
M. Kazemi
Department of Horticulture Sciences, Faculty of Agriculture and Natural Resources, Islamic Azad University, Karaj Branch, Young Researchers Club, Karaj, Iran
M. Aran
Department of Landscape, College of Agriculture, University of Zabol, Zabol, Iran
S. Zamani
Department of Horticulture Sciences, Faculty of Agriculture and Natural Resources, Islamic Azad University, Karaj Branch, Young Researchers Club, Karaj, Iran
American Journal of Plant Physiology
Year: 2011 | Volume: 6 | Issue: 3 | Page No.: 167-175







ABSTRACT
Combinations of glutamine, succinic acid, Salicylic acid and citric acid were used as preservative mixture for cut Lisianthus and their effect on regulation of senescence was examined. In this study, the recorded traits included vase life, total chlorophyll content (SPAD reading), anthocyanin leakage, malondialdehyde content and ACC-oxidase activity. The vase were placed in chambers at 19°C, relative humidity about 70% and 14 h photoperiod that was maintained using fluorescent lamps (light intensity of 15 μmol m-2 sec-1) at the top of the corolla. The results showed that glutamine, succinic acid and Salicylic acid treatments increased the vase life and decrease the percentage of wilting compared to the control. The vase solution containing 3 mM glutamine and 4 mM succinic acid with 2 mM salicylic acid significantly increased vase life compared to the control, in addition, the malondialdehyde accumulation and ACC-Oxidase activity reduced in the same solution while membrane stability was improved. Results suggest that glutamine, succinic acid and Salicylic acid increases vase life by affecting many of the age-related changes associated with Lisianthus petal senescence.
PDF Abstract XML References Citation
Received: April 25, 2011;
Accepted: May 26, 2011;
Published: July 09, 2011
How to cite this article
M. Kazemi, M. Aran and S. Zamani, 2011. Extending the Vase Life of Lisianthus (Eustoma grandiflorum Mariachii. cv. blue) with Different Preservatives. American Journal of Plant Physiology, 6: 167-175.
DOI: 10.3923/ajpp.2011.167.175
URL: https://scialert.net/abstract/?doi=ajpp.2011.167.175
DOI: 10.3923/ajpp.2011.167.175
URL: https://scialert.net/abstract/?doi=ajpp.2011.167.175
INTRODUCTION
Lisianthus is now an important commercial cut flower; there have been many studies of the environmental conditions required for Lisianthus production (Cho et al., 2001). Vase life of cut Lisianthus flower is usually short. Short postharvest vase life is one of the most important problems of the cut flowers. Senescence of cut flowers is induced by several factors e.g., water stress (Sankat and Mujaffar, 1994), carbohydrate depletion, microorganisms (Witte and van Doom, 1991) and ethylene effects. Ethylene enhanced flower senescence and wilting of cut flowers (Hojjati et al., 2007), increased permeability of petal cells and accelerated the decrease in cell membrane fluidity (Reezi et al., 2009). The other consequences include increase in cell membrane permeability and solute uptake capacity, degradation of membrane lipids and MDA production. Ethylene production causes a sharp increase in production of oxygen free radicals (ROS) which is responsible for stress dependent peroxidation of membrane lipids (Reezi et al., 2009). One effect of Reactive Oxygen Species (ROS) accumulation in plant cells under stress is lipid peroxidation via oxidation of unsaturated fatty acids leading to membrane damage and electrolyte leakage (Yildirim et al., 2008). Rapid accumulation of free proline is also a typical response to stress and when plants are exposed to stress, many plants accumulate high amounts of proline, in some cases several times the sum of all the other amino acids (Mansour et al., 2000). The role of proline in protection of cell membranes against stress injury has been discussed by Mansour et al. (1998). To scavenge ROS, plants posses specific mechanisms which include activation of antioxidant enzymes (Jaleel et al., 2006) and non enzymatic antioxidants such as, carotenoids and ascorbic acid (Mittler, 2002). Salicylic Acid (SA) is considered as a hormone-like substance which plays an important role in regulating a number of physiological processes and provide protection against biotic and abiotic stresses in plant (Hussein et