
Pradeep Kumar Patel
Department of Plant Physiology, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi-221005, India
A. Hemantaranjan
Department of Plant Physiology, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi-221005, India
American Journal of Plant Physiology
Year: 2012 | Volume: 7 | Issue: 4 | Page No.: 164-173







ABSTRACT
Significant differences in the antioxidant system of the leaves of four chickpea (Cicer arietinum L.) genotypes differing in tolerance to drought were observed in a pot experiment. Drought stress was imposed at pre- and post- anthesis stage (50 and 65 days after sowing). We studied the effect of drought stress on hydrogen peroxide (H2O2) content, lipid peroxidation and antioxidant enzyme activities in four chickpea genotypes viz., Tyson, ICC 4958, JG 315 and DCP 92-3. Drought treatment significantly increased hydrogen peroxide accumulation and lipid peroxidation, which indicated the extent of oxidative injury posed by stress conditions. Enzyme activities of Superoxide Dismutase (SOD), Peroxidase (POX), Catalase (CAT) and Ascorbate Peroxidase (APX) increased in chickpea leaves as a consequence of drought conditions and all the enzyme activities were significantly higher in drought stressed genotypes i.e., Tyson and ICC 4958 than in JG 315 and DCP 92-3. Relationship between Lipid Peroxidation (LPO), Hydrogen peroxide (H2O2), Superoxide Dismutase (SOD), Catalase (CAT), Peroxidase (POX) and Ascorbate Peroxidase (APX) was determined in order to find out whether these parameters can be used as selection criteria for drought tolerance in this crop. Early drought stress (pre-anthesis drought) was found to be more damaging than the late drought stress (post-anthesis drought).
PDF Abstract XML References Citation
Received: December 30, 2011;
Accepted: February 14, 2012;
Published: May 16, 2012
How to cite this article
Pradeep Kumar Patel and A. Hemantaranjan, 2012. Antioxidant Defence System in Chickpea (Cicer arietinum L.): Influence by Drought Stress Implemented at Pre- and Post-anthesis Stage. American Journal of Plant Physiology, 7: 164-173.
DOI: 10.3923/ajpp.2012.164.173
URL: https://scialert.net/abstract/?doi=ajpp.2012.164.173
DOI: 10.3923/ajpp.2012.164.173
URL: https://scialert.net/abstract/?doi=ajpp.2012.164.173
INTRODUCTION
Chickpea (Cicer arietinum L.) is the third most important grain legume crop in the world and first in the Mediterranean basin and South Asia that frequently experiences water stress during pod set and seed filling stage (terminal drought) in India and the Mediterranean basin, leading to a substantial yield loss (Turner et al., 2001). Drought stress during seed filling has been reported to be highly detrimental to yield in chickpea (Davies et al., 1999). In the current scenario of water limitation, there is little scope to increase the irrigated area of this crop. Moreover, the slow and steady migrations of this crop towards lower latitudes in India (Gowda et al., 2009).
Plants respond to drought stress and acclimatize through various physiological and biochemical changes (Farooq et al., 2009a). Drought induces oxidative stress in plants, in which reactive oxygen species (ROS), such as superoxide radical (O2-●), hydroxy radical (●OH), hydrogen peroxide (H2O2) and alkoxy radical (RO●) are produced (Munne-Bosch and Penuelas, 2003; Esfandiari et al., 2008). Hydrogen peroxide (H2O2) is the first stable compound among ROS produced in the plant cell under normal conditions and as a result of stress. It is suggested to be involved in ROS-mediated signaling because of its ability to penetrate the plasma membrane as an uncharged molecule. Therefore, it can be transported to the site of action (Foyer et al., 1997).
Oxidative damage in the plant tissue is alleviated by a concerted action of both enzymatic and non-enzymatic antioxidant metabolism (Hasegawa et al., 2000). These mechanisms include β-carotenes, Ascorbic Acid (AA), α-tocopherol (α-Toc), reduced glutathione (GSH) and enzymes including Superoxide Dismutase (SOD), Peroxidase (POX), Ascorbate Peroxidase (APX), Catalase (CAT), Polyphenol Oxidase (PPO) and Glutathione Reductase (GR) (Prochazkova et al., 2001).
