
Shahidul Islam
Department of Agriculture, Mail Slot 4913, 153 Woodard Hall, University of Arkansas at Pine Bluff, 1200 North University Drive, Pine Bluff, AR 71601, United State of America
Ehiorobo Izekor
Department of Agriculture, Mail Slot 4913, 153 Woodard Hall, University of Arkansas at Pine Bluff, 1200 North University Drive, Pine Bluff, AR 71601, United State of America
James O. Garner
Department of Agriculture, Mail Slot 4913, 153 Woodard Hall, University of Arkansas at Pine Bluff, 1200 North University Drive, Pine Bluff, AR 71601, United State of America
American Journal of Plant Physiology
Year: 2012 | Volume: 7 | Issue: 6 | Page No.: 252-260







ABSTRACT
Chilling injury is when morphological and physiological damage is sustained by plant tissue that is exposed to freezing temperatures. An experiment was conducted using four chilling tolerant sweet potato genotypes such as 105MS1, 108MS2, 180MS3 and 183MS4, which were selected from seventy nine lines from three major variety crosses for tolerant to chilling injury. The effects of chilling exposure on fatty acids compositions and peroxidase enzyme activity were studied. Chilling exposure increased the Peroxidase Enzyme Activity (PEA). Genotypes differences were also found in PEA following chilling exposure. All four genotypes that were tested had a high level (over 50%) of unsaturated fatty acid on their glycolipids. There were no differences found for the fatty acid percentage composition of the total glycolipid and phospholipid fractions from the four chilling tolerant genotypes except for C18:0 of the glycolipid fraction. However, C18:0 was found in low percentage and was not considered a major fatty acid. Sixty percent of total fatty acid in glycolipid fraction was C18:2 and C18:3. It was concluded that if lipids fatty acid composition was indeed a factor in chilling tolerance, it did not vary among the chilling tolerant genotypes. The result suggests that the genotypes were considered chilling tolerant to chilling environment, but that differences existed in their mechanism for tolerance. Thus, by breeding and selecting for chilling tolerance, it could enhance chilling tolerance in sweet potatoes.
PDF Abstract XML References Citation
Received: July 27, 2012;
Accepted: December 11, 2012;
Published: January 23, 2013
How to cite this article
Shahidul Islam, Ehiorobo Izekor and James O. Garner, 2012. Lipid and Fatty Acid Compositions of Chilling Tolerant Sweet potato (Ipomoea batatas L.) Genotypes. American Journal of Plant Physiology, 7: 252-260.
DOI: 10.3923/ajpp.2012.252.260
URL: https://scialert.net/abstract/?doi=ajpp.2012.252.260
DOI: 10.3923/ajpp.2012.252.260
URL: https://scialert.net/abstract/?doi=ajpp.2012.252.260
INTRODUCTION
Sweet potato (Ipomoea batatas L. Lam.) is an important food crop which is enriched in health beneficial components (Ibaraki and Murakami, 2007; Islam et al., 2009a). In general, sweet potato is growing well under the warm climate, low temperature caused chilling damaged. Chilling injury is the physiological damage sustained by plant tissues exposed to nonfreezing temperature of approximately 0-15°C. A number of mechanisms have already been proposed based on the physiological and biochemical functions associated with chilling injury (Al-Shoaibi, 2008; Saleh, 2007; Khorshidi and Nojavan, 2006; Anjum and Khatoon, 2003; Ali et al., 2000). The severity and length of time required to cause an irreversible dysfunction are generally determined by the temperature extreme, duration of exposure to cold conditions, plant species and morphological and physiological conditions of the plant material at time of exposure (Islam et al., 2009b). Chilling injury disrupts metabolic and physiological processes of higher plants, so it is unlikely that a single basic cause could explain injury. These include an increased concentration of cytosolic calcium; a marked decreased in protoplasmic streaming, an alteration in the cytoskeleton, a conformational change in some enzymes and temperature induced change in the molecular ordering of membrane lipids (Kim et al., 2007). The plasma membrane is regarded as the sensitive site of injury during low temperature stress in herbaceous plants (Zhang and Tian, 2009). Limited component begins to freeze at chilling temperature, the opposite occurred in chilling-resistance species (Sthapit and Witcombe, 1998).
