
Nasraoui-Hajjaji Afef
Unite de Recherche Nutrition et Metabolisme Azotes et Proteines de Stress, 99UR/09-20, Departement de Biologie, Faculte des Sciences de Tunis, Universite de Tunis Elmanar, 1060 Tunis, Tunisia
Saafi Leila
Unite de Recherche Nutrition et Metabolisme Azotes et Proteines de Stress, 99UR/09-20, Departement de Biologie, Faculte des Sciences de Tunis, Universite de Tunis Elmanar, 1060 Tunis, Tunisia
Bouthour Donia
Unite de Recherche Nutrition et Metabolisme Azotes et Proteines de Stress, 99UR/09-20, Departement de Biologie, Faculte des Sciences de Tunis, Universite de Tunis Elmanar, 1060 Tunis, Tunisia
Gouia Houda
Unite de Recherche Nutrition et Metabolisme Azotes et Proteines de Stress, 99UR/09-20, Departement de Biologie, Faculte des Sciences de Tunis, Universite de Tunis Elmanar, 1060 Tunis, Tunisia
Chaffei-Haouari Chiraz
Unite de Recherche Nutrition et Metabolisme Azotes et Proteines de Stress, 99UR/09-20, Departement de Biologie, Faculte des Sciences de Tunis, Universite de Tunis Elmanar, 1060 Tunis, Tunisia
American Journal of Plant Physiology
Year: 2011 | Volume: 6 | Issue: 6 | Page No.: 294-303







ABSTRACT
Cadmium (Cd) is a non-essential heavy metal that does not have any metabolic use. In this study the impacts of Cd added in nutrient solution on growth, physiology and carbon metabolism of tobacco (Nicotiana rustica) were investigated. Tobacco plants were grown in media containing CdCl2 at 0, 10, 20, 50 or 100 μM for 30 days. Regarding leaves, Dry Weight (DW) and leaf surface area were negatively affected by Cd. Soluble sugar content was decreased parallel with a reduce of ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBP) activity. The reduction in RuBP activity was accompanied by a decrease in Rubisco subunit amounts. These changes came with a dramatic loss in photosynthetic pigments. In another way, Cd stress enhanced the activities of phosphoenolpyruvate carboxylase (PEPC, EC 4.1.1.31) and NADP-isocitrate dehydrogenase (NADP-ICDH, EC 1.1.1.42) in tobacco leaves. This study suggested that readjustment of the C metabolism via modulation of Krebs cycle enzymes activities and C flow towards the amino acid biosynthesis were a strategy of tobacco plants to tolerate Cd stress.
PDF Abstract XML References Citation
Received: June 22, 2011;
Accepted: September 30, 2011;
Published: December 03, 2011
How to cite this article
Nasraoui-Hajjaji Afef, Saafi Leila, Bouthour Donia, Gouia Houda and Chaffei-Haouari Chiraz, 2011. Relationship Between Physiological and Biochemical Effects of Cadmium Toxicity in Nicotiana rustica. American Journal of Plant Physiology, 6: 294-303.
DOI: 10.3923/ajpp.2011.294.303
URL: https://scialert.net/abstract/?doi=ajpp.2011.294.303
DOI: 10.3923/ajpp.2011.294.303
URL: https://scialert.net/abstract/?doi=ajpp.2011.294.303
INTRODUCTION
Despite the importance of the type of abiotic stress, the effects of Cd contamination have undergone little study in certain plants of agricultural interest. Most of the information available about Cd physiology in plants comes from studies with the Cd-hyper accumulator Thlapsi caerulescens (Lombi et al., 2002) and Cd-tolerant plants such as Arabidopsis halleri, (Weber et al., 2006; Zhao et al., 2006) whereas less information is available in commercial crops. This is the case for tobacco plants. Tobacco (Nicotiana tabaccum and Nicotiana rustica) is a perennial herbaceous plant used primarily for commercial production of cigarettes and related products (Matt et al., 2002; Fu et al., 2010). Despite their economic importance few studies threat the quantitative response of growth. Regarding cadmium tobacco, can be considered as another origin of heavy metal contamination of the environment by Cd (McLaughlin and Singh, 1999).