Plants experience drought stress either when the water supply to roots is interrupted or when transpiration rate becomes very high. These two conditions often coincide under arid and semiarid climates. Drought stress tolerance has been seen in almost all plant species but its extent varies from species to species (Lin et al., 2006). To date, antioxidant responses of chickpea under abiotic stress conditions such as chilling (Nayar et al., 2005) salt (Eyidogan and Oz, 2007) and also drought (Moinuddin and Imas, 2007) have been reported. Drought stress altered lipid peroxidation, increasing peroxidase and superoxide dismutase activity (Saleh et al., 2007; Tatar and Gevrek, 2008). However, no data are available related to comparative antioxidant study under drought at two critical phases of development i.e., pre- and post- anthesis stages in chickpea. Therefore, the aims of this study were: (1) to understand how leaves of plants differing in drought tolerance respond to oxidative damage, (2) to characterize the genotypic drought tolerance variability in chickpea, (3) to identified the most drought-tolerant genotypes, (4) to determine the selection criteria of drought stress and (5) to identify the most sensitive stage of drought stress.
MATERIALS AND METHODS
Site description: The experiment was carried out during Rabi season 2009-10 in rain-protected wire- house at the Horticulture Research Farm, Institute of Agricultural Sciences, Banaras Hindu University Varanasi, India. The experimental area lies between latitudes 25.18°N, longitude 83.03°E and 123.93 m altitude. The average of climatic conditions calculated during the entire growth period is as follows: maximum/minimum temperature, Relative Humidity (RH) was 28.0°C /13.6°C, 71.3/36.5%, respectively and an average sunshine hour was 6.9.
Plant materials and treatment pattern: The seeds of chickpea (C. arietinum L.) genotypes (Tyson, ICC 4958, JG 315 and DCP 92-3) were obtained from Indian Institute of Pulse Research (IIPR-ICAR), Kanpur, India through Material Transfer Agreement (MTA). The healthy seeds of uniform size were selected and surface sterilized with 0.2% HgCl2 solution followed by repeated washing with Double Distilled Water (DDW). Thereafter 10 seeds of each genotype were subsequently sown (10 per pot), pot size “30x30 cm” filled with farm soil having 12-14% moisture at the time of sowing and were thinned to six uniform plants per pot at the first true leaf stage. The experimental soil was sandy loam containing organic carbon 0.31%, available nitrogen 228.00 kg ha-1, available phosphorus 17.00 kg ha-1, available potassium 180.00 kg ha-1 and pH 7.3. All the pots were applied with measured dose of fertilizer for chickpea at 20, 40, 20 kg ha-1 of N, P2O5 and K2O, respectively.
Drought stress applications: Each genotype was grouped in three sets viz., irrigated, early drought stress and late drought stress. Water stress treatment was instigated at 50 DAS.
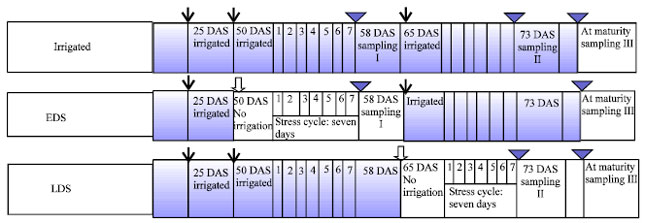 | |
| Fig. 1: | Schematic representation of experimental layout, |
Normal plants were given three irrigations (at 25, 50 and 65 DAS) from the date of sowing to maturity. Early drought stressed or pre-anthesis drought stressed plants (EDS) received two irrigations (25 and 65 DAS) whereas post-anthesis drought stressed plants (LDS) received two irrigations at 25 and 50 DAS. Stress cycle given seven days in both the conditions (Fig. 1).