Alterations in Membrane lipids composition during cold-acclimation and low temperature stress has been the focus of investigations for the last 20 years yet conclusive results have not been established (De la Roche et al., 1972). Research findings have shown significant elevated levels of unsaturated fatty acids especially fatty acids associated with phospholipids in response to chilling and freezing temperature stress (Levitt, 1980). Gerloff et al. (1966) found that the fatty acids composition of alfalfa roots changed during hardening due to differential accumulation of polyunsaturated fatty acids. Kuiper (1970) reported difference in lipids composition of alfalfa leaves relatively cold hardiness. On the hand, investigations failed to observe any increased in membrane lipid fatty acids unsaturation during and after acclimation and no recorded information linked membrane fatty acid unsaturation with increased chilling resistance (Vigh et al., 1985; De la Roche et al., 1972). Kenrick and Bishop (1986) showed that the content of high melting point fatty acids of phosphotidylglycerol was negatively correlated to chilling sensitively of plants rather it may be related to genetic origin of the plants. Wang (1982) stated that the primary response of plants to chilling temperature stress is the physical change of the membrane fatty acids composition. There was no correlation between chilling tolerance and the presence of unsaturated fatty acid, when the fatty acid composition of 13 chilling tolerant and sensitive plants were compared (Wilson and Crawford, 1974). According to Moon et al. (1995), study following cold-acclimation, lipids components in thylakoid membrane of tolerant wild-type and transgenic tobacco plants did not significantly differ in photosystem II and photo inhibition response to high or low temperature stress. The results suggest that unsaturation of fatty acids of phosphatidylglycerol in thylakoid membrane of the wild-type tobacco stabilized and enhanced the photosynthetic capacity and the recovery of photo system II protein complex during low temperature stress.
Chemical analyses of proxidase isoenzymes in acclimated chilled plants suggest possible association with cell wall lignification. Several authors (Bassal and El-Hamahmy, 2011; Cao et al., 2011; Lee and Lee, 2000) asserted that location of peroxidase enzymes in cell wall of acclimated seedling is an indicative that antioxidants likely improved and maintained the mechanical strength of chilling sensitive mesocotyl. Hodgson and Raison (1991) reported that carbohydrates were less utilized in comparison with photosynthetic rate by chilling sensitive plants compared with tolerant plant. Enzymes activity increased in sensitive plant partly attributable to the productions of more chilling tolerant isoenzymes. Hetherington et al. (1989) showed that high intensity light hastens the symptoms of chilling injury. Under chilling temperature of 4°C and high light intensity (800 μM m-2 sec-1), approximately 10-15% of the leaf area of mug bean (Vigna radiata) were necrotic, whereas under low light intensity (155 μM m-2 sec-1) no symptoms of chilling damages were apparent. The objectives of this study were: to evaluate lipids compositions and peroxidase enzyme activity among the selected genotypes of sweet potatoes to possibly explain tolerance to chilling injury.
MATERIALS AND METHODS
Plant materials and cultural methods: Plant selection to determine genotypes tolerant to chilling injury from lines of a genetically diverse population was commenced in the greenhouse. Four genotypes namely 105MS1, 108 MS2, 180MS3, 183MS4 were selected from seventy nine lines from three major variety crosses for tolerant to chilling injury (Islam et al., 2009c). The experiment was conducted during 1998 to 2002 at the Mississippi State University, USA. Stem cuttings from each line, approximately 12 cm, were placed in 3.8 L plastic pots containing peat-perlite vermiculite medium (2:1:1 by volume). The medium was amended with 4.56 kg dolomitic lime, 1.82 kg ON-9P-0K, 1.40 kg calcium nitrate and 0.17 kg fritted trace element per cubic meter. Following planting, the plants were kept in a glasshouse under intermittent mist to initiation roots. After five days, plants were moved to another bench in the same glasshouse where they were manually watered once a day for five days. Ten days old plants of each genotype were moved from the glasshouse to walk-in temperature controlled growth chamber maintained at 5°C and a control room 25°C with 85% relative humidity, 10 h photoperiod for duration of 72 h chilling treatment and a fluorescence light intensity of (12 μE M-2 sec-1) suspended over the canopy.