al., 2007). The protective function of SA includes the regulation of ROS and antioxidant enzymes (Khan et al., 2003; El-Tayeb et al., 2006; Ansari and Misra, 2007; Mba et al., 2007; Mahdavian et al., 2007; Canakci, 2008; Shi and Zhu, 2008). SA acid with increases the enzyme antioxidant activity cause delay, the onset of hydrolysis of structural cell components, decrease ROS production, ACC-oxidase activity and sensitivity (Li et al., 1992; Srivastava and Dwivedi, 2000). Succinic acid is found in all plant and animal materials as a result of the central metabolic role played by this dicarboxylic acid in the citric acid cycle. Succinate can solely as a catalytic system in carbohydrate respiration. The rapid oxidation of succinate was leading to the accumulation of malate. Succinate with increases Chlorophyll content cause delay in the onset of hydrolysis of structural cell components and sensitivity (Rahemi, 2005). Citric acid seems to act by reducing the pH of water and, consequently, the proliferation of bacteria which block the xylem vessels in the cut region and interfere with the normal flux of water through the stem (Nowak and Rudniki, 1990). The senescence of cut flowers is closely related to a considerable reduction of the energy needed for synthesis reactions. Therefore, an exogenous carbohydrate supplementation would be enough to delay the senescence, considering that the main effect would be to maintain the structure and activity of the mitochondria. Glutamine, a multifaceted amino acid used as an energy substrate for most cells. It is important as a constituent of proteins and as a central metabolite for amino acid transamination via α-ketoglutarate and glutamate. Glutamine plays an important role in the nitrogen and carbonskeleton exchange among different tissues, where this amino acid fulfils many different physiological functions. When glucose levels are low and energy demands are high, cells can metabolize amino acids for energy. Also, glutamine is one of the most readily available amino acids for use as an energy source and it is a major source of energy for many rapidly dividing cell types. Therefore, in this study, examined the preservative effects of salicylic acid, succinic acid, glutamine, citric acid and their interaction on the vase life of cut Lisianthus flowers with emphasis on the possibility of SA effect on antioxidative indicators of cut flower.
MATERIALS AND METHODS
Plant material: The experiment was started September 10, 2010 and chlorophyll content, Membrane stability, MDA content and ACC Oxidase activity were measured at 15th day of vase life. This study was on the effect of succinic acid, salicylic acid, citric acid and glutamine treatments on vase life of Lisianthus cut flowers, in a factorial test with complete randomized design with six replications. Cut flowers were harvested in half-open stage from local commercial greenhouses (Mahallat, arak, Iran), in the morning and transported with appropriate covers immediately. Cut flower stems of Lisianthus (Eustoma grandiflorum Mariachii. cv. blue) (40 cm in length) were placed in solution containing succinic acid (0, 2 and 4 mM), salicylic acid (0, 2 and 4 mM), citric acid (0 and 2 mM) and glutamine (0, 1.5 and 3 mM) after cutting. Six cut flowers were placed in a 300 mL bottle with 250 mL of solution. Distilled water was used for the controls and placed in chambers at 19°C. The relative humidity was about 70% while 14 h photoperiod was maintained using fluorescent lamps with a light intensity of 15 μmol m-2 sec-1 at the top of the corolla.
Vase life: The vase life of the inflorescence was considered terminated when 50% of the open flowers had wilted.
Chlorophyll content measurement: Total chlorophyll (a+b) content was measured by chlorophyll meter (SPAD-502, Minolta Co. Japan) which is presented by SPAD value. Average of 3 measurements from different spots of a single leaves was considered.
Determination of anthocyanin leakage: Anthocyanin leakage was measured based on the method of Poovaiah (1979).
Determination of Acc-oxidase activity: ACC oxidase activity was measured based on the method of Maye-Lean and John (1994).
Assays of MDA content (Lipid peroxidation): Oxidative damage to lipids was measured based on the method of Heath and Packer (1968).