Methodology
Hydrogen peroxide estimation: Hydrogen peroxide was estimated with titanium reagent (Terenashi et al., 1974). 1 g of titanium dioxide and 10 g potassium sulphate was digested in 150 mL concentrated sulphuric acid over a hot plate for 2 h. The digested mixture was diluted to 500-600 mL and stirred with a magnetic stirrer cum heater at 70-80°C till a clear transparent solution was obtained. This solution was diluted to 1.5 L and stored in a dark brown bottle and used as titanium reagent. Sample preparation and H2SO4 estimation was done as described by Mukherjee and Choudhuri (1983). Leaf material (0.5 g) was homogenized in 10 mL of cold acetone (90%, v/v). The homogenate was filtered through Whatman No. 10 filter paper. To the filtrate, 4 mL of titanium reagent was added followed by 5 mL of concentrated ammonium solution to precipitate the peroxide-titanium complex. The reaction mixture was centrifuged in a refrigerated centrifuge for 5 min at 10000x g, the supernatant was discarded and precipitate was dissolved in 10 mL of 2 M H2SO4. It was re-centrifuged to remove the undissolved material and the absorbance was recorded at 415 nm against blank. Concentration of H2O2 was determined using a standard curve plotted with known concentration of H2O2. All spectro-photometric measurements were carried out by spectrophotometer (Shimadzu UV-VIS, Japan).
Lipid peroxidation: The level of lipid peroxidation was measured in terms of malondialdehyde (MDA) content, a product of lipid peroxidation (Hodges et al., 1999). Leaf sample (0.5 g) was homogenized in 10 mL of 0.1% TCA. The homogenate was centrifuged at 15000x g for 5 min. To 1.0 mL aliquot of the supernatant, 4.0 mL of 0.5% thiobarbituric acid (TBA) in 20% TCA was added. The mixture was heated at 95°C for 30 min and then quickly cooled in an ice bath. After centrifugation at 10000x g for 10 min, the absorbance of supernatant was recorded at 532 nm. The value for nonspecific absorption at 600 nm was subtracted. The MDA equivalent was calculated as follows:
Antioxidant enzyme assays: Superoxide dismutase (EC 1.15.1.1) assay was performed as per the protocol of Dhindsa et al. (1981). Leaf sample (100 mg) was homogenized in 5 mL extraction buffer (0.1 M phosphate buffer of pH 7.5 containing 0.5 mM EDTA). Enzyme extract was centrifuged at 10,000 x g for 10 min at 4°C. After centrifugation, supernatant was collected and used as enzyme source. Reaction mixture (3.0 mL) consisted of 0.1 mL 1.5 M sodium carbonate, 0.2 mL 200 mM methionine, 0.1 mL 2.25 mM NBT, 0.1 mL 3 mM EDTA, 1.5 mL 100 mM potassium phosphate buffer, 0.7 mL distilled water and 0.2 mL enzyme extract. The reaction was started by adding 0.1 mL riboflavin (60 μM) and placing the tubes below a light source of two 15 W florescent lamps for 15 min. Illuminated and non-illuminated reactions without enzyme were used for calibration. Absorbance was recorded at 560 nm in spectrophotometer. One unit of SOD activity was defined as 50% inhibition of the basic rate of the reaction.
Peroxidase (EC 1.11.1.7) activity was estimated as per the protocol of Kar and Mishra (1976) with slight modifications. The enzyme was extracted by homogenizing 100 mg leaf tissue in 5.0 mL 0.1 M phosphate buffer (pH 6.4). The crude extract was centrifuged at 10,000 x g for 20 min at 4°C. Supernatant was collected. Reaction mixture was prepared by adding 4.5 mL 0.1 M phosphate buffer (pH 6.4), 0.2 mL pyrogallol (50 μM) and 0.1 mL 50 μM H2O2 and 0.2 mL supernatant. The mixture was incubated at 25°C for 5 min. Then 0.5 mL of 5.0% H2SO4 was added to terminate the reaction. Absorbance was recorded at 420 nm. One unit of enzyme activity is defined as that amount of enzyme, which forms 1 mol of purpurogallin formed per minute under the assay conditions.