Extraction and analysis of fatty acids: Total lipids analysis was performed on 10 days old sweet potato leaves to determine fatty acids composition of glycolipid and phospholipids. The experimental design was a split-plot with genotypes as the main plot and temperature as the subplot. The data was analyzed as Completely Randomized Design (CRD) with three replications. Isolation and identification of fatty acids in the leaf were performed as outlined by Whitaker (1986) and modified by Phromtong (1993) and Islam et al. (2009b). Table 1 gives the general and chemical characteristics of the fatty acids compositions in sweet potatoes.
Deveined leaf tissue, 3 g fresh weight was grounded in a mortar with liquid nitrogen. The grounded tissues were transferred to 16x100 mm pyrex culture tubes and extracted with hot isopropanol. Isopropanol was added to approximately half the volume of tissue in the tubes and the tubes were incubated in boiling water for five minutes and then cooled at room temperature. Isopropanol was removed from tissue with a Pasteur pipette and refluxed under nitrogen (N2) until dry. Tissue was re-extracted with chloroform-methanol (2:1, v/v). Both extracts were dried under nitrogen and resuspended in chloroform-methanol (2:1, v/v) and combined. Removal of non-lipids’ contaminant was performed as outlined by Folch et al. (1957). Tubes were flushed with nitrogen and sealed at each stage. The combined washed lipids’ extract was reflushed under nitrogen until dry. The extracts were then redissolved with 2 mL chloroform. Total lipids were separated by column chromatography using silicic acid (hydrated silicon dioxide) 100-200 mesh Bio-sil a (Bio-Rad Laboratories, Richmond, CA). The lipids were separated into neutral glycolipids, (fraction 3) and phospholipids (fraction 4 and 5). Fractionated eluted lipids were collected and reflushed under nitrogen, until dry.
| Table 1: | Features of the fatty acid compositions in sweet potatoes |
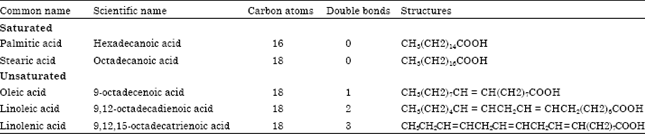 | |
Samples were redissolved in 2 mL of chloroform in 7 mL borosilicate glass vials sealed under nitrogen and stored at-10°C until further use. Transesterification of individual fractions of polar lipids was performed by reflushing samples in 7 mL borosilicate glass vials under nitrogen until dry. Samples were redissolved in 0.5 mL chloroform, followed by addition of 0.5 mL of a 0.6 N potassium hydroxide in dry methanol. Tubes were then flushed with nitrogen, tightly sealed, wrapped with aluminum foil and placed on a roto-torque rotator at room temperature for two hours. Following the addition of 0.5 mL distilled water and 50 μL of 6 N HCl, Fatty Acids Methyl Esters (FAME) were reconverted by extraction with 2 mL of hexane.
Quantitative analysis of FAME was performed by Gas Liquid Chromatography (GLC) on a Varian 3300 Gas Chromatograph (GC, Varian Associates, Sugar Land TX), equipped with a flame ionization detector and utilizing a supelcowax 10 fused silica wide bore capillary column, 30 m in lengthx0.53 mm i.d. and 10 μL films thickens (Supelco, Inc.,Bellenfonte). Injector and detector temperature were 250 and 300°C, respectively.
