible to study the expression of different genes implicated in stress tolerance of plant species. In plant tissue culture, different explants such as shoot apex, nodal segments, seeds and root segments are used to propagate tomato16.
Tomato (Solanum lycopersicum L.) is a vegetable crop grown for its high agro-economic value in the world. Tomato (family Solanacea) is one of the best crops to study salt tolerance in the dicotyledonous crops due to its well-known genetics and effortlessly transforming capabilities17. Tomato is moderately tolerant to salinity by regulating water and ionic homeostasis according to Martinez-Rodriguez et al.18. High salinity adversely affects seed germination and inhibit growth and fruit development in tomato19. In recent years, cultivation of tomato in Saudi Arabia increased from 310,401 ha in 1994 to 525,000 ha in 2012. However, it is still low when compared to the world average productivity because of exposure to salinity stress occurring under arid region conditions20 due to saline irrigation. The objective of this experiment was to study the salt tolerance of selected tomato cultivars under various levels of NaCl concentration in vitro. The experiment also delves explicitly on the effect of NaCl on these tomato cultivars concerning plant growth and its mineral composition. The purpose of this study was to alleviate the deleterious impact of salinity stress on the vegetative growth of specific tomato cultivars to improve growth and quality production of tomato crops in the country. Little research is available on these effects on the growth and yield of tomato crops.
MATERIALS AND METHODS
Preparation of plant material: This experiment was conducted at the Department of Biotechnology, King Faisal University (KFU), Saudi Arabia during the cropping season of 2017. The seeds from 10 local tomato cultivars (Super Marmande, Super strain B, Castle Rock, Sultana 7, Rams, C10, Liyan hybrid, Mahaly, Edkawy and Peto 86) were used to grow in vitro tomato plants (Table 1). The seeds were washed with autoclaved water for 15 min and sterilized by soaking in Clorox (Sodium hypochlorite) solution for 10 min, followed by thoroughly rinsing in tap water.
| Table 1: | Screening of tomato (Solanum lycoversicum L.) cultivars under different concentrations of NaCl |
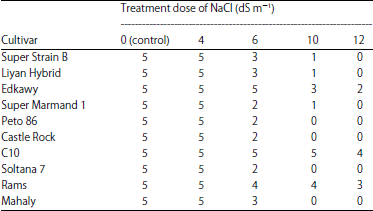 | |
| 0: No growth (plant dead), 1: Weak growth, 2-3: Moderate growth, 4: Good growth, 5: Very good growth | |
Then the seeds were immersed in 70% ethanol for 1 min and immersed in Clorox for 25 min followed by washing with sterilized distilled water. All the above-mentioned steps were carried out under laminar airflow cabinet.
Nutrient medium for seed germination development: The sterilized seeds were transferred to the Murashge and Sokge (MS)-media supplemented with 30 g L–1 sucrose, 40 mg L–1 adenine-sulfate, 200 mg L–1 glutamine, 100 mg L–1 myo-inositol, 0.1 mg L–1 biotin, 170 mg L–1 NaH2PO4, 0.1 mg L–1 thiamine HCl, 0.5 mg L–1 pyridoxine, 0.5 mg L–1 nicotinic acid, 3.0 mg L–1 2-isopentenyl adenine and 7.0 g L–1 agar. In all studied experiments, 30 g L–1 sucrose and BA at 1.0, 1.5, 2.0 mg L–1 with NAA at 0.5, 1.0, 1.5 mg L–1 were added to culture nutrient medium. The pH of the media was adjusted at 5.7 by 1M NaOH and the culturing jars were immediately capped and autoclaved at 121°C for 20 min. The seed explants were then incubated at 27±2°C and 16 h/daylight provided by white fluorescent tubes giving the intensity of about 2000 lux. After every three weeks interval, the explants were sub-cultured using the same media.
Culture conditions and NaCl concentration: After treatment with different concentrations of NaCl, the cultures were transferred to growth chamber for 25 days. The experiment was designed by following the complete randomized design (CRD) with three replicates for each treatment and 20 seeds per replicate were used for this experiment. There were six levels of NaCl concentration treatments ranging from 0, 20, 40, 60, 80 and 100 mM NaCl. These salt treatments were applied after seed germination.
Plant growth measurements: The physiological measurements included total fresh weight, total dry weight, seed germination, number of leaves and plant length. Moreover, the leaf Na, K, Ca and N were also determined.