It is generally admitted that growth of tobacco plants was inhibited by Cd (Dguimi et al., 2009). The reduction of biomass production by Cd could be related, at least in part, to photosynthesis impair. It can enter chloroplasts and disturb chloroplast function by inhibiting the enzymatic activities of chlorophyll biosynthesis, pigment-protein complexes (Scheible et al., 2000; Saleh, 2007). It is agreed that Tobacco leaves could accumulate Cd in the apoplast, by ionic interactions with carboxyl and/or sulphydryl groups from components of the cell wall (Vogeli-Lange and Wagner, 1996). Part of the metal could be complexed by phytochelatins or other ligands and sequestered in vacuole (Cho et al., 2003; Nouairi et al., 2006; Elhammadi et al., 2006).
One of the common changes noted in plants exposed to cadmium stress is the modulation of carbon metabolism enzymes (Gouia et al., 2003; Chiraz et al., 2008). Phosphoenolpyruvate carboxylase (PEPC) plays an anaplerotic role in the provision of C skeletons for amino acid synthesis. The increase in PEPC activity enhances the C flow through the anaplerotic pathways by providing C skeletons for the tricarboxylic acid cycle and for amino acid synthesis.
It is known that the activities of PEPC and ICDH, enzyme responsible of L-ketoglutarate synthesis, co-ordinate C and N metabolism (Scheible et al., 2000). Ribulose 1,5-bisphosphate carboxylase/oxygenase (Rubisco), a bifunctional enzyme with the capacity to competitively use CO2 or O2, is the key enzyme responsible for overall CO2 fixation during photosynthesis. Plants exposed to stress exhibit enhanced photorespiration that is believed to be a waste full process involved in release of CO2 (Azam and Farooq, 2003; Bota et al., 2004).
This study reports physiological and biochemical responses in leaves of Tobacco plants subjected to increasing concentrations of CdCl2. In this context, the first objective was to describe growth, determine photosynthesis pigments, ammonium and soluble sugar contents. The second aim was to investigate modulations of Rubisco, PEPC and NADPH-ICDH activities.
MATERIALS AND METHODS
Plant material and growth conditions: The study was derived a long 2009/2010 and 2010/2011 using tobacco (Nicotiana rustica, souffi, v). After imbibition, the seeds were germinated on moistened filter paper at 25°C in the dark. After 12 to 15 days, the uniform seedlings were transferred to plastic beakers (3 to 4 plants per beaker) filled with continuously aerated, control nutrient solutions, containing 8 mM KNO3, 1 mM Ca(NO3)2, 1 mM KH2PO4, 0,5 mM MgSO4, 32,9 Fe-EDTA, 30 mM H3BO3, 5 mM MnSO4, 1 mM CuSO4, 1 mM ZnSO4 and 1 mM (NH4)6MO7O24. Plants were grown in a growth chamber, (26°C/70% relative humidity during the day, 20°C/90% relative humidity during the night). The photoperiod was 16 h with a light irradiance of 150 μmol m-2 sec at the canopy level. At the age of 30 days plants were transferred to media containing CdCl2 at 0, 10, 20, 50 or 100 μM. Harvest was realised after one week of Cd treatment.
Determination of leaf area: The fresh leaves were collected and were scanned in order to determine their area.
Chlorophyll determination: Chla, Chlb and total Chl contents were determined by the method of Arnon (1949). The absorbance of a sample was read at 645, 651 and 663 nm and then contents were calculated.
Cadmium content: Cadmium content in foliar tissue was analyzed by digestion of dried samples with an acid mixture (HNO3/HClO, 4/1 v/v). Cadmium concentration was determined by atomic absorption spectrophotometry (Perkin-Elmer, Analyst 300).
Ammonium content: Ammonium was extracted from plant material at 4°C with 0.3 mM H2SO4 and 0.5% (w/v) polyclar AT. Ammonium content was quantified according to the reaction of Berthelot modified by Weatherburn (1967).
Soluble sugar content: The content of total soluble carbohydrates in the studied samples was determined according to McCready et al. (1950) using glucose as a standard. The absorption was determined by spectrophotometry at 640 nm.
Enzymatic assay: Enzymes were extracted from frozen leaves materials stored at -80°C. All extractions were performed at 4°C.
