
Desai Nivas
Biological Oceanography Division, National Institute of Oceanography, India
D.K. Gaikwad
Department of Botany, Shivaji University, Kolhapur, India
P.D. Chavan
Department of Botany, Shivaji University, Kolhapur, India
American Journal of Plant Physiology
Year: 2011 | Volume: 6 | Issue: 3 | Page No.: 157-166







ABSTRACT
A pot experiment was conducted to investigate the effects of different salt concentrations on growth features, photosynthetic pigments and nutrient status in two species of Morinda: Morinda citrfolia and Morinda pubescens seedlings. Increased salinity had significantly altered the qualitative growth parameters measured in terms of height and average leaf area. It was also observed that the chl. a, chl. b and total chlorophylls decreased with increasing salinity treatments in leaves of M. pubescens while in leaves of M. citrifolia chl. a and total chlorophylls remains stable under saline conditions whereas chl. b exhibited increasing trend with increasing salt doses. The level of sodium content in leaves of M. citrifolia increased with increasing salinity treatment, the potassium content also increased with increasing salinity treatments in leaves of M. pubescens and M. citrifolia. The Ca content is significantly reduced at 200 and 300 mM NaCl concentrations in M. pubescens leaves. The growth, photosynthetic pigments and nutrient responses of Morinda to different salt stress conditions indicated that this species could resilient to salt availabilities at lower concentrations and adapt to salinity conditions very well.
PDF Abstract XML References Citation
Received: February 21, 2011;
Accepted: May 18, 2011;
Published: July 09, 2011
How to cite this article
Desai Nivas, D.K. Gaikwad and P.D. Chavan, 2011. Physiological Responses of Two Morinda Species under Saline Conditions. American Journal of Plant Physiology, 6: 157-166.
DOI: 10.3923/ajpp.2011.157.166
URL: https://scialert.net/abstract/?doi=ajpp.2011.157.166
DOI: 10.3923/ajpp.2011.157.166
URL: https://scialert.net/abstract/?doi=ajpp.2011.157.166
INTRODUCTION
Abiotic environmental stresses such as low temperature, high salinity, drought, heat and flooding usually results in water deficits and affect plant growth and productivity (Holmberg and Bulow, 1998). Stresses induce an increased resistance to water uptake by roots (Lee et al., 2004). According to Roychoudhury et al. (2008), saline soils are one of the major biotic stresses that adversely affect the overall metabolic activities and cause plant demise. Salinity is one of the major environmental factors that leads to a deterioration of agricultural land and reduction in crop productivity worldwide (Chinnusamy et al., 2005). Under salt stress the osmotic potential in the soil solution exceeds the osmotic potential of plant cells due to the presence of higher concentration of salt which reduces the ability of plants to take up water and other essential nutrients (Munns et al., 2006). In halophytic species, the growth is not affected or even stimulated by salinity but in glycophytic species opposite response is observed. An ability of plant to acclimate to salt stress includes alterations at the leaf level, associated with morphological, physiological and biochemical characteristics whereby many plants adjust to high salinity and the consequent low soil water availability (Munns, 2002; Ashraf, 2004). The plants are exposed to both osmotic stress and specific ion toxicities during the salinity stress. Entry of sodium and chloride ions in large amount in chloroplasts leads to several disorders. Salt stress (NaCl) has both osmotic (cell hydration) and toxic (ion accumulation) effect on plants (Joseph and Jini, 2011). According to Munns and Tester (2008), induced nutritional disorders is one of the noticeable effects of salt stress in plants. The normal absorption of mineral nutrition is affected by salinity. The distribution of essential nutrients in plants mainly influnced by NaCl. The nutrient essenensiality varies from plant to plant and regulated by internal ionic status of root. Sodium is dominant cation in saline soil which is essential micronutrient. In different plant parts it is passively accumulated. Reducing the spread of salinization by better irrigation and farm management practices and increasing the salt tolerance of crops are hence important global issues (Munns, 2002).
