
Sonam Sneha
Department of Biotechnology and Allied Sciences, Jayoti Vidyapeeth Women�s University, Jaipur, 303122, Rajasthan, India
Anirudha Rishi
Department of Biotechnology and Allied Sciences, Jayoti Vidyapeeth Women�s University, Jaipur, 303122, Rajasthan, India
Subhash Chandra
Department of Biotechnology and Allied Sciences, Jayoti Vidyapeeth Women�s University, Jaipur, 303122, Rajasthan, India
American Journal of Plant Physiology
Year: 2014 | Volume: 9 | Issue: 1 | Page No.: 32-37







ABSTRACT
In this study the 21 days old seedlings of Pennisetum glaucum were subjected to short term salt stress in order to observe the effect on chlorophyll content, protein and antioxidative enzyme activity (CAT and APX) responses. The aim of the experiment was to evaluate the changes in chlorophyll content and antioxidant enzymes (Catalase and Ascorbate peroxidase). The 21 days old seedlings were subjected to salt stress by supplementing Hoagland’s solution with different concentrations of Sodium chloride (50, 100, 150 and 200 mM). Measurement of chlorophyll content and antioxidant enzyme activity were taken at different time intervals of salt treatment (12, 24, 48, 72, 96 and 120 h). The results showed statistically significant differences in traits for the salt treatment. A significant increase in Catalase activity was observed under all the salt concentrations while no significant activity of Ascorbate peroxidase activity was observed. The protein content and chlorophyll content decreased with increasing salt concentration. This result shows salt stress affects the photosynthesis rate by decreasing chlorophyll content. Catalase enzyme plays an important role in scavenging reactive oxygen species generated due to salt stress in the plant cell.
PDF Abstract XML References Citation
Received: October 09, 2013;
Accepted: November 13, 2013;
Published: March 28, 2014
How to cite this article
Sonam Sneha, Anirudha Rishi and Subhash Chandra, 2014. Effect of Short Term Salt Stress on Chlorophyll Content, Protein and Activities
of Catalase and Ascorbate Peroxidase Enzymes in Pearl Millet. American Journal of Plant Physiology, 9: 32-37.
DOI: 10.3923/ajpp.2014.32.37
URL: https://scialert.net/abstract/?doi=ajpp.2014.32.37
DOI: 10.3923/ajpp.2014.32.37
URL: https://scialert.net/abstract/?doi=ajpp.2014.32.37
INTRODUCTION
Pearl millet (Pennisetum glaucum (L.) R. Br.) is an important crop of the semi-arid tropics in Africa and India. Pearl millet is the fifth most important cereal crop and the most important millet with more than 55% of global millet production. In India pearl millet occupies fourth position among cereal crops next to rice, wheat and sorghum. Pearl millet is mostly grown in the area inflicted with salinity and drought. Both salinity and drought affects the morphological characteristics as well as the metabolic activity. Salinity causes a range of injurious effect such as inhibition of photosynthesis rate by decreasing chlorophyll content, damage to plasma membrane permeability and other metabolic commotions (Karimi et al., 2005; Ashraf and Praveen, 2002). Changes in activities of various antioxidant enzymes under salinity stress have been reported (Venkatesan and Sridevi 2009; Koskeroglu and Levent, 2008; Hernandez et al., 2000). Stress tolerance/adaptation seems to be correlated with spur of antioxidant enzymes and the enhanced ability to remove ROS and a higher concentration of CAT is noted (Santos et al., 2001; Bettaieb et al., 2007; Xu and Huang, 2004). CATs and SOD are principle enzymes which scavenge active oxygen species and reduce the protein degradation, lipid peroxidation, cell membranes damage and chlorophyll degradation (Foyer et al., 1994; Smirnoff, 1993). Catalase and ascorbate peroxidase acts as the main defense against ROS produced in various parts of plant cells (Apel and Hirt, 2004).
Catalase (EC 1.11.1.6; CAT) is a tetrameric hemecontaining enzyme that is found in all aerobic organisms and serve to rapidly degrade H2O2 at an extremely rapid rate. The expression of Catalase gene is not only influenced by genetic and developmental signals both physical and chemical. Ascorbate peroxidase (EC 1.11.1.11; APX) are heme binding enzymes, plays a key role in detoxification of H2O2 in chloroplast. In plants it is found in cytosol and chloroplast. The activity of APX increases along with CAT and SOD. It reduces H2O2 to H2O by using ascorbic acid as reducer agent.