Helium and nitrogen gas were used as carrier and makeup gas at a flow rate of 6 and 24 mL min-1, respectively. Initial column temperature was 190°C and after three minutes initial holding time, the column oven temperature was raised 6°C min-1, to a final temperature of 220°C and maintained for 20 min. The instrument (GC) was connected to a Varian 4290 Integrator, for data collection. Standards of FAME mixture GLC Reference #6 (Alltech associates. Inc. Deer Field, III) were injected for qualitative and qualitative analysis.
Peroxidase enzyme activity: The activity of peroxidase as crude soluble enzyme and cell wall bound enzymes was determined from 10 days old sweet potato leaves. The experimental design was a split-plot with genotypes as the main plots and temperature as the subplots. Data was analyzed in a Completely Randomized Design (CRD) with three replications. The extracts for enzyme analysis were prepared by grinding 3 g fresh weight of deveined leaf tissue in 9 mL cold citric phosphate buffer (pH 4.5), using a pre-chilled mortar and pestle. The homogenate was transferred to plastic centrifuge tubes and centrifuged in a Sorvall RC-5B automatic super speed refrigerated centrifuge (Sorvall Instrument, Du point Co., Wilmington, DL) at 10,000 xg for 10 min. The supernatant was saved as the soluble enzymes fraction. The remaining pellets were washed with 9 mL deionized water and centrifuged at 10,000 xg, for 10 min. The supernatant was discarded and the pellets resuspended in 9 mL of 0.2 M calcium chloride (CaCl2) solution and centrifuged at 10,000 xg, for 10 min. The supernatant was saved in 7 mL borosilicate glass vial and analyzed as crude cell wall bound enzymes. Peroxidase, activity was measured according to Venkatarayappa et al. (1984) and Islam et al. (2009b).
Peroxidase activity determinations were made by adding 50 μL of the enzyme extract to 3.0 mL of 0.05 M phosphate buffer (pH 4.5) and 50 μL guaiacol solutions (20 mM). The reaction was initiated by adding 0.5 mL of 8 mM H2O2 (0.03%) to the mixture. The change in absorbance at 420 nm was followed against a blank containing phosphate buffer. Activity was expressed as the inverse of the change in absorbance over time every 10 seconds up to 60 seconds.
Statistical analysis: Data for the different parameters were analyzed by analysis of variance (ANOVA) using the General Linear Models procedures of SAS version 8.1. Mean separations were done using Fisher’s protected Least Significant Difference (LSD) tests.
RESULTS AND DISCUSSION
Lipid and fatty acid composition of chilling tolerant sweet potato genotypes: There was no effect of chilling temperature on the fatty acid composition of leaves from the four sweet potatoes studied (data not shown). The fatty acid composition of the glycolipid fraction of the leaves from the genotypes is given in Table 2. Linolenic (18:3) and palmitic (16:0) were the major fatty acids, at approximately 50% (47 to 58%) and 25% (23 to 29%) of the total fatty acids in the glycolipids. No genotypes differences in percent composition were found for palmitic (16:0), oleic (18:1), linoleic (18:2) or linolenic (18:3). Statistical differences among the genotypes were found in oleic (18:0) acid only. Fatty acid composition of genotypes 180MS3 was higher (5.60) than the remaining genotypes tested. However, oleic (18:0) acid ranged from 1.75 to 5.6% in composition of the total fatty acids of the glycolipid fraction.