Catalase (EC 1.11.1.6) activity was assayed as per the protocol of Aebi (1983) with suitable modifications.100 mg leaf tissue was homogenized in 5 mL of 0.1 M phosphate buffer (pH 6.4) in a chilled pestle and mortar. The extract was centrifuged at 10,000 x g for 20 min at 4°C. Supernatant was collected and used in the enzyme assay. Reaction mixture consisted of 2.6 mL 0.1 M phosphate buffer (pH 6.4), 0.2 mL enzyme extract and 0.1 mL, 10 mM H2O2 with final volume 3.0 mL. The reaction mixture was mixed rapidly at room temperature. Changes in absorbance at 230 nm at an interval of 15 s were noticed for 2 min on an UV visible spectrophotometer. One unit of catalase activity was defined as the amount of enzyme catalyzing the decomposition of 1 nmol H2O2 per minute (extinction coefficient 36 mM cm-1).
Ascorbate peroxidase (EC 1.11.1.11) was assayed as per the protocol of Nakano and Asada (1980). Enzyme extract for APX was prepared by grinding 100 mg sample with 5.0 mL of extraction buffer (0.1 M phosphate buffer pH 7.5 containing 0.5 mM EDTA and 1 mM ascorbic acid). The homogenate was centrifuged at 10,000x g for 10 min at 4°C. After centrifugation, supernatant was collected and was used as enzyme source. The 3 mL reaction mixture contained 50 mM potassium phosphate buffer (pH 7.0), 0.5 mM ascorbic acid, 0.1 mM EDTA, 0.1 mM H2O2, 0.2 mL enzyme extract. Reaction was started by adding H2O2. Decrease in absorbance for a period of 30 sec was measured at 290 nm in an UV-visible spectrophotometer. One unit was defined as micromoles of ascorbate oxidized per minute (extinction coefficient 2.8 mM cm-1).
Statistical analysis: All experimental data recorded were average mean values for at least three independent assays with three replicates each. The data were subjected to ANOVA for completely randomized design factorial. Differences at p<0.05 were considered statistically significant (Gomez and Gomez, 1984).
RESULTS AND DISCUSSION
Effect of drought stress on malondialdehyde (MDA) content in four chickpea (C. arietinum L.) genotypes are presented in Fig. 2. Lipid peroxidation estimated as MDA content was varied among the genotypes and, it was generally increased under both, early and late drought stress as compared to optimal conditions. Lipid peroxidation was significantly higher in pre- anthesis compare to post-anthesis, as for as genotype is concern, JG 315 (3.23 nmol g-1 fresh weight (FW)) showed higher lipid peroxidation over control (1.19 nmol g-1 FW) followed by genotype DCP 92-3 (3.11 nmol g-1 FW) whereas, minimum was recorded in ICC (1.51 nmol g-1 FW) (Fig. 3). Drought induces oxidative stress in plants by generation of reactive oxygen species (ROS) (Farooq et al., 2009b). As increasing ROS level causes oxidative damage to biomolecules such as lipids, proteins and nucleic acids (Mittler, 2002). Reactive oxygen species e.g., peroxides of polyunsaturated fatty acids generate malondialdehyde (MDA) on decomposition and in many cases MDA is the most abundant individual aldehydic lipid breakdown product (Esterbauer and Cheeseman, 1990) and strikingly malondialdehyde (MDA) is a widely used marker of oxidative lipid injury whose concentration varies in response to abiotic stress (Moller et al., 2007; Davey et al., 2005). A decrease in membrane stability reflects the extent of lipid peroxidation caused by ROS. Furthermore, lipid peroxidation is an indicator of the prevalence of free radical reaction in tissues. Present findings support that the prevention of membrane damage may be related to the induction of antioxidant responses which protects the plant from oxidative damage.