Statistical analysis: The experimental data were subjected to two-way analysis of variance and the mean values were compared using least significant difference (LSD) test at the 5% level of probability using SPSS statistical software as given in SASs Institute, Inc.21. All the data were also statistically analyzed using factorial with Complete Randomized Design (CRD) using the same software. Lastly, the percentages were transformed to the arcsine to find the binomial percentage according to McDonald22.
RESULTS AND DISCUSSION
It was observed from the study results that the performance of all the treated germ plasm was similar when compared to control at 0 and 4 dS m–1 NaCl. However, the cultivar Rams and C10 performed better at higher NaCl concentrations than all the other cultivars (Table 1). Based on their performance under salt stress conditions, two cultivars “Rams” and “C10” were selected for further analysis. The germination (%) at 0, 20, 40, 60, 80 and 100 mM NaCl for cultivar Rams was 16, 14.5, 14.3, 12, 11.3 and 10% while it was 12, 8, 7.1, 4, 2 and zero percentage for C10, respectively. It is clear that the lowest concentration of NaCl showed little effect on the germination but with an increase in salt concentration the germination (%) of both tomato cultivars i.e., Rams and C10 was reduced (Fig. 1a). Regarding the fresh weight (Fig. 1b) and dry weights (Fig. 1c). The data for some leaves and plant length showed that the cultivar Rams out-classed the cultivar C10 with better performance (Fig. 1c, d). The cultivar Rams performed better under high salt levels than other cultivars. However, at the highest salt concentration, the cultivar C10 did not produce any plantlet growth compared to cultivar Rams (Fig. 1e). Previously, the salt tolerance ability of many plants was evaluated by in vitro tissue culturing technique23,24. Also, the effect of different growth regulators was also investigated for morphogenic tomato cultures25,26.
The accumulation of minerals such as Na, K, Ca and N was found in both the cultivars i.e., Rams and C10 (Fig. 2). Also, the cultivar Rams accumulated less Sodium at all the NaCl concentrations compared to cultivar C10 (Fig. 2b). Whereas, the accumulation of K element was less in cultivar Rams than the cultivar C10 (Fig. 2a). This was obvious that cultivar Rams accumulated less Na ion in its leaves and therefore; the accumulation of K was reduced (Fig. 2b). The data in Fig. 2c showed that the accumulation of Ca element was less in cultivar Rams when compared to cultivar C10 in some salt treatments. Similar, Fig. 2d showed that concentration of N was slightly was higher in cultivar Rams than cultivar C10 under different treatments.
This can also be explained by the high Na/K ratio of the cultivar Rams (Fig. 3a) compared to the cultivar C10 (Fig. 3b). The Na/Ca ratio showed decreasing trend with increasing in NaCl concentration in Rams cultivar (Fig. 3c), when compared to C10 cultivar (Fig. 3d). This indicated that cultivar Rams showed better salt tolerance capacity over other cultivars.
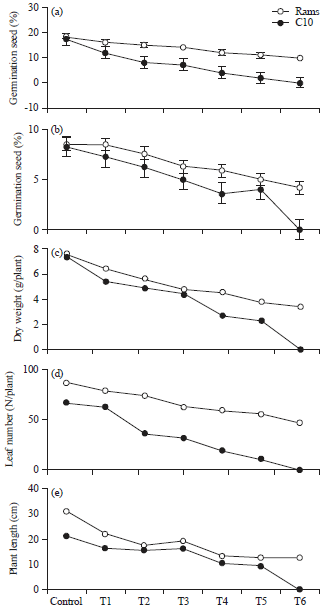 | |
| Fig. 1(a-e): | Effect of different levels of NaCl on (a) Seed germination, (b) Fresh weight, (c) Dry weight, (d) Number of leaves and (e) Plant growth of the tomato cultivars Rams and C10, respectively |
The results of this study agreed with the findings of Tuna et al.27, whereas, the seed germination was decreased with increasing the dose of NaCl. Salinity usually causes adverse effects on the seed germination and also suppresses plant growth and development in tomato cultivars19. Similar results were obtained in rice by Karlidag et al.28 and Horie et al.29 and strawberry by Keutgen and Pawelzik30 in terms of seed germination. In better salt adapted or salt tolerant plants, the Na/K and Na/Ca ratios are vital. Salt tolerant plants contain less dynamic Na nevertheless more Ca and K in their cells according to Chen et al.31 and Flowers et al.4. The salinity stress applied through Na salts not only reduces Ca availability but also reduces the mobility of Ca into the growing parts in the plant.
