
Sardar A. Farooq
Department of Biology, Sultan Qaboos University, P.O. Box 36, Al Khod, PC123, Oman
LiveDNA: 91.24000
Talat T. Farook
Department of Biology, Sultan Qaboos University, P.O. Box 36, Al Khod, PC123, Oman
Salim H. Al Rawahy
ORIGO Group, P.O. Box 1236, Muscat, PC133, Oman
American Journal of Plant Physiology
Year: 2018 | Volume: 13 | Issue: 2 | Page No.: 53-57







ABSTRACT
Background and Objective: Clerodendrum inerme (L.) Gaertn. is commonly grown as a hedge plant in tropical countries, belongs to dicotyledon family Verbenaceae. It is used in the herbal medicine and has several useful phytochemicals, which can be exploited commercially without destruction of natural populations. To achieve large-scale micropropagation of plants, a rapid reproducible protocol is reported. Materials and Methods: Nodal and leaf explants were surface sterilized with 0.1% HgCl2 and plated on woody plant medium with three different concentrations of each auxin (2, 4-D and IBA); cytokinins (BAP and 2-ip) and a combination of these two. One way ANOVA is used and means are compared by Duncan’s multiple range test (p<0.01). Results: Adventitious (direct) organogenesis produced multiple shoots from nodal explants in media containing BAP while leaf explants yielded callus in IBA and 2, 4-D media. Denovo (indirect) differentiation of the callus produced somatic embryoids when transferred to cytokinin (BAP) media. Shoots were successfully rooted on PGR free media with reduced sucrose. About 25% mortality occurred in regenerated plants during acclimatization process. Conclusion: An efficient, rapid and reproducible plantlet regeneration protocol using nodal and leaf explants is reported for Clerodendrum inerme. Adventitious (direct) organogenesis from nodal explants produced multiple shoot buds within 4 weeks while denovo (indirect) somatic embryogenesis through callus is prevalent in leaf explants.
PDF Abstract XML References Citation
How to cite this article
Sardar A. Farooq, Talat T. Farook and Salim H. Al Rawahy, 2018. Adventitious and Denovo Somatic Embryogenesis from Nodal and Leaf Explants of Clerodendrum inerme. American Journal of Plant Physiology, 13: 53-57.
DOI: 10.3923/ajpp.2018.53.57
URL: https://scialert.net/abstract/?doi=ajpp.2018.53.57
DOI: 10.3923/ajpp.2018.53.57
URL: https://scialert.net/abstract/?doi=ajpp.2018.53.57
INTRODUCTION
Multiple medicinal uses of plants resulted in large scale cutting down of plants from the local Flora, lead to the depletion of plant species making them endangered and extinct ultimately. Such over-exploitation of natural resources calls for conservation of plants. However, multiplication of wild plants is not easy task because very little is known about their reproductive seed biology, while seed propagation is limited by their inherent low seed viability and germination potential1,2. In light of these constraints, biotechnology applications such as plant tissue culture play an important role in the mass propagation in ex situ conservation and increase in sustainable productivity of native plants that can be pivotal in the rehabilitation of desert in Arabian peninsula1. The tissue culture system allows the propagation of selected genotypes with high multiplication rates in an aseptic, temperature-controlled environment and the miniaturization of explants allows the reduction of space requirements and would prove to be economical in the long run2-5. The genus Clerodendrum has 8 species of which Clerodendrum inerme (L.) Gaertn is an important medicinal plant distributed in tropical Asia and middle east in various habitats and is a salt tolerant species grown as a hedge plant in poor or marginal soils. The plant known to have mosquitocidal, nematicidal, antihepatotoxic, antipyretic properties. In folklore medicine used in treatment of Elephantiasis, Dengue fever and in Malaria as a substitute for quinine6 and also used in rheumatism, skin disease, venereal infections, beriberi and tumors, cough, skin rashes, boils and fever7. Clerodendrum inerme is a cross pollinated species and propagated by seeds, hence show lot of variation. Clerodendrum has several useful phytochemicals such as inerminosides, iridoid glycosides, phenylpropanoids and antiviral peptides. Of late the commercial establishments ruthlessly cut down this plant for extraction of these chemicals in large scale. Consequently, natural populations of Clerodendrum are dwindling at a faster pace, paving the need for ex situ conservation of this species In the recent past, plant tissue culture is extensively used in the process of ex situ conservation. In Clerodendrum plant tissue culture and micropropagation was attempted earlier with limited success8-12. However, there is inconsistency in the published protocols and their reproducibility. Therefore the present studies undertaken to evolve an efficient, rapid, reproducible protocol for mass propagation of C. inerme from nodal and leaf explants. Successful regeneration of plants in vitro depends on the genotype, physiological status and growth conditions of the donor plants. The process of organogenesis or somatic embryogenesis is classified into two categories viz., Adventitious (direct) and denovo (indirect). In the present study, the former confined to the nodal explants while the latter was predominant in leaf explants.