Environmental stresses such as salt, drought, high temperature and light intensity greatly influences the activity of both Catalase and ascorbate peroxidase. Salinity causes oxidative stress which affects the biochemical and enzymatic component in the plant cell under stressed condition. The main objective of this study was to observe the effect of salt stress on chlorophyll content, protein content and antioxidant enzyme activity of CAT and APX.
MATERIALS AND METHODS
Collection of plant material: The seeds of Pennisetum glaucum (L.) R.Br. cv. WC-C75 was collected from All India Coordinated Pearl Millet Improvement Project, Jodhpur, India.
Salt stress treatment: For the experiments seeds of P. glaucum were first washed with Tween 20 and the further surface sterilized with 0.1% HgCl2 for 3 min and washed thoroughly with autoclaved distilled water and then the seeds were grown in hydroponic condition. The Hoagland’s solution was aerated externally to prevent anaerobic condition and kept in 16 h photoperiod at 25±2°C.
Salt stress treatment was given to the 21 days old seedling. The salt treatment was given by adding known concentration of the salt (NaCl) at concentrations of 50, 100, 150 and 200. The solutions were changed every day to reduce any contamination. The hydroponic system was kept in 16 h photoperiod at temperature 25±2°C.
Preparation of enzyme extract: Total leaf protein was extracted by the Polyvinylpolypyrrolidone (PVPP) precipitation method. Fresh leaf tissues (0.5 g) were homogenized in 50 mM sodium phosphate buffer containing 10% (w/v) insoluble PVPP using a prechilled mortar and pestle. The homogenates were centrifuged at 12,000 rpm for 20 min at 4°C (Remi Instruments, India). The supernatant was collected and kept under -20°C for protein estimation and enzyme assay.
Determination of protein: Protein content was determined according to Bradford (1976) spectrophotometrically at 595 nm, using Comassie brilliant blue G 250 as a protein binding dye (Bradford, 1976). Bovine fraction V was used as a protein standard.
Catalase assay: A modified protocol set by Chandlee and Schanalios (1984) was used to measure the Catalase activity (Chandlee and Schanalios, 1984). In the cuvette 0.2 mL of enzyme extract was mixed with 2 mL of phosphate buffer (pH-7.0) to it 3% H2O2 was added, OD was taken immediately after adding H2O2 at 240 nm as initial reading and after 3 min again OD was taken as final reading. Phosphate buffer was used as a blank for spectrophotometer.
Ascorbate peroxidase assay: A modified protocol of Asada and Takahashi (1987) was followed to measure the Ascorbate peroxidase activity (Asada and Takahashi, 1987). In cuvette 0.1 mL of enzyme extract was mixed with 2 mL of phosphate buffer (pH-7.0) and 0.2 mL 50 μM Ascorbic acid was added to it 3% H2O2 was added and OD was recorded immediately at 290 nm as initial reading and after 3 min final reading was taken. Phosphate buffer was used as a blank reading for spectrophotometer.
Chlorophyll estimation: The chlorophyll content was estimated according to Arnon (1949), tissue pieces were macerated with 80% acetone and pinch of calcium carbonate was also added. The homogenate was centrifuged at 5000 rpm for 10 min; the supernatant was made upto known volume with 80% acetone. OD of the supernatant was taken at 645 and 663 nm against 80% acetone as blank.
All data are expressed as means of triplicate experiments unless mentioned otherwise.
RESULTS
In the present study a significant elevation in antioxidant enzyme activity (Catalase) was recorded while little changes in Ascorbate peroxidase activity was observed. A linear increase in Catalase activity was observed at both 150 and 200 mM concentration of salt with increase in time interval (Fig. 3). But a variation in Catalase activity was observed at 50 and 100 mM (Fig. 3). At 12 h the Catalase activity decreased as compared to control (no salt treatment) and then increased. A maximum of 3 folds increase in Catalase activity was recorded at 200 mM concentration of salt. It was observed that as the salt concentration increased Ascorbate peroxidase activity decreased as compared to control (with no salt concentration) (Fig. 4).