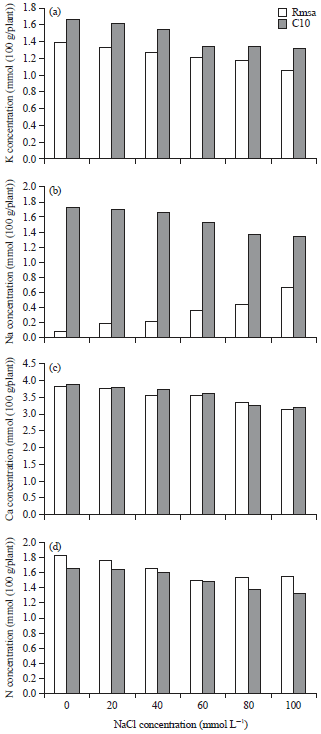 | |
| Fig. 2(a-d): | Effect of different levels of NaCl on (a) K, (b) Na, (c) Ca and (D) N in the tomato cultivars Rams and C10, respectively |
Moreover, the high salinity induced N deficiency in tomato, Cabbage and Lettuce as reported by Yokas et al.32. High levels of Na concentration in the plants increases the osmotic potential thus causing low or decreased water uptake. Otherwise, the concentration of K plays an important role in the plant since it shows little changes under salinity stress. Thus, the increase of K in the plant means it can be used by breeding programmers in future studies.
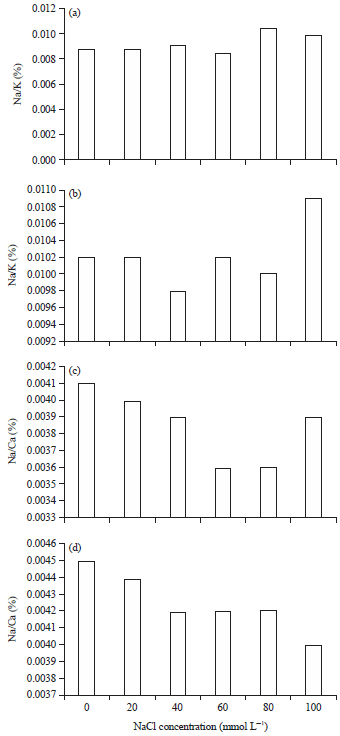 | |
| Fig. 3(a-d): | Different levels of (a-b) Na/K and (c-d) Na/Ca ratios in the leaves of two tomato A, C: Cultivars Rams and B, D: C10, respectively |
Also the study results agree with the findings of many researchers namely Aazami et al.33, who found significant differences among cultivars regarding the highest callus relative growth (RGR), while the cultivar Ram showed the lowest RGR under salt levels. Any increase in salinity levels in the media led to decrease of RGR and in contrast increased the dry matter (DM). Also Sardoei and Mohammadi34 and Raza et al.35 observed that increased salt level results in reduction of plumule fresh weight indices of flat ch irani, cal ji and chef flat amrica cultivar; however the fresh weight of plumule was not significantly different between cal ji and flat ch irani.
CONCLUSION
The results showed that the response to salinity differs significantly among various tomato cultivars. However, this study provided useful information on the salt tolerance mechanism for in vitro screening of tomato plants under high salt stress conditions. The cultivar Rams proved to be the best suited for salt stress conditions. Furthermore, field trials of this cultivar would be an important step for the selection and final validation of its performance under field conditions. In conclusion, the study findings under high salt stress conditions would be useful for the plant breeders to select and choose this cultivar in ongoing tomato breeding programs for sustainable tomato production.
SIGNIFICANCE STATEMENT
This study discovered highly detrimental effects of high salinity on different growth parameters of various tomato cultivars. The study findings under high salt stress conditions are useful and will help the plant breeders to select and choose an appropriate tomato cultivar in the ongoing crop breeding programs for sustainable tomato production.