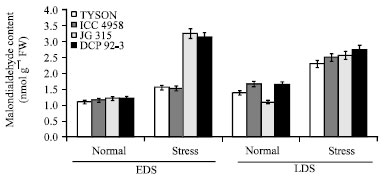 | |
| Fig. 2: | Effect of drought stress on Malondialdehyde (MDA) content in four chickpea (C. arietinum L.) genotypes |
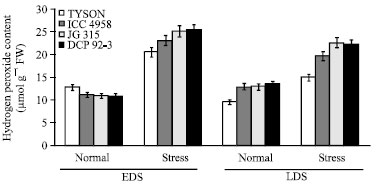 | |
| Fig. 3: | Effect of drought stress on H2O2 content of four chickpea (C. arietinum L.) genotypes |
Hydrogen peroxide (H2O2) concentrations of the genotypes greatly differed among the genotypes in optimal and both pre- and post-anthesis (early and late) drought stress conditions. The accumulation of H2O2 as a result of drought stress was generally increased in the genotypes of chickpea. Hydrogen peroxide was higher in pre- anthesis compare to post-anthesis, as for as genotype is concern, JG 315 (25.10 μmol g-1 FW) showed higher H2O2 content whereas, minimum was recorded in Tyson (20.60 μmol g-1 FW) (Fig. 3). Under drought conditions, plants produce reactive oxygen species, which are harmful to plant growth due to their detrimental effects on the sub-cellular components and metabolism of the plant leading to the oxidative destruction of the cells. It is generally accepted that might be converted to hydrogen peroxide (H2O2). Hydrogen peroxide is a toxic compound produced as a result of scavenging of superoxide radical and its higher concentrations are injurious to plants, resulting in lipid peroxidation and membrane injury (Sairam et al., 1998; Baisak et al., 1994; Menconi et al., 1995). In the present work, in parallel to H2O2 accumulation, membrane damage (MDA content and membrane permeability) of the chickpea cultivars were generally increased. But there were no significant relationship between the H2O2 concentration and drought resistance. In addition to this membrane permeability positively correlated with MDA at both stress conditions and lipid peroxidation in the stress-tolerant genotypes (Tyson and ICC 4958) were found to be lower as compared to stress-susceptible genotypes (JG 315 and DCP 92-3). Higher membrane stability (lower MDA content) has also been reported.
Antioxidant defense system plays vital role in plant’s tolerance to stressful conditions. It is obvious from the results that drought has induced antioxidant enzymes activities more than the normal plants during the drought period. Results showed that increase in SOD, POX CAT and APX activities in chickpea may be related to the induction of antioxidant responses that protect the plant from oxidative damage. Among antioxidant enzymes, SOD converts the toxic O2¯ to H2O2 which must be scavenged to the O2 and H2O by the antioxidant enzymes such as CAT, POX and APX.
Superoxide dismutase (SOD) activity increase in all chickpea genotypes under drought condition. The higher SOD activity was noticed at post-anthesis stage than pre-anthesis stage. As for as genotype was concern maximum in ICC 4958 [243.59 units mg-1(protein) min-1] and it was minimum in DCP 92-3 [124.05 units mg-1(protein) min-1] (Fig. 4). Superoxide dismutase constitutes the first line of defence via detoxification of superoxide radicals (Sairam and Saxena, 2000), thereby maintaining membranes of plant tissue.
Catalase (CAT) eliminates H2O2 by breaking it down directly to water and oxygen. In our study, total CAT activity was enhanced by drought in the leaves of chickpea. Drought leads to significant increase in CAT activity.