Superoxide dismutase (SOD) activity: The activity of superoxide dismutase was measured based on the method of Beauchamp and Fridovich (1971).
Carbohydrates determination: Carbohydrates were measured based on the method of Hassan (2005).
Determination of proline: Proline was quantified by using ninhydrin reagent and measured according to Bates et al. (1973).
Experimental design and statistical analysis: Experiment was arranged in a factorial test with complete randomized design with six replications. Analysis of variance was performed on the data collected using the General Linear Model (GLM) procedure of the SPSS software )Version 16, IBM Inc.). The mean separation was conducted by tukey analysis in the same software (p = 0.05).
RESULTS
Anthocyanin leakage and ACO activity: The results indicate that anthocyanin leakage and ACO activity was not affected by citric acid treatment (p = 0.05). Treatment with 2 mM succinic acid +0 mM SA+3 mM glutamine solution slightly reduced the Anthocyanin leakage and ACO activity and extended vase life of the Lisianthus (Table 1). While the improvement in membrane stability with increased concentrations of SA and succinic acid solution, flowers treated with 4 mM succinic acid +2 mM SA and 3 mM glutamine solution significantly decreased Anthocyanin leakage and ACO activity (p = 0.05). Highest means of ACO activity was found in cut flowers treated with 4 mM SA (Table 1).
| Table 1: | Mean comparisons of chlorophyll content, vase life, MDA, SOD activity, membrane stability and ACC oxidase activity in succinic acid, SA and glutamin treatments and their interaction |
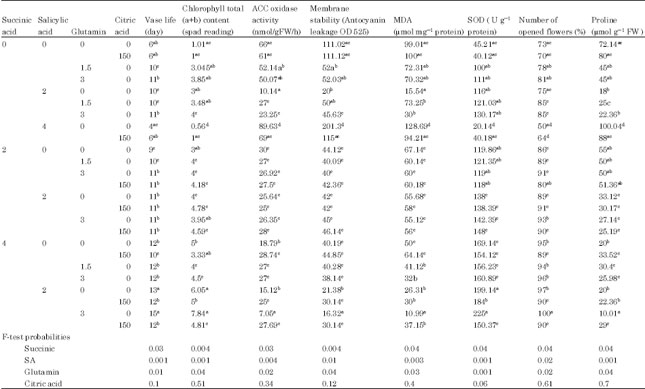 | |
| Means in each column followed by similar letters are not significantly different at 5% level | |
MDA content and superoxide dismutase activity: MDA accumulation was inhibited in the flowers of Lisianthus by succinic acid, Salicylic acid and glutamine to a larger extent when compared to control. The results indicate that MDA accumulation was not affected by citric acid treatment (p = 0.05). A significant negative correlation was observed between succinic acid concentrations and the MDA content in Lisianthus cut flowers (Table 1). Highest means of MDA accumulation was found in cut flowers treated with 4 mM SA (p = 0.05). Lisianthus flowers treated by SA alone or together with glutamine and succinic acid had more Superoxide dismutase activity. The maximum Superoxide dismutase activity was recorded in flowers treated by 4 mM succinic acid +2 mM SA+3 mM glutamine compared other treatments and control (p = 0.05). The minimum Superoxide dismutase activity was noted in 4 mM SA (Table 1). A significant positively correlation was observed between succinic acid concentrations and the Superoxide dismutase activity in Lisianthus cut flowers.
Total chlorophyll content: The results indicate that total chlorophyll content increased with increased succinic acid and glutamine concentration in Lisianthus cut flower (Table 1). Result showed that 4 mM succinic acid +2 mM SA+3 mM glutamine leaded to a considerable delay in degradation of chlorophyll compared to other concentrations (4 mM SA and control) (p = 0.05). Highest means of degradation was found in cut flowers treated with 4 mM SA (Table 1).