Ribulose 1,5-bisphosphate carboxylase/oxygenase: RuBP was extracted at 4°C by homogenizing frozen leaves samples in extraction buffer (50 mM Tris-HCl, (pH 8.5). Reaction was started after incubation at 30°C by the addition of 0.2 mM RuBP. The activity (as μmol NADH oxided. g-1 FW. min-1) was assayed spectrophotometrically at 340 nm according to Pehu et al. (1990).
Phosphoenolpyruvate carboxylase activity: PEPC activity was measured spectrophotometrically at 340 nm, in a final volume of 1 mL containing 100 mM Tris-HCl (pH 8.0), 3.5 mM MgCl2, 5 mM NaF and 0.2 mM NADH, at the optimal (pH 8.0) and at 4 mM PEP (Foyer et al., 1994). Assays were initiated by addition of the plant extracts.
Isocitrate dehydrogenase activity: Total ICDH activity was measured in 1 mL reaction set containing 0.1 mM potassium phosphate buffer (pH 7.6), 50 mM MgCl2, 50 mM isocitrate, 5 mM NADP+ and appropriate amount of crude extract. The reaction was initiated by adding isocitrate and the NADPH formed was followed at 340 nm (Galvez and Gadal, 1994).
Gel electrophoresis: Proteins were extracted as described above for Rubisco assay. Proteins were separated by SDS-PAGE (Laemmli, 1970). An equal amount of protein was loaded in each track.
Statistical analysis: Analyses of variance (ANOVA) with orthogonal contrasts and mean comparison procedures were used to detect differences between treatments. Mean separation procedures were carried out using the multiple range tests with Fisher’s least significant difference (LSD) (p<0.05).
RESULTS
Growth response to cadmium: Cadmium treatment leads to a progressive decrease in DW production (Fig. 1a). This reduction was more important at 100 μM Cd treatment. The leaf surface area was gradually decreased with increasing cadmium concentration in the nutrient medium (Fig. 1b).
The decrease of DW production and surface area in leaves was associated with a reduction of Chl a, Chl b and total Chl contents. In plants stressed with 100 μM Cd tenors of Chla, Chlb and total Chl are respectively 0.00725, 0.0715 and 0.0145 mg/gFW (Fig. 1c).
Cd, NH4+ and soluble sugar contents: Cadmium ion accumulated in leaves raised with increasing cadmium concentration in culture medium (Fig. 2a).
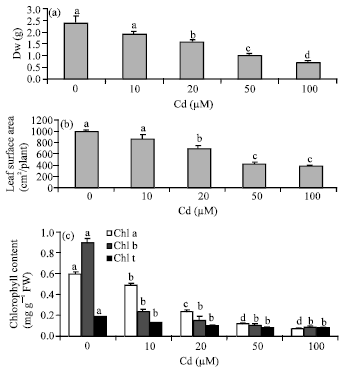 | |
| Fig. 1(a-c): | Changes in (a) Dry weight, (b) surface leaf area and (c) chlorophyll a, chlorophyll b and total contents in leaves of tobacco plants submitted to increasing dose of Cd. Values are means of five replications±SE. Data followed by different letters in bars are significantly different at p<0.05 |
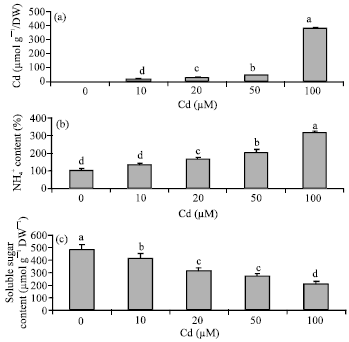 | |
| Fig. 2(a-c): | (a) Cadmium, (b) ammonium and (c) soluble sugar contents in leaves of Tobacco plants submitted to increasing dose of Cd. Values are means of five replications±SE. Data followed by different letters in bars are significantly different at p<0.05 |
Ammonium contents in the leaves increased at 50 μM Cd treatments. At 100 μM Cd, ammonium contents amounted by 200% relative to control (Fig. 2b).