There were no differences in the fatty acid composition of the total phospholipids from the leaves of the four sweet potato genotypes (Table 3). The major lipids of the phospholipids fraction included C16:0, C18:0, C18:2. C16:0 ranged in compositions from 40.6 to 44.8%. C18:2 and C18:3 were approximately 22 and 25%, respectively. The minimum response obtained for fatty acids compositions, indicated that the four genotypes were not influenced differently by the fatty acids makeup of their lipids. However, individual lipids are not examined. Woods et al. (1991) reported that only minor changes in fatty acids compositions of sweet potato chloroplast membrane occurred following chilling. Bartkowski et al. (1978) studied the effects of chilling on long staple cotton membrane with respect to a change in unsaturated fatty acids composition. Their results showed that chilling effects were localized in the microsomal membrane with a significant decrease in unsaturated fatty acid level in the mitochondrial membrane. The nuclear membrane exhibited no change following chilling treatment.
| Table 2: | Effect of genotype on major fatty acids composition of the glycolipid fraction of chilling tolerant genotype following exposure average of 25 and 5°C |
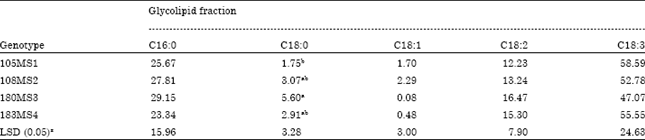 | |
| zLSD (0.05); comparison of means between genotypes, Means followed by the same letter not differ at p<0.05 level | |
| Table 3: | Effect of genotype on fatty acids composition of the phospholipid fraction of chilling tolerant genotype following exposure average of 25 and 5°C, respectively for 24 h |
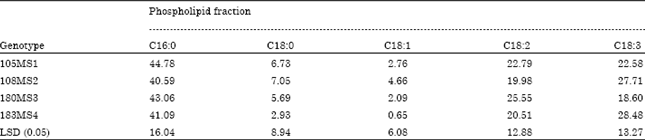 | |
| No statistical difference at p<0.05 was found | |
Additionally, their data showed that greatest change in unsaturated fatty acid level during cotton chilling occurs with linoleic acids and linolenic acids of the microsomal membrane. They found that the limiting factor in chilling resistance of higher plant is the ability of the microsomes to desaturated fatty acids and efficiently distribute these unsaturated fatty acids to other membranes. This limiting factor may possibly be related to the diminishing effects of lower temperature on Phospholipids Exchange Proteins (PEP) which transfers phospholipids from the microsomes to the mitochondrial (Bartkowski et al., 1978). Murata et al. (1982) worked with blue green algae cell (Anacystis nidulans) and found that when temperature was reduced from 38 to 28°C, higher desaturation and reduction in hydrocarbons chain of the glycerol moiety of lipids occurred. Consequently, reduction in chain length and unsaturation of the fatty acids was suggested to have enhanced the fluidity and lowered the transition temperature of lipids phase of the thylakoid membrane. Palta et al. (1993), comparing cold-acclimated wild types and non-acclimated genotypes Solanum species, found that cold-acclimation wild types had increased unsaturated fatty acids, free sterol, sitosterol and slight decrease in cerebroside compared with non-acclimated genotypes. Cold-acclimated wild types had increased total lipids and phospholipids of 17 and 25%, respectively compared with non-acclimated genotypes which had a lower ration of sterol phospholipids ratio. Yamaki and Uritani (1972), found that more than 90 molar percent of the total lipids in the mitochondrial membrane of sweet potato are phospholipids and composed of the following components: phosphatidylethanolamine, phosphatidylcholine and lysophosphatidylcholine collectively rich in (18:2) linoleic. These components were reported to have decreased 40, 20 and 10%, respectively following chilling treatment compared with the control, according to Yamaki and Uritani (1974). These studies also found that 65-molar percent of the total mitochondria membrane fatty acids of white potato phospholipids, including components reported above were not altered following treatment. The chilling tolerance difference between two genotypes to fatty acid composition could not be related. Phromtong (1993), showed the C18:3 increased with cold treatment in a chilling tolerant sweet potato genotypes on the phospholipids fraction of the total lipids. Therefore, it is concluded that the mode of tolerance among genotypes tested in this study was not different with respect to fatty acid composition.