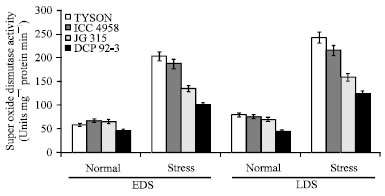 | |
| Fig. 4: | Effect of drought stress on superoxide dismutase (SOD) activity in four chickpea (C. arietinum L.) genotypes |
 | |
| Fig. 5: | Effect of drought stress on catalase (CAT) activity in four chickpea (C. arietinum L.) genotypes |
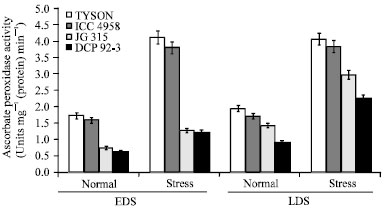 | |
| Fig. 6: | Effect of drought stress on ascorbate peroxidase (APX) activity in four chickpea (C. arietinum L.) genotypes |
Maximum CAT activity under drought stress noticed in genotype ICC 4958 (59.45 Units mg-1 (protein) min-1) while minimum in JG (31.44 Units mg-1 (protein) min-1) (Fig. 5). Catalase and peroxidases are the most important enzymes involved in regulation of intracellular level of H2O2 (Prasad et al., 1995). APX convert H2O2 into H2O along with the regeneration of NADP+, therefore, the enzymes play important roles in stress conditions (Jimenez et al., 1998). When one considers that CAT acts in peroxisomes, it is logical to suggest that photorespiration in the leaves of chickpea was also affected by drought treatment. Omidi (2010) recently noticed that enzyme activities of Ascorbate Peroxidase (APX) and Catalase (CAT) were highly significant difference between drought and genotypes in respect to CAT and APX. These were increase under drought stress.
APX which is primarily located both in chloroplasts and cytosol acts as a key enzyme of the glutathione-ascorbate pathway. It scavenges peroxides by converting ascorbic acid to dehydroascorbate and it is one of the most important enzymes playing a vital role in eliminating toxic H2O2 from plant cell in Asada-Halliwell pathway. In the present study, drought stress increased total APX activity differed prominently in stressed leaves as compared to that of non-stressed leaves. Higher activity of APX in Tyson and ICC 4958 under stressed leaves, generally its activity was 2-fold more over control, suggests a more effective H2O2 removal which might be produced by an enhanced activity of SOD in chickpea (Fig. 6).
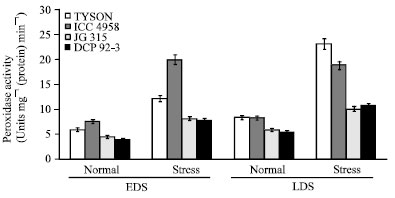 | |
| Fig. 7: | Effect of drought stress on peroxidase (POX) activity in four chickpea (C. arietinum L.) genotypes |
This well elucidates that POX should play a more significant role than CAT in detoxifying the produced H2O2 since the activity of POX increased, in contrast to that of CAT (Dey et al., 2007). It is well documented that CAT is less efficient than POX in scavenging of H2O2 because of its low substrate affinity (Erdal and Dumlupinar, 2010). Highest values of POX activity were observed in Tyson [22.88 unit mg-1(protein) min-1] followed by ICC 4958 [18.66 unit mg-1(protein) min-1] (Fig. 7). In tolerant genotype, POX activity was found to be higher, providing protection against the oxidative stress. Remarkably higher levels of POX in drought-stressed seedlings of chickpea might also be considered as a higher capacity to decompose H2O2 more rapidly.
CONCLUSION
Based on analyses of four chickpea genotypes and there were substantial variation in tolerance to drought within chickpea genotypes. A perusal of the results shows that drought tolerance genotypes have higher enzymatic activities such as SOD, POX, CAT, APX and lower level of MDA and H2O2 in comparison to drought susceptible genotypes. These parameters showed a considerable variability and heritability under drought stress conditions. Based on the significant and well defined relationships between lipid peroxidation and antioxidant enzyme activities, we concluded that these parameters could be instrumental in predicting the drought tolerance of chickpea genotypes. It is also concluded that pre- anthesis stage was most sensitive whereas, genotype ICC 4958 was most tolerant to drought.