ACKNOWLEDGMENTS
This project was funded by the Deanship of Scientific Research, King Faisal University (KFU), Saudi Arabia with a Grant No. 140246. I would like to thank Dr. Abdel Rezk, Department of Biotechnology, King Faisal University for his help and valuable suggestions. Also great appreciation to Dr. Hesham Ali, Date Palm Research Center, King Faisal University for his help in some laboratory work.
REFERENCES
- Zhang, L., L. Song, H. Shao, C. Shao and M. Li et al., 2014. Spatio-temporal variation of rhizosphere soil microbial abundance and enzyme activities under different vegetation types in the coastal zone, Shandong, China. Plant Biosyst. Int. J. Dealing Aspects Plant Biol., 148: 403-409.
CrossRefDirect Link - Ruan, C.J., J.A.T. da Silva, S. Mopper, P. Qin and S. Lutts, 2010. Halophyte improvement for a salinized world. Crit. Rev. Plant Sci., 29: 329-359.
CrossRefDirect Link - Lutts, S., J.M. Kinet and J. Bouharmont, 1996. NaCl-induced senescence in leaves of rice (Oryza sativa L.) cultivars differing in salinity resistance. Ann. Bot. 78: 389-398.
CrossRefDirect Link - Flowers, T.J., R. Munns and T.D. Colmer, 2014. Sodium chloride toxicity and the cellular basis of salt tolerance in halophytes. Ann. Bot., 115: 419-431.
CrossRefDirect Link - Song, J. and B. Wang, 2014. Using euhalophytes to understand salt tolerance and to develop saline agriculture: Suaeda salsa as a promising model. Ann. Bot., 115: 541-553.
CrossRefDirect Link - Kalaji, H.M., A. Jajoo, A. Oukarroum, M. Brestic and M. Zivcak et al., 2016. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant., Vol. 38.
CrossRefDirect Link - Albaladejo, I., V. Meco, F. Plasencia, F.B. Flores, M.C. Bolarin and I. Egea, 2017. Unravelling the strategies used by the wild tomato species Solanum pennellii to confront salt stress: From leaf anatomical adaptations to molecular responses. Environ. Exp. Bot., 135: 1-12.
CrossRefDirect Link - Munns, R. and M. Tester, 2008. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol., 59: 651-681.
CrossRefDirect Link - Chen, T.H.H. and N. Murata, 2002. Enhancement of tolerance of abiotic stress by metabolic engineering of betaines and other compatible solutes. Curr. Opin. Plant Biol., 5: 250-257.
CrossRefPubMedDirect Link - Munns, R. and M. Gilliham, 2015. Salinity tolerance of crops-what is the cost? New Phytol., 208: 668-673.
CrossRefDirect Link - Deinlein, U., A.B. Stephan, T. Horie, W. Luo, G. Xu and J.I. Schroeder, 2014. Plant salt-tolerance mechanisms. Trends Plant Sci., 19: 371-379.
CrossRefDirect Link - Hayat, S., Q. Hayat, M.N. Alyemeni, A.S. Wani, J. Pichtel and A. Ahmad, 2012. Role of proline under changing environments: A review. Plant Signal. Behav., 7: 1456-1466.
CrossRefDirect Link - Seckin, B., A.H. Sekmen and I. Turkan, 2009. An enhancing effect of exogenous mannitol on the antioxidant enzyme activities in roots of wheat under salt stress. J. Plant Growth Regul., 28: 12-20.
CrossRefDirect Link - Hasanuzzaman, M., K. Nahar and M. Fujita, 2013. Plant Response to Salt Stress and Role of Exogenous Protectants to Mitigate Salt-Induced Damages. In: Ecophysiology and Responses of Plants under Salt Stress, Ahmad, P., M.M. Azooz and M.N.V. Prasad (Eds.). Chapter 2, Springer, New York, USA., ISBN-13: 9781461447467, pp: 25-87.
- Karan, R. and P.K. Subudhi, 2012. A stress inducible SUMO conjugating enzyme gene (SaSce9) from a grass halophyte Spartina alterniflora enhances salinity and drought stress tolerance in Arabidopsis. BMC Plant Biol., Vol. 12, No. 1.
CrossRefDirect Link - Yin, W., Z. Hu, J. Hu, Z. Zhu, X. Yu, B. Cui and G. Chen, 2017. Tomato (Solanum lycopersicum) MADS-box transcription factor SlMBP8 regulates drought, salt tolerance and stress-related genes. Plant Growth Regul., 83: 55-68.