Peroxidase enzyme activity: The genotypes 180 MS3 and 183 MS4 had higher peroxidase enzyme activity compared to the genotypes 105 MS1 and 108MS2 (Fig. 1, 2). The chilling temperature showed significantly higher peroxidases activity as compared to higher temperature (25°C) studied. Similar results obtained by El-hilali et al. (2003). They reported that the peroxidase activity increases continuously at 4°C over the period of storage in fortune mandarin fruits.
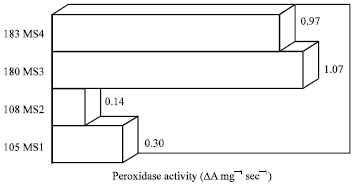 | |
| Fig. 1: | Peroxidase enzyme activity of chilling tolerant sweet potato genotypes of studied |
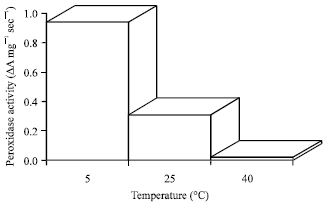 | |
| Fig. 2: | Peroxidase enzyme activity of chilling tolerant genotypes of sweet potatoes following 24 h of 5°C chilling, 25 and 40°C not chilled exposure chilling exposure |
If the peroxidase enzyme activity is a factor in chilling tolerance as suggested by Woods et al. (1991), a difference in the tolerance mechanism is indicated. Chilling sensitive and chilling tolerant plant’s responses to low temperature stress by producing differential level of toxic oxygen compounds were suggested to be the products metabolisms (Li et al., 2011; Zahra et al., 2009; Lin et al., 2006; Hodgson and Raison, 1991). Superoxide dismutase (SOD), catalyzes (CAT) and various peroxidases and ascorbate peroxidases, glutathione reductases constitute the cellular defense mechanism against oxidative stress (Bowler and Chua, 1994). The results revealed that genotypes 180MS3 and 183MS4 may have a better protective mechanism with respect to peroxidase enzyme activities. Usually peroxidases are ubiquitous enzymes that have diverse biochemical functions in higher plants and are involved in the response of plants to chilling stress (Li et al., 2011; Safizadeh et al., 2007). Thus, the tolerance of 105MS1 and 108MS2 could be from other physiological mechanisms (Badea and Basu, 2009; Falcone et al., 2004).
From the aforementioned results and discussion, it was clinched that selecting for chilling tolerance could enhance the chilling tolerance in sweet potatoes. The basis or biochemical mechanism for chilling tolerance was not the same for the four sweet potato genotypes tested; therefore combining traits for tolerance could lead to higher tolerance levels.
REFERENCES
- Ali, A., C. Stushnoff and D.L. Johnson, 2000. Negative association of endogenous sorbitol with cold hardiness in lentil. Pak. J. Biol. Sci., 3: 2026-2029.
CrossRefDirect Link - Al-Shoaibi, A.A., 2008. Photosynthetic response to the low temperature in elephant grass (Pennisetum purpureum) and Zea mays. Int. J. Bot., 4: 309-314.
CrossRefDirect Link - Anjum, Z.I. and A. Khatoon, 2003. Chilling effect on germination and seedling vigour of some cultivated species of Gossypium. Asian J. Plant Sci., 2: 297-299.
CrossRefDirect Link - Bartkowski, E.J., F.R.H. Katterman and D.R. Buxton, 1978. Influence of exogenous fatty acids on cottonseed germination. Physiol. Plant., 44: 153-156.
CrossRef - Bassal, M. and M. El-Hamahmy, 2011. Hot water dip and preconditioning treatments to reduce chilling injury and maintain postharvest quality of Navel and Valencia oranges during cold quarantine. Postharvest Biol. Tech., 60: 186-191.
CrossRef - Badea, C. and S.K. Basu, 2009. The effect of low temperature on metabolism of membrane lipids in plants and associated gene expression. Plant Omics J., 2: 78-84.
Direct Link - Bowler, C. and N.H. Chua, 1994. Emerging themes of plant signal transduction. Plant Cell, 6: 1529-1541.