Carbohydrate content: Data of Table 2 show that fructose, glucose and sucrose were the main soluble carbohydrates in petals and stems of cut Lisianthus. glucose was the major component in the petals as well as in stems but, generally, its value was higher than in stems. Sucrose contents in petals and stems were lower than those of glucose (p = 0.05). The results indicate that the carbohydrate content significantly increased as a result of using 3 mM glutamine till the 6rd day then sharply decreased on the 15th day at which the vase life of control was terminated (p = 0.05) flowers treated with 1.5 and 3 mM glutamine solution significantly increased Carbohydrate Content in cut flowers (p = 0.05).
| Table 2: | Effect of glutamine with or without succinic and SA on carbohydrate content (in mg.1 dry weight) for petals and stems of Lisianthus cut flowers |
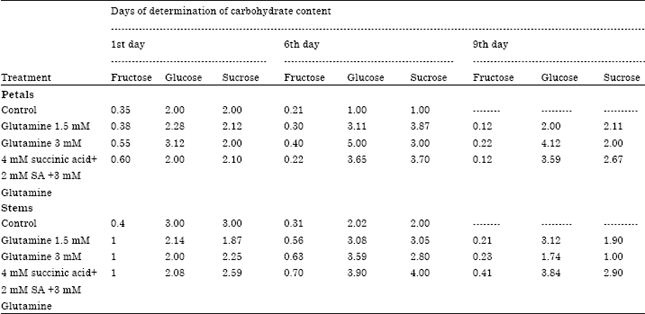 | |
The results indicate that a significant positive correlation was observed between glutamine concentrations and the carbohydrate content in cut flowers (Table 1).
Proline content: The results indicate that proline content in flower not change in condition which 2 mM succinic acid was used as treatment but when high concentration of succinic acid (4 mM) was used proline significantly increased (p = 0.05). Succinic acid, SA and glutamine treatments significantly decreased proline in flowers which were treated with 4mM succinic acid+2 mM SA+3 mM glutamine (Table 1).
Vase life: Holding Lisianthus cut flowers in vase solutions containing 4 mM succinic acid +2 mM SA+3 mM glutamine significantly increased their vase life and delayed flower senescence compared to flowers either held in 0 and 2 mM succinic acid or distilled water (p = 0.05). Succinic acid was found to be significantly and positive correlated with vase life of the Lisianthus cut flowers as well. In present experiment adding glutamine and SA to vase solutions containing succinic acid could increase the vase life of cut flowers compared to control (Table 1). As the glutamine content of the vase solution increased, the postharvest performance and quality of the flowers changed dramatically (Table 1). Also, the results indicate that the number of flowers opening increased significantly between 1.5 and 3 glutamine concentrations.
DISCUSSION
In the present experiments we used succinic acid, salicylic acid, citric acid and glutamine treatments as senescence retarding agent. While citric acid alone treatments did not have effects on Acc-oxidase and vase life cut flowers; succinic acid, glutamine and SA treatments reduced Acc-oxidase activity and increased vase life cut flowers. Present result showed that adding succinic acid in vase water significantly reduced Acc-oxidase activity, this result indicates that succinic acid inhibits the conversion of ACC to ethylene in Lisianthus petals. The above mentioned results are agreed with those recorded by Fan et al. (2008) and Yuping (2009), they found that SA was inhibit the conversion of ACC to ethylene in gebrera. In present experiment; succinic, SA and glutamine interaction reduced anthocyanin leakage, MDA and proline accumulation and increased total chlorophyll content and superoxide dismutase activity. Succinate with increases chlorophyll content cause delay the onset of hydrolysis of structural cell components and sensitivity (Rahemi, 2005). Present result showed that increased chlorophyll content in cut flower lisianthus was observed with increased Succinic acid concentration. The protective function of SA includes the regulation of ROS and antioxidant enzymes (Khan et al., 2003; Shi and Zhu, 2008). SA acid with increases the enzyme antioxidant activity cause delay the onset of hydrolysis of structural cell components, decrease ROS production and sensitivity. Other studies have shown that exogenous SA can regulate the activities of antioxidant enzymes and increase plant tolerance to abiotic stress (He et al., 2002). SA also decreased electrolyte leakage under stress (Karlidag et al., 2009). Present result was in agreement with the report of Zhou et al. (1999) that reported photosynthetic pigments were increased in corn with SA application. Khan et al. (2003) showed that SA increased