MATERIALS AND METHODS
Chemicals used: All chemicals used in this study were obtained from Sigma-Aldrich (Plant cell culture tested, Bio Reagents).
Explant collection: Young leaves and nodal segments were collected from healthy Clerodendrum inerme plants growing in the Botanical garden of Sultan Qaboos University, Al Khod, Oman, in November, 2015.
Surface sterilization: Leaf and Nodal segments were washed thoroughly for 15 min with a detergent solution mixed with 2-3 drops of tween 20, followed by soaking in Mercuric Chloride HgCl2 0.1% for 10 min and rinsed thrice with sterile water. The leaf was cut into 1.5 cm square pieces with cut margins on all four sides. In case of nodes, 2.5 cm segments with axillary buds were cut and inoculated aseptically media.
Media: Woody plant medium (WPM) of Lloyd and McCown13 containing 3% sucrose, 8% agar, vitamins and plant growth regulators (PGR) was used. Different concentrations of auxin (IBA or 2, 4-D) and the cytokinins (BAP or 2-ip) were added to media (Table 1). Plant growth regulator concentrations were different in callusing or shoot induction or rooting media. The pH of the medium was adjusted to 5.6±0.2. Ventral side of the leaf was kept in contact with the media, while the nodal explants were inserted slantly at 45° in the media.
Culture conditions: The cultures were incubated at 25±2° C with 16 h. light regime (1000 lux from cool fluorescent tubes) in Binder KBW 720 Plant Growth Chamber. Subculture was done after every 4 weeks.
Rooting: When the shoots attained about 1.5 cm size were transferred to rooting medium containing reduced WPM salts (0.5x) and 1.5% sucrose. The rooted plantlets transferred to autoclaved vermiculite and covered to maintain 70-80% humidity to acclimatize before transfer to green house.
Statistical analysis: At least three replicates were maintained for each PGR treatment. The data were recorded from the cultures regularly and statistically analyzed. The SPSS 19 was used to analyze Mean, Standard Deviation and one way Analysis of variance (ANOVA). Comparison of Means was done by Duncan’s multiple range test (p<0.01).