PubMed - Cao, S., Z. Yang, Y. Cai and Y. Zheng, 2011. Fatty acid composition and antioxidant system in relation to susceptibility of loquat fruit to chilling injury. Food Chem., 127: 1777-1783.
CrossRef - El-hilali, F., A. Ait-Oubahou, A. Remah and O. Akhayat, 2003. Chilling injury and peroxidase activity changes in Fortune mandarin fruit during low temperature storage. Bulg. J. Plant Physiol., 29: 44-54.
Direct Link - Falcone, D.F., P.J. Ogas and C.R. Somerville, 2004. Regulation of membrane fatty acid composition by temperature in mutants of Arabidopsis with alterations in membrane lipid composition. BMC Plant Biol., Vol. 4.
CrossRef - Folch, J., M. Lees and G.H.S. Stanley, 1957. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem., 226: 497-509.
CrossRefPubMedDirect Link - Gerloff, E.D., T. Richardson and M.A. Stahmann, 1966. Changes in fatty acids of alfalfa roots during cold hardening. Plant Physiol., 41: 1280-1284.
CrossRefDirect Link - Hetherington, S.E., J. He and R.M. Smillie, 1989. Photoinhibition at low temperature in chilling-sensitive and -resistant plants. Plant Physiol., 90: 1609-1615.
PubMedDirect Link - Hodgson, R.A.J. and J.K. Raison, 1991. Superoxide production by thylakoids during chilling and its implication in the susceptibility of plants to chilling-induced photoinhibition. Planta, 183: 222-228.
CrossRefDirect Link - Ibaraki, Y. and J. Murakami, 2007. Distribution of chlorophyll fluorescence parameter Fv/Fm within individual plants under various stress conditions. Acta Hortc., 761: 255-260.
Direct Link - Islam, I., A.U. Shaikh and I.M. Shahidul, 2009. Antioxidative and antimutagenic potentials of phytochemicals from Ipomoea batatas (L.) Lam. Int. J. Cancer Res., 5: 83-94.
CrossRefDirect Link - Islam, S., E. Izekor and J.O. Garner, 2009. Development of selection procedures of sweetpotato genotypes according to chilling tolerance. J. Food Agric. Environ., 7: 237-245.
Direct Link - Islam, S., R.C.R. Carmen and J.O. Jr. Garner, 2009. Carbohydrate compositions and peroxidase activity in ungerminated, cotyledon and embryo tissues of Vigna unguiculata L. Walp seed grown under stress temperatures. Am. J. Plant Physiol., 4: 9-17.
CrossRefDirect Link - Kenrick, J.R. and D.G. Bishop, 1986. The fatty acid composition of phosphatidylglycerol and sulfuquinovosyldiacylglycerol of higher plants on relation to chilling sensitivity. Plant Physiol., 81: 946-949.
Direct Link - Khorshidi, M. and A.M. Nojavan, 2006. The effects of abscisic acid and CaCl2 on the activities of antioxidant enzymes under cold stress in maize seedlings in the dark. Pak. J. Biol. Sci., 9: 54-59.
CrossRefDirect Link - Kim, Y.H., S.M. Han, J.C. Lee, W.K. Song and J.W. Bang et al., 2007. Differential expression of 10 sweetpotato peroxidases in response to sulfur dioxide, ozone and ultraviolet radiation. Plant Physiol. Biochem., 45: 908-914.
CrossRef - Lee, D.H. and C.B. Lee, 2000. Chilling stress-induced changes of antioxidant enzymes in the leaves of cucumber: In gel enzyme activity assays. Plant Sci., 159: 75-85.
CrossRefDirect Link - Li, Q., B. Yu, Y. Gao, A.H. Dai and J.G. Bai, 2011. Cinnamic acid pretreatment mitigates chilling stress of cucumber leaves through altering antioxidant enzyme activity. J. Plant Physiol.
CrossRef - Lin, K.H., F.H. Pai, S.Y. Hwang and H.F. Lo, 2006. Pre-treating paclobutrazol enhanced chilling tolerance of sweetpotato. Plant Growth Regul., 49: 249-262.