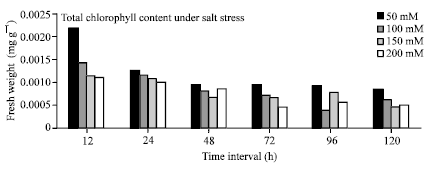 | |
| Fig. 1: | Effect of salt treatment on total chlorophyll content in pearl millet at different time intervals |
 | |
| Fig. 2: | Effect of salt treatment on protein content in pearl millet at different time intervals |
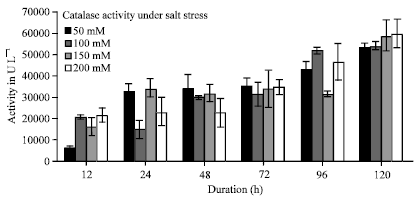 | |
| Fig. 3: | Effect of salt treatment on catalase activity in pearl millet at different time intervals |
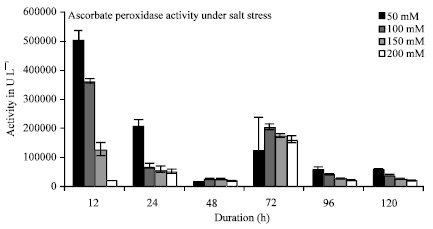 | |
| Fig. 4: | Effect of salt treatment on ascorbate peroxidase activity in pearl millet at different time intervals |
As the duration of salt stress progressed the activity of Catalase was observed to enhance, showing that it plays an active role in ROS scavenging. While Ascorbate peroxidase does not seem to play any important role in ROS scavenging. Catalase has a distinctive role on protection against salt stress in Pearl millet. APX activity reduction with stress may be related to its detoxification (reduction) capacity being below the oxidation capacity of H2O2. As the salt concentration increased the protein content decreased inversely with increasing time interval (Fig. 2). The chlorophyll content also decreased at all the concentrations of salt showing chlorophyll damage due to salt stress (Fig. 1). There is a positive correlation between resistance to oxidative damage and is thought to be resulting from the reduction in enzyme activity that causes a reduction in protein amount. It has been determined that chlorophyll level declines, increased content of proline and free amino acid has been observed in Pearl millet under salt stressed condition.
DISCUSSION
Salinity causes a wide range of responses in plants such as decreased growth, increased osmotic potential and most of important production of ROS due to oxidative stress in the cell. ROS are highly reactive species which readily oxidizes proteins, lipids, nucleic acid and also causes damage to cell membrane. As reported in many plants both enzymatic and non-enzymatic antioxidant plays an important role in scavenging the ROS. From this study we can say that enzymatic antioxidants (CAT and APX) do play an important role in Pearl millet under salt stress. It has already been reported CAT and APX plays a major role in ROS scavenging mechanism under salt stress as reported in Barley, Onion, French bean, Wheat, Rice and Horse gram (Yildiz and Terzi, 2013; Kim et al., 2005; El-Baky et al., 2003; Babu and Devaraj, 2008; Naji and Devaraj, 2011; Hameed et al., 2008; Sahu et al., 2010). As the salt (NaCl) concentration increased the protein content decreased this is due to oxidation of proteins due to ROS generated. Plants have developed Antioxidative defence mechanisms to detoxify/scavenge the Reactive Oxygen Species (ROS) produced due to various environmental stress. The antioxidant defense mechanism includes enzymatic (CAT, APX, SOD, GR and POX) and non enzymatic (proline, free amino acid, glycine betain, ascorbic acid and glutathione). It has already been reported that proline and free amino acid content increased with increased salt (NaCl) concentration in Pearl millet (Sneha et al., 2013). The qualitative changes in the activities of several enzymes including Catalase and Ascorbate peroxidase isolated form various plants subjected to salinity stress were observed by many researchers as mentioned above is similar to the results obtained in case of Pearl millet under salt stress.
ACKNOWLEDGMENTS
The authors would like to thank the Management Committee, Jayoti Vidyapeeth Women’s University, Jaipur, Rajasthan, India for providing the financial assistance and lab facilities.
REFERENCES
- Karimi, G., M. Ghorbanli, H. Heidari, R.A. Khavari Nejad, M.H. Assareh, 2005. The effects of NaCl on growth, water relations, osmolytes and ion content in Kochia prostrate. Biol. Plant, 49: 301-304.
CrossRef - Ashraf, M. and N. Parveen, 2002. Photosynthetic parameters at the vegetative stage and during grain development of two hexaploid wheat cultivars differing in salt tolerance. Biol. Plant, 45: 401-407.