| Table 1: | Effect of different concentrations of plant growth regulators on shoot/root induction in Clerodendrum inerme |
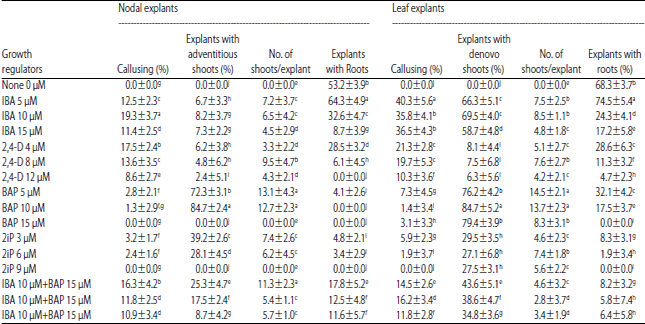 | |
| Data presented as Mean±Standard deviation with different letters within a column indicate significant differences at p<0.01 according to Duncan’s multiple range test | |
RESULTS AND DISCUSSION
The leaf has mature, quiescent cells which dedifferentiated and returned to meristematic activity due to exogenous auxins (IBA or 2, 4-D) that caused swelling of leaf explants after 7 days of incubation in the media. White crystalline callus was noticed along the cut margins of leaf segments in 15 days old cultures. There was very little callus from nodal explants in IBA media, instead the elongation of axillary buds occurred in media with BAP. Multiple shoot buds formed adventitiously from axillary buds of nodal explants. Sprouting of axillary buds into adventitious shoots was more pronounced in media with BAP from nodal explants (Fig. 1), whereas no shoot buds formed in IBA and 2, 4-D media. This indicated that nodal explants have endogenous auxin that suppressed development of axillary buds due to apical dominance whereas, BAP nullified this phenomenon and allowed lateral buds to form multiple shoots. These results are in agreement with the properties of auxin and its action14. It is interesting to note that adventitious (direct) organogenesis occurred only in nodal explants whereas callus mediated denovo (indirect) somatic embryogenesis or organogenesis occurred in leaf explants. This suggested that nodal explants have PEDC (Pre Embryonic Determined Cells) and the PGRs in the media trigger these cells to resort to direct somatic embryogenesis15,16. Proliferation of shoot buds from nodal explants in C. inerme was reported earlier9-11 with 8.8-16 μM BA. This study incorporates a comparative account of 3 levels of two auxins (IBA and 2, 4-D) and two cytokinins (BAP and 2-ip) in induction of somatic embryogenesis from nodal and leaf explants (Table 1). Adventitious embryos also formed from the cut ends in the leaf explant at relatively lower frequency. It is presumed that the injury caused to leaf explant due to cut margins exposed the cells and activated SERK gene (Somatic Embryogenesis Receptor-like Kinase) to express at relatively higher level in the presence of BAP and allowed formation of somatic embryos17,18. It can be noticed that there is no relationship between the concentration of PGR in the media and the embryogenic or callogenic response because at higher concentration of IBA or BAP or 2, 4-D or 2- ip there is reduced response. This indicates that somatic embryogenesis does not entirely depend on the absolute amount of PGRs in the media, instead it may depend on the endogenous A: C (auxin to cytokinin) ratio. It is possible that such response might depend more on threshold level of endogenous and exogenous PGRs. Similar opinion was reported in Triticale embryogenesis19 where genotype-specific threshold level of PGR was required and higher endogenous accumulation diminished the effectiveness of the process.
The callus growth was rapid in leaf explants in 10 μM IBA while slow in 4 μM 2, 4-D media (Table 1). The auxin initiates the dedifferentiation of mature, quiescent cells in leaf explant and induces repeated divisions in them resulting in callus. This process is optimized by varying the auxin to cytokinin ratio in the medium20. In these experiments IBA was more efficient in callusing than 2, 4- D, contradicting earlier reports favoring 2, 4-D in certain plant species. Besides callus induction and shoot-bud regeneration frequency varies with genotype, explant, and plant-growth regulators21,22. Even though callus is amorphous, the texture was different in different culture media.
 | |
| Fig. 1(a-e): | (a) Clerodendrum plant, (b) Callus from leaf explants, (c, d) Somatic embryoids from leaf callus and (e) Multiple shoots from nodal explant |
Soft callus was common in 2, 4-D and 2-ip supplemented media while compact, friable callus was noticed in IBA media, the latter has better organogenic potential. Presence of BAP and 2-ip triggered callus cells to differentiate into embryogenic competent cells.