CrossRef - Moon, B.Y., S.I. Higashi, Z. Gombos and N. Murata, 1995. Unsaturation of the membrane lipids of chloroplasts stabilizes the photosynthetic machinery against low-temperature photoinhibition in transgenic tobacco plants. Proc. Natl. Acad. Sci. USA., 92: 6219-6223.
Direct Link - Murata, N., N. Sato, N. Takahashi and Y. Hamazaki, 1982. Compositions and positional distributions of fatty acids in phospholipids from leaves of chilling-sensitive and chilling-resistant plants. Plant Cell Physiol., 23: 1071-1079.
Direct Link - Palta, J.P., B.D. Whitaker and L.S. Weiss, 1993. Plasma membrane lipids associated with genetic variability in freezing tolerance and cold acclimation of Solanum species. Plant Physiol., 103: 793-803.
PubMed - Safizadeh, M.R., M. Rahemi and M. Aminlari, 2007. Effect of postharvest calcium and hot-water dip treatments on catalase, peroxidase and superoxide dismutase in chilled lisbon lemon fruit. Int. J. Agric. Res., 2: 440-449.
CrossRefDirect Link - Sthapit, B.R. and J.R. Witcombe, 1998. Inheritance of tolerance to chilling stress in rice during germination and plumule greening. Crop Sci., 38: 660-665.
Direct Link - Venkatarayappa, T., R.A. Fletcher and J.E. Thompson, 1984. Retardation and reversal of senescence in bean leaves by benzyladenine and decapitation. Plant Cell Physiol., 25: 407-418.
Direct Link - Vigh, L., F. Joo, M. Droppa, L.I. Horvath and G. Horvath, 1985. Modulation of chloroplast membrane lipids by homogenous catalytic hydrogenation. Eur. J. Biochem., 147: 477-481.
PubMed - Wang, C.Y., 1982. Physiological and biochemical responses of plants to chilling stress. Hortscience, 17: 173-186.
Direct Link - Wilson, J.M. and R.M.M. Crawford, 1974. The acclimatization of plants to chilling temperatures in relation to the fatty-acid composition of leaf polar lipids. New Phytol., 73: 805-820.
CrossRef - Whitaker, B.D., 1986. Fatty-acid composition of polar lipids in fruit and leaf chloroplasts of “16:3”- and “18:3”-plant species. Planta, 169: 313-319.
CrossRefPubMedDirect Link - Woods, F.M., J.O. Garner, J.L. Silva and C. Phromtong, 1991. Estimation of chilling sensitivity in leaves of sweet potato by chlorophyll fluorescence and electrolyte leakage. Phyton, 51: 33-37.
Direct Link - Yamaki, S. and I. Uritani, 1972. The mechanism of chilling injury in sweet potato VI. Changes of lipids components in the mitochondrial membrane during chilling storage. Plant Cell Physiol., 13: 67-79.
Direct Link - Yamaki, S. and I. Uritani, 1974. Mechanism of chilling injury in sweet potato. XI. Irreversibility of physiological deterioration. Plant Cell Physiol., 15: 385-388.
Direct Link - Zahra, P., R. Majid and B. Amin, 2009. Seasonal changes of peroxidase, polyphenol oxidase enzyme activity and phenol content during and after rest in pistachio (Pistacia vera L.) flower buds. World Applied Sci. J., 6: 1193-1199.
Direct Link - Zhang, C. and S. Tian, 2009. Crucial contribution of membrane lipids unsaturation to acquisition of chilling-tolerance in peach fruit stored at 0°С. Food Chem., 115: 411-415.
CrossRef - Saleh, A.A.H., 2007. Amelioration of chilling injuries in mung bean (Vigna radiata L.) seedlings by paclobutrazol, abscisic acid and hydrogen peroxide. Am. J. Plant Physiol., 2: 318-332.
CrossRefDirect Link