CrossRefDirect Link - Martinez-Rodriguez, M.M., M.T. Estan, E. Moyano, J.O. Garcia-Abellan and F.B. Flores et al., 2008. The effectiveness of grafting to improve salt tolerance in tomato when an excluder genotype is used as scion. Environ. Exp. Bot., 63: 392-401.
CrossRefDirect Link - Cuartero, J., M.C. Bolarin, M.J. Asins and V. Moreno, 2006. Increasing salt tolerance in the tomato. J. Exp. Bot., 57: 1045-1058.
CrossRefDirect Link - Feleafel, M.N. and Z.M. Mirdad, 2014. Alleviating the deleterious effects of water salinity on greenhouse grown tomato. Int. J. Agric. Biol., 16: 49-56.
Direct Link - McDonald, J.H., 2014. Handbook of Biological Statistics. 3rd Edn., Sparky House Publishing, Baltimore, Maryland, USA., Pages: 305.
Direct Link - Amini, F. and A.A. Ehsanpour, 2006. Response of tomato (Lycopersicon esculentum Mill.) cultivars to MS, water agar and salt stress in in vitro culture. Pak. J. Biol. Sci., 9: 170-175.
CrossRefDirect Link - Bhatia, P., N. Ashwanth, T. Senaratna and D. Midmore, 2004. Tissue culture studies of tomato (Lycopersicon esculentum). Plant Cell. Tissue Organ Cult., 78: 1-21.
CrossRefDirect Link - Branca, C., A. Torelli, P. Fermi, M.M. Altamura and M. Bassi, 1994. Early phases in in vitro culture of tomato cotyledons: Starch accumulation and protein pattern in relation to the hormonal treatment. Protoplasma, 182: 59-64.
CrossRefDirect Link - Liza, L.N., A.N.M. Nasar, K.M.A. Zinnah, M.A.N. Chowdhury and M. Ashrafuzzaman, 2013. In vitro growth media effect for regeneration of tomato (Lycopersicon esculentum) and evaluation of the salt tolerance activity of callus. J. Agric. Sustainab., 3: 132-143.
Direct Link - Tuna, A.L., B. Burun, I. Yokas and E. Coban, 2002. The effects of heavy metals on pollen germination and pollen tube length in the tobacco plant. Turk. J. Biol., 26: 109-113.
Direct Link - Karlidag, H., E. Yildirim and T. Metin, 2009. Salicylic acid ameliorates the adverse effect of salt stress on strawberry. Sci. Agric., 66: 180-187.
Direct Link - Horie, T., I. Karahara and M. Katsuhara, 2012. Salinity tolerance mechanisms in glycophytes: An overview with the central focus on rice plants. Rice, Vol. 5.
CrossRefDirect Link - Keutgen, A.J. and E. Pawelzik, 2009. Impacts of NaCl stress on plant growth and mineral nutrient assimilation in two cultivars of strawberry. Environ. Exp. Bot., 65: 170-176.
CrossRefDirect Link - Chen, Z., I. Newman, M. Zhou, N. Mendham, G. Zhang and S. Shabala, 2005. Screening plants for salt tolerance by measuring K+ flux: A case study for barley. Plant Cell Environ., 28: 1230-1264.
Direct Link - Yokas, I., A.L. Tuna, B. Burun, H. Altunlu, F. Altana and C. Kaya, 2008. Responses of the tomato (Lycopersicon esculentum Mill.) plant to exposure to different salt forms and rates. Turk. J. Agric. For., 32: 319-329.
Direct Link - Aazami, M.A., M. Torabi and F. Shekari, 2010. Response of some tomato cultivars to sodium chloride stress under in vitro culture condition. Afr. J. Agric. Res., 5: 2589-2592.
Direct Link - Sardoei, A.S. and G.A. Mohammadi, 2014. Study of salinity effect on germination of tomato (Lycopersicon esculentum L.) genotypes. Eur. J. Exp. Bio., 4: 283-287.
Direct Link - Raza, M.A., A. Saeed, H. Munir, K. Ziaf and A. Shakeel et al., 2017. Screening of tomato genotypes for salinity tolerance based on early growth attributes and leaf inorganic osmolytes. Arch. Agron. Soil Sci., 63: 501-512.
CrossRefDirect Link