Denovo somatic embryogenesis induced when the callus was transferred to IBA+BAP medium. The development started with globular embryoids which developed into heart shaped and torpedo stages. Even though, the embryoids were bipolar, there was more shoot growth, while root formation was slow or rudiment, hinting at unipolar organogenetic pathway. Root growth progressed at faster pace when BAP was withdrawn from the media. Irrespective of the origin of calli (from IBA or 2, 4-D) the ability to form shoot buds did not differ much. The green shoots were noticed in all the media used. The highest frequency of shoot regeneration was recorded in media with 10 μM IBA+10 μM BAP and the lowest frequency was obtained with 12 μM 2, 4-D. The combination of BAP+IBA enhanced the somatic embryogenesis (Table 1). Somatic embryoids were also induced from callus in 2-ip media, however at a lower frequency. In Clerodendrum 2-iP is found to be less potent in inducing shoots on contrary to other reports in some plant species23-25. At higher concentration of BAP and 2- ip the somatic embryogenesis was reduced. This may be due to the high concentration of IBA may disturb the balance between A:C ratio and this may be a factor in reducing shoot buds. These results find support from Muller and Sheen26. Transfer of callus to BAP media did not improve callogenesis, but induced somatic embryogenesis. These observations are in conformity with the results obtained in Acacia and date palm27.
Rhizogenesis of the shoot buds was successfully achieved on reduced sucrose (1.5%) coupled with reduced WPM salts (0.5x). Low IBA was more effective in rooting. Never the less, hormone free WPM (0.5x) with 1.5% sucrose was equally effective in inducing roots (Table 1). The BAP and 2- ip were least effective. Acclimatization of regenerated plants was a critical step where 75-80% survival recorded. To minimize mortality, the regenerated plants were transferred to autoclaved vermiculite and covered with transparent plastic cups and sprayed with distilled water to maintain high humidity to prevent from desiccation. The acclimatized plants transferred to the field and grown to maturity. There were no observable phenotypic variations in regenerated plants. It can be noticed that the protocol used in this study for mass propagation is superior to earlier reports in C. inerme 9-11.
CONCLUSION
It is concluded that reproducible protocol for adventitious shoot buds from nodal explants and denovo somatic embryogenesis from leaf callus. IBA is more efficient in inducing callus than 2,4-D whereas BAP is superior over 2-ip for somatic embryogenesis. Profuse root formation occurs in media having 0.5x WPM salts with 1.5% sucrose 75% regenerated plants survived after acclimatization.
SIGNIFICANCE STATEMENT
Present studies resulted an efficient, rapid and reproducible plantlet regeneration protocol using nodal and leaf explants for micropropagation of Clerodendrum inerme. Adventitious (direct) organogenesis obtained from nodal explants within 4 weeks, while the leaf explants produced copious amounts of callus that later induced denovo somatic embryogenesis. This study recommends woody plant media with specific concentration of BAP and IBA is for rapid multiplication of Clerodendrum inerme.
ACKNOWLEDGMENTS
I would like to thank Sultan Qaboos University, Oman which provided facilities for this study.
REFERENCES
- Sudhersan, C., M. Abo El-Nil and J. Hussain, 2003. Tissue culture technology for the conservation and propagation of certain native plants. J. Arid Environ., 54: 133-147.
CrossRefDirect Link - Mounce, R., P. Smith and S. Brockington, 2017. Ex situ conservation of plant diversity in the world’s botanic gardens. Nat. Plants, 3: 795-802.
CrossRefDirect Link - Raven, P. and K. Havens, 2014. Ex situ plant conservation and cryopreservation: Breakthroughs in tropical plant conservation. Int. J. Plant Sci., 175: 1-2.
CrossRefDirect Link - Canter, P.H., H. Thomas and E. Ernst, 2005. Bringing medicinal plants into cultivation: Opportunities and challenges for biotechnology. Trends Biotechnol., 23: 180-185.
CrossRefDirect Link - Chethana, G.S., K.R.H. Venkatesh and S.M. Gopinath, 2013. Review on Clerodendrum inerme. J. Pharm. Scient. Innov., 2: 38-40.
Direct Link - Gupta, S.P., S. Somkuwar and G. Garg, 2010. Clerodendron inerme: An update of its indigenous uses, phytochemistry and pharmacology. Int. J. Chem. Sci., 8: 203-212.