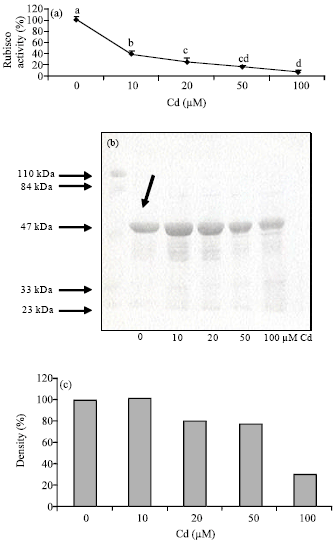 | |
| Fig. 3(a-c): | Changes in (a) RuBP activity gel, (b) electrophoresis and (c) density of RuBP protein in leaves of Tobacco plants submitted to increasing dose of Cd. Results were expressed in % of the control (0.69±0.658 μmol/g FW/min). Values are means of five replications±SE. Data followed by different letters in a graph line are significantly different at p<0.05 |
The measurement of soluble sugar content in leaves of tobacco showed that after Cd treatment, an important reduction of the soluble sugar content was detected under 10 μM Cd (Fig. 2c). The reduction was accentuated reaching approximately 80% at 100 μM Cd.
Effects of CdCl2 on several carbon metabolism enzymes: The presence of Cd in the culture medium induced a decrease of RuBP activity in tobacco leaves (Fig. 3a). RuBP activity was considerably reduced, reaching an annulation of its activity in leaves treated with 100 μM of Cd (Fig. 3a). Lose of activity was campaigned by diminution of RuBP protein amount (Fig. 3b). Density of each band was detected and demonstrated in Fig. 3c.
PEPC activity changed with Cd supply in culture medium in leaves Tobacco (Fig. 4a). A significant stimulation of PEPC activity was observed in leaves 10 times of the control at 100 μM Cd.
The effect of cadmium treatment on ICDH activity is presented in Fig. 4b which shows that exogenously applied Cd clearly stimulated NADP+-ICDH activity in leaves.
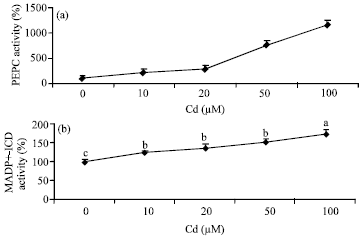 | |
| Fig. 4(a-b): | (a) Changes in PEPC and (b) ICDH activity in leaves of Tobacco plants submitted to increasing dose of Cd. PEPc and ICDH activity were expressed respectively in % of control (0.089± 0.042 μmol/g FW/min; 0.0785±0.0068 μmol/gFW/min. Values are means of five replications±SE. Data followed by different letters in a graph line are significantly different at p<0.05 |
DISCUSSION
It is well known that Cd inhibits plant growth; this is consistent with the results presented in this study. Such us shown in many other studies realized with different plant species, cadmium provoked a considerable reduction in growth of tobacco seedlings accompanied by a decline in leaf surface area (Shaukat et al., 1999; Azmat et al., 2005; El-Shora and Ali, 2011). One recognized explanation of the impact of heavy metals on the plant physiology is that its results in several nutritional disturbances and in chlorophyll contents. Data presented in this work demonstrated a clear decline of photosynthetic pigments tenors. This is in agreement with previous results demonstrating that in major plant species, cadmium stress led to a leaf yellowing related to chlorophyll breakdown (Diaz et al., 2006). This decrease could cause in part a photosynthesis and growth reduction (Singh et al., 2008). This correlation was well documented and shown in several species such as soybean (Krupa, 1999), bean (Ismail, 2008) and tomato (Nasraoui-Hajaji et al., 2010). Regarding Tobacco plants, the toxic effects of Cd on growth and development are well demonstrated (Bi et al., 2009; Dguimi et al., 2009).
In this article, cadmium effects on the processes related to growth inhibition is reviewed. Photosynthesis, photorespiration and the photo energy consumption and reductant supply were indirectly studied via investigation of the principal enzymes of carbon metabolism.