CrossRef - Venkatesan, A. and S. Sridevi, 2009. Response of antioxidant metabolism to NaCl stress in the halophyte Salicornia brachiata roxb. J. Phytol., 4: 242-248.
Direct Link - Koskeroglu, S. and T.A. Levent, 2008. The investigation on growth and dome antioxidative enzymes of the maize (Zea mays L.) plant under salt and water stress. Proceedings of the International Meeting on Soil Fertility Land Management and Agroclimatology, October 29-1 November, 2008, Turkey, pp: 765-775.
- Hernandez, J.A., A. Jimenez, P. Mullineaux and F. Sevilia, 2000. Tolerance of pea (Pisum sativum L.) to long-term salt stress is associated with induction of antioxidant defences. Plant Cell Environ., 23: 853-862.
CrossRefDirect Link - Santos, C.L.V., A. Campos, H. Azevedo and G. Caldeira, 2001. In situ and in vitro senescence induced by KCl stress: Nutritional imbalance, lipid peroxidation and antioxidant metabolism. J. Exp. Bot., 52: 351-360.
Direct Link - Bettaieb, T., M. Mahmoud, J.I. Ruiz de Galarreta and P.D. Jardin, 2007. Relation between the low temperature stress and catalase activity in Gladiolus somaclones (Gladiolus grandiflorus Hort.). Sci. Hortic., 113: 49-51.
CrossRefDirect Link - Xu, Q. and B. Huang, 2004. Antioxidant metabolism associated with summer leaf senescence and turf quality decline for creeping bentgrass. Crop Sci., 44: 553-556.
Direct Link - Foyer, C.H., P. Descourvieres and K.J. Kunert, 1994. Protection against oxygen radicals: An important defence mechanism studied in transgenic plants. Plant Cell Environ., 17: 507-523.
CrossRefDirect Link - Smirnoff, N., 1993. The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol., 125: 27-58.
CrossRefDirect Link - Apel, K. and H. Hirt, 2004. Reactive oxygen species: Metabolism, oxidative stress and signal transduction. Annu. Rev. Plant Biol., 55: 373-399.
CrossRefPubMedDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Chandlee, J.M. and J.G. Scandalios, 1984. Analysis of variants affecting the catalase developmental program in maize scutellum. Theor. Applied Gen., 69: 71-77.
CrossRef - Arnon, D.I., 1949. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol., 24: 1-15.
CrossRefPubMedDirect Link - Kim, S.Y., J.H. Lim, M.R. Park, Y.J. Kim and T.I. Park et al., 2005. Enhanced antioxidant enzymes are associated with reduced hydrogen peroxide in barley roots under saline stress. J. Biochem. Mol. Biol., 38: 218-224.
PubMedDirect Link - El-Baky, A., H. Hanaa, M.A. Amal and M.M. Hussein, 2003. Influence of salinity on lipid peroxidation, antioxidant enzymes and electrophoretic patterns of protein and isoenzymes in leaves of some onion cultivars. Asian J. Plant Sci., 2: 1220-1227.
CrossRefDirect Link - Babu, N.R. and V.R. Devaraj, 2008. High temperature and salt stress response in French bean (Phaseolus vulgaris). Aust. J. Crop Sci., 2: 40-48.
Direct Link - Naji, K.M. and V.R. Devaraj, 2011. Antioxidant and other biochemical defense responses of Macrotyloma uniflorum (Lam.) Verdc. (Horse gram) induced by high temperature and salt stress. Braz. J. Plant Physiol., 23: 187-195.
Direct Link - Hameed, A., S. Naseer, T. Iqbal, H. Syed and M.A. Haq, 2008. Effects of NaCl salinity on seedling growth, senescence, catalase and protease activities in two wheat genotypes differing in salt tolerance. Pak. J. Bot., 40: 1043-1051.
Direct Link - Sahu, S., P. Das, M. Ray and S.C. Sabat, 2010. Osmolyte modulated enhanced rice leaf catalase activity under salt-stress. Adv. Biosci. Biotechnol., 1: 39-46.
CrossRefDirect Link - Sneha, S., A. Rishi, A. Dadhich and S. Chandra, 2013. Effect of Salinity on Seed Germination, Accumulation of Proline and Free Amino Acid in Pennisetum glaucum (L.) R. Br. Pak. J. Biol. Sci., 16: 877-881.
CrossRefDirect Link