Direct Link - Baburaj, S., P. Ravichandran and M. Selvapandian, 2000. In vitro adventitious shoot formation from leaf cultures of Clerodendrum inerme (L.) Gaertn. Indian J. Exp. Biol., 38: 1274-1276.
Direct Link - Avani, K., P. Harish and S. Neeta, 2006. Ex situ conservation method for Clerodendrum inerme: A medicinal plant of India. Afr. J. Biotechnol., 5: 415-418.
Direct Link - Nataraj, M., M.M. Kher and J.A.T. da Silva, 2016. Micropropagation of Clerodendrum L. species: A review. Rendiconti Lincei, 27: 169-179.
CrossRefDirect Link - Lloyd, G. and B. McCown, 1980. Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot-tip culture. Proc. Int. Plant Propagator's Soc., 30: 421-426.
Direct Link - Sharp, W.R., M.R. Sondahl, L.S. Caldas and S.B. Maraffa, 1980. The physiology of in vitro asexual embryogenesis. Hortic. Rev., 2: 268-310.
CrossRefDirect Link - Rengel, Z., 1987. Factors involved in initiation of somatic embryogenesis in cereal tissue culture. Acta Bot. Croat., 46: 33-43.
Direct Link - Lu, D., W. Wei, W. Zhou, L.D. McGuigan and F.Y. Ji et al., 2017. Establishment of a somatic embryo regeneration system and expression analysis of somatic embryogenesis-related genes in Chinese chestnut (Castanea mollissima Blume). Plant Cell Tissue Organ Cult., 130: 601-616.
CrossRefDirect Link - Zur, I., E. Dubas, M. Krzewska, P. Waligorski, M. Dziurka and F. Janowiak, 2015. Hormonal requirements for effective induction of microspore embryogenesis in triticale (× Triticosecale Wittm.) anther cultures. Plant Cell Rep., 34: 47-62.
CrossRefDirect Link - Pasternak, T., P. Miskolczi, F. Ayaydin, T. Meszaros, D. Dudits and A. Feher, 2000. Exogenous auxin and cytokinin dependent activation of CDKs and cell division in leaf protoplast-derived cells of alfalfa. Plant Growth Regul., 32: 129-141.
CrossRefDirect Link - Venkatachalam, P. and N. Jayabalan, 1997. Effect of auxins and cytokinins on efficient plant regeneration and multiple-shoot formation from cotyledons and cotyledonary-node expiants of groundnut (Arachis hypogaea L.) by in vitro culture technology. Appl. Biochem. Biotechnol., 67: 237-247.
CrossRefDirect Link - Wahyuni, D.K., A.N.M. Ansori and F. Vidiyant, 2014. GC-MS analysis of the leaf-derived callus of Justicia gendarussa Burm.f. Biosci. Res., 14: 668-677.
Direct Link - D'Souza, M.C. and M. Sharon, 2001. In vitro clonal propagation of annatto (Bixa oreliana L.). In Vitro Cell. Dev. Biol.-Plant, 37: 168-172.
CrossRefDirect Link - Jana, S., I. Sivanesan and B.R. Jeong, 2013. Effect of cytokinins on in vitro multiplication of Sophora tonkinensis. Asian Pac. J. Trop. Biomed., 3: 549-553.
CrossRefPubMedDirect Link - Chen, R., M. Zhang, J. Lu, X. Zhang, J.A.T. da Silva and G. Ma, 2014. Shoot organogenesis and somatic embryogenesis from leaf explants of Valeriana jatamansi Jones. Sci. Hortic., 165: 392-397.
CrossRefDirect Link - Muller, B. and J. Sheen, 2008. Cytokinin and auxin interaction in root stem-cell specification during early embryogenesis. Nature, 453: 1094-1097.
CrossRefDirect Link - Sane, D., A. Borgel, J.L. Verdeil and Y.K. Gassama, 2000. Plantlet regeneration via somatic embryogenesis in immature zygotic embryo callus from a tree species adapted to arid lands: Acacia tortilis subsp. raddiana (Savi.) Brenan. Acta Bot. Gallica, 147: 257-266.
Direct Link