Data shown in the present study were in agreement with the idea that under cadmium stress, CO2 assimilation rate is limited since the amount and activity of Rubisco was decreased. More that, RuBPCase activity is regulated to maintain a balance between the capacities of the photosynthetic apparatus to produce and consume RuBP and triose phosphates (Stiborova, 1988). Targeting the accumulation of Cd in leaves enabled us to investigate its effects on the most abundant enzyme, Rubisco. Both the carboxylase and the oxygenase activities of Rubisco are known to be susceptible to abiotic stresses. It was clearly evident that once the onset of decline in the activity of Rubisco coupled with the decrease in protein enzyme amount, became evident. Since in the leaf, Rubisco dominates the C3 and C4 protein species profile, exhibiting roughly 50% and 30% of the total protein respectively (Feller et al., 2008). Decrease in Rubisco activity was accompanied by declined level of soluble sugars suggests that under in situ cadmium stress sugar synthesis is reduced, relative to CO2 fixation capacity of Rubisco (Leitao et al., 2003). Different observations thus suggest that cadmium causes a marked alteration in the behaviour of sugar metabolising enzymes leading to a starvation of total soluble sugar. Such impairment in carbohydrate metabolism would influence photosynthate partitioning and would ultimately lead to impaired growth of tobacco seedling in Cd polluted soils.
Since photorespiration and Rubisco activity diminished, it is suggests that ammonia accumulation in leaves cannot derive from photorespiratory recycling. Thus, we supposed that accumulation of ammonia in Cd-treated plants is rather a consequence of protein proteolysis and amino-acid hydrolysis. Therefore, the ability of cells to switch from one development state to another or to adapt the new environmental condition often requires the rapid dismantlement of existing regulatory networks through proteolysis. In addition, Cd stress induced PEPC activity and the induction was partially due to PEPC protein synthesis. This PEPC activity showed a marked increase to about 4-10 times of the control at 100 μM in leaves. PEPC is the primary cytosolic enzyme replenishing oxaloacetate in the tricarboxylic acid cycle (Champigny and Foyer, 1992). Carbon skeletons provided by sugar mobilization from leaves is consumed by the mitochondria phosphorylation pathway in roots.
In Cd-treated leaves we observed that also ICDH activity was increased. This suggests that the synthesis of 2-oxoglutarate allowed to amino-acid synthesis and peculiarly to glutamate maintenance continue and that the availability of reducing power remained however sufficient (Gouia et al., 2003; Smeets et al., 2005; Chaffei-Haouari et al., 2009). ICDH activity was generally inhibited by the other heavy metals such as Zn (Kasim, 2007).
The capacity of plant to provide carbon skeletons through carbon fixation by plant to assimilate ammonium PEPC is considered to be an important enzyme involved in the strategy of plant to tolerate cadmium stress and ammonium toxicity.
At this state we can suggest that under stress condition, Rubisco and PEPC activities result from a complex interplay of stomatal and non-stomatal responses to Cd presence. More that, carbon metabolism is an important process which have a range of functions signaling and defense against abiotic stress.
CONCLUSION
Data presented in this paper showed that after Cd treatment, PEPC and ICDH also increased in Cd treated tobacco plants suggesting that they divert C away from malate synthesis towards 2-oxoglutarate production in favour of ammonium assimilation into glutamate. This study added further support to the hypothesis that the extent of Cd treatment moderation of several metabolic pathways, including carbon fixation via PEPC and Rubisco.
This late metabolic adjustment seems to correlate with the growth inhibition. However, increase in PEPC activity could give rise to the amino acids required to allow the production of anti-oxidant enzymes and all other stress related proteins which support the hypersensitive response of tobacco leaves to cadmium.
REFERENCES
- Arnon, D.I., 1949. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol., 24: 1-15.
CrossRefPubMedDirect Link - Azam, F. and S. Farooq, 2003. Elevated CO2 and stress tolerance in crop plants with particular reference to agro-climatic conditions of Pakistan. Pak. J. Biol. Sci., 6: 1096-1107.
CrossRefDirect Link - Azmat, R., Y. Akhtar, R. Talat and F. Uddin, 2005. The inhibition of bean plant metabolism by CD metal and atrazine: I. The effect of atrazine with CD metal on growth, photosynthesis, nutritional level and rhizosphere of soil. Biotechnology, 4: 238-242.
CrossRefDirect Link - Bi, Y., W. Chen, W. Zhang, Q. Zhou, L. Yun and D. Xing, 2009. Production of reactive oxygen species, impairment of photosynthetic function and dynamic changes in mitochondria are early events in cadmium-induced cell death in Arabidopsis thaliana. Biol. Cell, 101: 629-643.