Meagre information is available regarding genotypic variation for drought tolerance in chickpea. Henceforth, this study could help to understand some adaptive mechanisms developed by chickpea genotypes in contributing useful identified traits for chickpea breeding programme.
ACKNOWLEDGMENTS
We extend our sincere thanks to the University Grant Commission (UGC) for research fellowship for Ph.D. Programme and the Indian Institute of Pulse Research (IIPR- ICAR) Kanpur, India for their kind providing of chickpea genotypes through material transfer agreement (MTA).
REFERENCES
- Saleh, A.A.H., D.Z. Abdel-Kader and A.M. El Elish, 2007. Role of heat shock and salicylic acid in antioxidant homeostasis in Mungbean (Vigna radiata L.) plant subjected to heat stress. Am. J. Plant Physiol., 2: 344-355.
CrossRefDirect Link - Baisak, R., D. Rana, P.B.B. Acharya and M. Kar, 1994. Alterations in the activities of active oxygen scavenging enzymes of wheat leaves subjected to water stress. Plant Cell Physiol., 35: 489-495.
Direct Link - Davey, M.W., E. Stals, B. Pains, Keulemans and R.L. Swennen, 2005. High-throughput determination of malondialdehyde in plant tissues. Anal. Biochem., 347: 201-207.
CrossRefDirect Link - Davies, S.L., N.C. Turner, K.H.M. Siddique, J.A. Plummer and L. Leport, 1999. Seed growth of desi and kabuli chickpea (Cicer Arietinum) in a short season Mediterranean-type environment. Aust. J. Exp. Agric., 39: 181-188.
Direct Link - Dey, S.K., J. Dey, S. Patra and D. Pothal, 2007. Changes in the antioxidative enzyme activities and lipid peroxidation in wheat seedling exposed to cadmium and lead stress. Braz. J. Plant Physiol., 19: 53-60.
Direct Link - Dhindsa, R.S., P. Plumb-Dhindsa and T.A. Thorpe, 1981. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation and decreased levels of superoxide dismutase and catalase. J. Exp. Bot., 32: 93-101.
CrossRefDirect Link - Esfandiari, E., M.R. Shakiba, S.A. Mahboob, H. Alyari and S. Shahabivand, 2008. The effect of water stress on the antioxidant content, protective enzyme activities, proline content and lipid peroxidation in wheat seedling. Pak. J. Biol. Sci., 11: 1916-1922.
CrossRefPubMedDirect Link - Esterbauer, H. and K.H. Cheeseman, 1990. Determination of aldehydic lipid peroxidation products: Malonaldehyde and 4-hydroxynonenal. Methods Enzymol., 186: 407-421.
CrossRefPubMedDirect Link - Eyidogan, F. and M.T. Oz, 2007. Effect of salinity on antioxidant responses of chickpea seedlings. Acta Physiol. Plant, 29: 485-493.
CrossRef - Farooq, M., A. Wahid, N. Kobayashi, D. Fujita and S.M.A. Basra, 2009. Plant drought stress: Effects, mechanisms and management. Agron. Sustainable Dev., 29: 185-212.
CrossRefDirect Link - Farooq, M., A. Wahid, O. Ito, D.J. Lee and K.H.M. Siddique, 2009. Advances in drought resistance of rice. Crit. Rev. Plant Sci., 28: 199-217.
CrossRef - Foyer, C.H., H. Lopez-Delgado, J.F. Dat and I.M. Scot, 1997. Hydrogen peroxide- and glutathione-associated mechanisms of acclimatory stress tolerance and signalling. Physiol. Plant., 100: 241-254.
CrossRefDirect Link - Hasegawa, P.M., R.A. Bressan, J.K. Zhu and H.J. Bohnert, 2000. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol., 51: 463-499.
CrossRefDirect Link - Omidi, H., 2010. Changes of proline content and activity of antioxidative enzymes in two canola genotype under drought stress. Am. J. Plant Physiol., 5: 338-349.