photosynthetic rate in corn and soybean. Present result showed that increased enzyme antioxidant activity in cut flower Lisianthus was observed with increased SA application while treatment with succinic acid solution slightly increased the enzyme antioxidant activity. Presumably, succinic and SA were able to alter the sterol biosynthesis and changed the composition of sterol in the plasmamembrane. This change in sterol composition may induce changes in cell membrane that may be reflected in increased membrane stability and reduced MDA and proline accumulation in cut flowers. Proline in generally assumed to serve as a physiologically compatible solute that is increase needed to maintain a favorable osmotic potential between the cell and its surroundings (Pollard and Jones, 1979). The data showed that proline content in flower decreased as the SA increased. These results are in agreement with those of Mohammad et al. (1998) who found that SA decreses the proline content in the leaves of tomato. In the present study when SA was applied there was a reduction in proline accumulation Thus, proline can be considered to be one of the important factors involved in SA induced protective mechanism in flower in response to stress. When glucose levels are low and energy demands are high, cells can metabolize glutamine for energy. Glutamine is one of the most readily available amino acids for use as an energy source and it is a major source of energy for many rapidly dividing cell types. The opening of Lisianthus flowers is associated with substantially increased concentrations of glucose in the corolla. The glucose probably serves to provide osmotic potential for the expansion of the petal cells and the availability of soluble carbohydrate for that purpose is probably partly responsible for the improved opening of the flowers in preservative solution. Present result showed that the glucose in stems and petals content significantly increased as a result of using glutamine in vase solution. The addition of glutamine greatly improved opening and vase life of the opened flowers but citric acid alone provided no significant additional benefit. Therefore, in this study, use of succinic acid, SA and glutamine as a preservative mixture ingredient increased vase life of cut flowers significantly. The observed reduction in Anthocyanin leakage, Acc-oxidase activity and MDA and proline accumulation and increase vase life cut flowers by succinic acid, SA and glutamine application supports our conclusion of considering it as a practical agent for retarding of Lisianthus cut flower senescence.
CONCLUSION
From the results of the present study, it can be concluded that glutamine, succinic acid and Salicylic acid treatments significantly decrease produce MDA and ACC-oxidase activity, in vase flower preservative solution and reduce the membrane permeability and peroxidation of lipids compared to the control. glutamine, succinic acid and salicylic acid also proved more effective in delaying petal senescence and/or flower wilting. However, result our showed that glutamine, succinic acid and salicylic acid treatments maintained the vase life of flowers for a longer period.
REFERENCES
- Ansari, M.S. and N. Misra, 2007. Miraculous role of salicylic acid in plant and animal system. Am. J. Plant Physiol., 2: 51-58.
CrossRefDirect Link - Beauchamp, C. and I. Fridovich, 1971. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem., 44: 276-287.
CrossRefPubMedDirect Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Canakci, S., 2008. Effects of salicylic acid on fresh weight change, chlorophyll and protein amounts of radish (Raphanus sativus L.) seedlings. J. Biol. Sci., 8: 431-435.
CrossRefDirect Link - El-Tayeb, M.A., A.E. El-Enany and N.I. Ahmed, 2006. Salicylic acid-induced adaptive response to copper stress in sunflower (Helianthus annuus L.). Int. J. Bot., 2: 372-379.
Direct Link - Hojjati, Y., A. Khalighi and A.R. Farokhzad, 2007. Chemical treatments of Eustoma cut flower cultivars for enhanced vase life. J. Agric. Soc. Sci., 3: 75-78.
Direct Link - Hussein, M.M., L.K. Balbaa and M.S. Gaballah, 2007. Salicylic acid and salinity effects on growth of maize plants. Res. J. Agric. Biol. Sci., 3: 321-328.