PubMed - Bota, J., H. Medrano and J. Flexas, 2004. Is photosynthesis limited by decreased Rubisco activity and RuBP content under progressive water stress? New Phytol., 162: 671-681.
CrossRefDirect Link - Chaffei-Haouari, C., E. Carrayol, M.H. Ghorbel and H. Gouia, 2009. Physiological and biochemical effects of cadmium toxicity in enzymes involved in nitrogen and amino-acid metabolism in tomato plants. Acta Bot. Gallica, 156: 477-486.
Direct Link - Chiraz, C., G. Houda, D. Mohamed and G.M. Habib, 2008. Differential toxicological response to cadmium stress of bean seedlings grown with NO3- or NH4+ as nitrogen source. Int. J. Bot., 4: 14-23.
CrossRefDirect Link - Champigny, M. and C. Foyer, 1992. Nitrate activation of cytosolic protein kinases diverts photosynthetic carbon from sucrose to amino acid biosynthesis. Plant Physiol., 100: 7-12.
Direct Link - Cho, M., A.N. Chardonnens and K.J. Dietz, 2003. Differential heavy metal tolerance of Arabidopsis halleri and Arabidopsis thaliana: A leaf slice test. New Phytol., 158: 287-293.
Direct Link - Dguimi, H.M., M. Debouba, M.H. Ghorbel and H. Gouia, 2009. Tissue-specific cadmium accumulation and its effects on nitrogen metabolism in tobacco (Nicotiana tabaccum, Bureley v. Fb9). Com Rendu. Sci., Biol., 332: 58-68.
CrossRefDirect Link - Elhammadi, M.A., R. Chaffai, A. Tekitek and E. El-Ferjani, 2006. Analysis of cadmium-peptide complexes in sunflower (Helianthus annuus L.) roots as detected by gel-filtration chromatography. J. Plant Sci., 1: 201-211.
CrossRefDirect Link - El-Shora, H.E. and A.S. Ali, 2011. Changes in activities of nitrogen metabolism enzymes in cadmium stressed marrow seedlings. Asian J. Plant Sci., 10: 117-124.
CrossRefDirect Link - Feller, U., I. Anders and T. Mae, 2008. Rubiscolytics: Fate of Rubisco after its enzymatic function in a cell is terminated. J. Exper. Bot., 59: 1615-1624.
CrossRef - Foyer, C.H., G. Noctor, M. Lelandai, J.C. Lescure, M.H. Valadier, J.P. Boutin and P. Horton, 1994. Short-term effects of nitrate, nitrite and ammonium assimilation on photosynthesis, carbon partitioning and protein phosphorylation in maize. Planta, 192: 211-220.
CrossRefDirect Link - Fu, H., P.A. Machado, T.S. Hahm, R.J. Kratochvil, C.I. Wei and Y.M. Lo, 2010. Recovery of nicotine-free proteins from tobacco leaves using phosphate buffer system under controlled conditions. Bioresour. Technol., 101: 2034-2042.
CrossRef - Galvez, S. and P. Gadal, 1994. On the function of the NADP-dependent isocitrate dehydrogenase isoenzymes in living organisms. Plant Sci., 105: 1-14.
CrossRef - Gouia, H., A. Suzuki, J. Brulfert and M.H. Ghorbal, 2003. Effects of cadmium on the co-ordination of nitrogen and carbon metabolism in bean seedlings. J. Plant Physiol., 160: 367-376.
CrossRef - Ismail, M.A., 2008. Involvement of Ca2+ in alleviation of Cd2+ toxicity in common bean (Phaseolas vulgaris L.) plants. Asian J. Biol. Sci., 1: 26-32.
CrossRefDirect Link - Krupa, Z., 1999. Cadmium against higher plant photosynthesis-a variety of effects and where do they possible come from. Zeitsschrift fuer Naturforschung, 54: 723-729.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Leitao, L., P. Goulas and J. Biolley, 2003. Time-course of Rubisco oxidation in beans (Phaseolus vulgaris L.) subjected to a long-term ozone stress. Plant Sci., 165: 613-620.