CrossRefDirect Link - Hodges, D.M., J.M. DeLong, C.F. Forney and R.K. Prange, 1999. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta, 207: 604-611.
CrossRefDirect Link - Jimenez, A., J.A. Hernandez, G. Pastori, L.A. del Rio and F. Sevilla, 1998. Role of the ascorbate-glutathione cycle of mitochondria and peroxisomes in the senescence of pea leaves. Plant Physiol., 118: 1327-1335.
Direct Link - Kar, M. and D. Mishra, 1976. Catalase, peroxidase and polyphenoloxidase activities during rice leaf senescence. Plant Physiol., 57: 315-319.
CrossRefDirect Link - Lin, K.H.R., C.C. Tsou, S.Y. Hwang, L.F.O. Chen and H.F. Lo, 2006. Paclobutrazol pre-treatment enhanced flooding tolerance of sweet potato. J. Plant Physiol., 7: 750-760.
CrossRefDirect Link - Menconi, M., C.L.M. Sgherri, C. Pinzino and F. Navari-Izzo, 1995. Activated oxygen production and detoxification in wheat plants subjected to a water deficit programme. J. Exp. Bot., 46: 1123-1130.
CrossRef - Mittler, R., 2002. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci., 7: 405-410.
CrossRefPubMedDirect Link - Moinuddin and P. Imas, 2007. Evaluation of potassium compared to other osmolytes in relation to osmotic adjustment and drought tolerance of chickpea under water deficit environments. J. Plant Nutr., 30: 517-535.
CrossRefDirect Link - Møller, I.M., P.E. Jensen and A. Hansson, 2007. Oxidative modifications to cellular components in plants. Annu. Rev. Plant Biol., 58: 459-481.
CrossRefPubMedDirect Link - Mukherjee, S.P. and M.A. Choudhuri, 1983. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Plant Physiol., 58: 166-170.
CrossRefDirect Link - Munne-Bosch, S. and J. Penuelas, 2003. Photo and antioxidative protection and a role for salicylic acid during drought and recovery in field-grown Phillyrea angustifolia plants. Planta, 217: 758-766.
PubMed - Nakano, Y. and K. Asada, 1980. Spinach chloroplasts scavenge hydrogen peroxide on illumination. Plant Cell Physiol., 21: 1295-1307.
Direct Link - Nayyar, H., T.S. Bains and S. Kumar, 2005. Chilling stressed chickpea seedlings: Effect of cold acclimation, calcium and abscisic acid on cryoprotective solutes and oxidative damage. Environ. Exp. Bot., 54: 275-285.
CrossRefDirect Link - Tatar, O. and M.N. Gevrek, 2008. Influence of water stress on proline accumulation, lipid peroxidation and water content of wheat. Asian J. Plant Sci., 7: 409-412.
CrossRefDirect Link - Prasad, T.K., M.D. Anderson and C.R. Stewart, 1995. Localization and characterization of peroxidases in the mitochondria of chilling acclimated maize seedlings. Plant Physiol., 108: 1597-1605.
Direct Link - Prochazkova, D., R.K. Sairam, G.C. Srivastava and D.V. Singh, 2001. Oxidative stress and antioxidant activity as the basis of senescence in maize leaves. Plant Sci., 161: 765-771.
CrossRefDirect Link - Sairam, R.K. and D.C. Saxena, 2000. Oxidative stress and antioxidants in wheat genotypes: Possible mechanism of water stress tolerance. J. Agron. Crop Sci., 184: 55-61.
CrossRefDirect Link - Sairam, R.K., P.S. Deshmukh and D.C. Saxena, 1998. Role of antioxidant systems in wheat genotypes tolerance to water stress. Biologia Plantarum, 41: 387-394.
CrossRefDirect Link - Turner, N.C., G.C. Wright and K.H.M. Siddique, 2001. Adaptation of grain legumes (pulses) to water-limited environments. Adv. Agron., 71: 193-231.
CrossRefDirect Link