Direct Link - Jaleel, C.A., R. Gopi, G.M.A. Lakshmanan and R. Panneerselvam, 2006. Triadimefon induced changes in the antioxidant metabolism and ajmalicine production in Catharanthus roseus (L.) G. Don. Plant Sci., 171: 271-276.
CrossRefDirect Link - Karlidag, H., E. Yildirim and T. Metin, 2009. Salicylic acid ameliorates the adverse effect of salt stress on strawberry. Sci. Agric., 66: 180-187.
Direct Link - Khan, W., B. Prithiviraj and D.L. Smith, 2003. Photosynthetic responses of corn and soybean to foliar application of salicylates. J. Plant Physiol., 160: 485-492.
CrossRefDirect Link - Li, N., B.L. Parsons, D. Liu and A.K. Mattoo, 1992. Accumulation of wound-inducible ACC synthase transcript in tomato fruit is inhibited by salicylic acid and polyamines. Plant Mol. Biol., 18: 477-487.
PubMedDirect Link - Mahdavian, K., K.M. Kalantari and M. Ghorbanli, 2007. The effect of different concentrations of salicylic acid on protective enzyme activities of pepper (Capsicum annuum L.) plants. Pak. J. Biol. Sci., 10: 3162-3165.
CrossRefPubMedDirect Link - Mansour, M.M.F., 1998. Protection of plasma membrane of anion epidermal cells by glycinbetaine and praline against NaCl stress. Plant Phsiol. Biochem., 36: 767-772.
CrossRefDirect Link - Mansour, M.M.F., 2000. Nitrogen containing compounds and adaptation of plants to salinity stress. Biol. Plant, 43: 491-500.
CrossRefDirect Link - Mba, F.O., X. Zhi-Ting and Q. Hai-Jie, 2007. Salicylic acid alleviates the cadmium toxicity in Chinese cabbages (Brassica chinensis). Pak. J. Biol. Sci., 10: 3065-3071.
CrossRefPubMedDirect Link - Mittler, R., 2002. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci., 7: 405-410.
CrossRefPubMedDirect Link - Mohammad, M., R. Shibli, M. Ajouni and L. Nimri, 1998. Tomato root and shoot responses to salt stress under different levels of phosphorus nutrition. J. Plant Nutr., 21: 1667-1680.
Direct Link - Pollard, A. and R.G.W. Jones, 1979. Enzyme activities in concentrated solutions of glycinebetaine and other solutes. Plants, 144: 291-298.
CrossRef - Reezi, S., M. Babalar and S. Kalantari, 2009. Silicon alleviates salt stress, decreases malondialdehyde content and affects petal color of salt stressed cut rose (Rosa xhybrida L.) Hot Lady. Afr. J. Biotechnol., 8: 1502-1508.
Direct Link - Sankat, C.K. and S. Mujaffar, 1994. Water balance in cut anthurium flowers in storage and its effect on quality. Acta Hort., 368: 723-732.
Direct Link - Shi, Q. and Z. Zhu, 2008. Effects of exogenous salicylic acid on manganese toxicity, element contents and antioxidative system in cucumber. Environ. Exp. Bot., 63: 317-326.
CrossRefDirect Link - Witte, Y. and W.G. van Doom, 1991. The mode of action of bacteria in the vascular occlusion of cut rose flowers. Acta Hort., 298: 165-167.
Direct Link - Yildirim, B., F. Yasar, T. Ozpay, D. TurkOzu, O. Terzio lu and A. Tamkoc, 2008. Variations in response to salt stress among field pea genotypes (Pisum sativum sp. Arvense L.). J. Anim. Vet. Adv., 7: 907-910.
Direct Link - Zhou, X.M., A.F. Mackenzie, C.A. Madramootoo and D.L. Smith, 1999. Effects of stem-injected plant growth regulator, with or without sucrose, on grain production, biomass and photosynthetic activity of field-grown corn plants. J. Agron. Crop Sci., 183: 103-110.
CrossRef - Heath, R.L. and L. Packer, 1968. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys., 125: 189-198.
CrossRefPubMedDirect Link