CrossRef - Lombi, E., K.L. Tearall, J.R. Howarth, F.J. Zhao, M.J. Hawkesford and S.P. McGrath, 2002. Influence of iron status on cadmium and zinc uptake by different ecotypes of the hyperaccumulator Thlaspi caerulescens. Plant Physiol., 128: 1359-1367.
CrossRef - McCready, R.M., J. Guggolz, V. Silviera and H.S. Owens, 1950. Determination of starch and amylose in vegetables. Anal. Chem., 22: 1156-1158.
CrossRefDirect Link - Matt, P., A. Krapp, V. Haake, H. Mock and M. Stitt, 2002. Decreased Rubisco activity leads to dramatic changes of nitrate metabolism, amino acid metabolism and the levels of phenylpropanoids and nicotine in tobacco antisense RBCS transformants. Plant J., 30: 663-677.
PubMed - Nasraoui-Hajaji, A., F. Gharbi, M.H. Ghorbel and H. Gouia, 2010. Cadmium stress effects on photosynthesis and PSII efficiency in tomato grown on NO3- or NH4+ as nitrogen source. Acta Bot. Gallica, 157: 101-115.
Direct Link - Nouairi, I., A.W. Ben, Y.N. Ben, M.D.D. Ben, G.M. Habib and Z. Mokhtar, 2006. Variations in membrane lipid metabolism in Brassica juncea and Brassica napus leaves as a response to cadmium exposure. J. Agron., 5: 299-307.
CrossRefDirect Link - Pehu, E., A. Nurmi and M.A.J. Parry, 1990. Variation in net photosynthesis, rubisco activity and chloroplast ultrastructure among somatic hybrids of Solanum tuberosum and S. brevidens. J. Exp. Bot., 41: 769-774.
CrossRef - Saleh, A.A.H., 2007. Influence of UVA+B radiation and heavy metals on growth, some metabolic activities and antioxidant system in pea (Pisum sativum) plant. Am. J. Plant Physiol., 2: 139-154.
CrossRefDirect Link - Scheible, W.R., A. Krapp and M. Stitt, 2000. Reciprocal diurnal changes of phosphoenolpyruvate carboxylase expression and cytosolic pyruvate kinase, citrate synthase and NADP-isocitrate dehydrogenase expression regulate organic acid metabolism during nitrate assimilation in tobacco leaves. Plant Cell Env., 23: 1155-1167.
CrossRef - Singh, S., N.A. Khan, R. Nazar and N.A. Anjum, 2008. Photosynthetic traits and activities of antioxidant enzymes in blackgram (Vigna mungo L. Hepper) under cadmium stress. Am. J. Plant Physiol., 3: 25-32.
CrossRefDirect Link - Smeets, K., A. Cuypers, A. Lambrechts, B. Semane, P. Hoet, A. Van Laere and J. Vangronsveld, 2005. Induction of oxidative stress and antioxidative mechanisms in Phaseolus vulgaris after Cd application. Plant Physiol. Biochem., 43: 437-444.
CrossRef - Shaukat, S.S., M. Mushtaq and Z.S. Siddiqui, 1999. Effect of cadmium, chromium and lead on seed germination, early seedling growth and phenolic contents of Parkinsonia aculeata L. and Pennisetum americanum (L.) Schumann. Pak. J. Biol. Sci., 2: 1307-1313.
CrossRefDirect Link - Weatherburn, M.W., 1967. Phenol-hypochlorite reaction for determination of ammonia. Anal. Chem., 39: 971-974.
CrossRefDirect Link - Weber, M., A. Trampczynska and S. Clemens, 2006. Comparative transcriptome analysis of toxic metal responses in Arabidopsis thaliana and the Cd2+-hypertolerant facultative metallophyte Arabidopsis halleri. Plant Cell Environ., 29: 950-963.
CrossRef - Kasim, W.A., 2007. Physiological consequences of structural and ultra-structural changes induced by Zn stress in Phaseolus vulgaris. II. enzymes, amino acids and protein profile. Int. J. Bot., 3: 33-39.
CrossRefDirect Link - Zhao, F.J., R.F. Jiang, S.J. Dunham and S.P. McGrath, 2006. Cadmium uptake, translocation and tolerance in the hyperaccumulator Arabidopsis halleri. New Phytol., 172: 646-654.
CrossRef