The study was aimed to evaluate salt tolerance mechanism of two Morinda species under different salinity doses and the effect of salt stress on physical and physiological aspects of the species.
MATERIALS AND METHODS
Effect of NaCl salinity on Morinda seeds are obtained from the fruits of M. citrifolia growing in coastal areas of Goa and M. pubescens plants growing in the drought prone region of Osmanabad in October (2008). The seeds were isolated from fruits and healthy seeds were sorted out. Since the seeds have hard seed coat which restricted seed germination, seeds were mechanically scarified with nail cutter. The seeds were treated with 0.1% Mercuric chloride for 5 min for surface sterilization. The seeds were rinsed with distilled water for 4 to 5 times. Then the seeds were sown in earthenware pots (35 cm diameter with hole at the bottom) in garden soil containing farmyard manure in proportion of 3: 1. The pots were equally watered, twice a week and every care was taken to raise healthy and vigorously growing plants in each pot. The sturdy well established two-month-old seedlings were then transferred in separate pot. Two-month-old pots grown plants were subjected to salinity treatment (100, 200 and 300 mM NaCl) twice a week alternating with watering to 2 month old plants for a period of 40 days. Thus, the plants subjected to various treatments were of the same age at the time of harvesting for biochemical analysis. a. Seedling Growth analysis Growth analysis were carried out after completion of treatments, for this, five plants from each treatment were randomly selected, carefully uprooted, washed thoroughly with water to remove any dirt and dust particles on the surface of the plant parts and blotted to surface dry. Following growth parameters analysed for their growth and development using various parameters such as:
| • | Height of plant |
| • | Number of leaves per plant |
| • | Leaf area |
Photosynthetic pigments
Chlorophylls: Chlorophylls were estimated following the method of Arnon (1949) and chlorophylls were expressed in terms of mg/100 g fresh weight.
Inorganic constituents
Preparation of extract: Plant material (leaves) from each treatment and control was carefully washed in water and blotted to dryness leaves and roots were separated and subjected to drying at 60°C for 10 days till dried plant had constant weight. The oven dried plant material was powdered and Acid digestion method of Toth et al. (1948) has been followed for the analysis of inorganic constituents.
Estimation of mineral elements: Sodium and potassium were estimated using Flame Photometer (Systronics-128) by preparing standard concentrations. Calcium was estimated using Atomic Absorption Spectrophotometer. In case needed, appropriate dilution of plant extract was made with distilled water.
Statistical analysis: The experimental design was the randomized complete block design. The data are represented with respected standard deviation of means. Analysis of Variance (ANOVA) technique was employed for carrying out statistical analysis. The means values were compared with Least Significant Difference (LSD) Test at 5% level, following Snedecor and Cochran (1980).
RESULTS AND DISCUSSION
The effect salt stress on height of plant, number of leaves and leaf area of M. citrifolia and M. pubescens is depicted in Table 1 and 2. It is observed from the table that the qualitative growth parameters measured in terms of height and average leaf area are significantly altered by salt stress. It is evident that lower levels of salinity (100 and 200 mM) treatment showed increase in height of the M. citrifolia and M. pubescens. This increase in height at lower concentration (100 mM) is significant in M. pubescens while in M. citrifolia elevation in height is noticeable upto 200 mM salinity. The number of leaves increases significantly at lower levels of salinity (100 mM) in M. pubescens whereas M. citrifolia shows slight but steady increase in number of leaves at 100 mM and 200 mM salinity doses. It is also noticed that the number of leaves is twofold higher in M. pubescens than the M. citrifolia in control and 100 mM treatments, whereas number of leaves of M. pubescens considerably decreases at 200 mM and 300 mM salinity doses as compared to M. citrifolia.
| Table 1: | Comparison of means of salinity treatments for salt tolerance in Morinda citrifolia L. |
 | |
| Small letter indicates statistical difference among different treatments (values Mean±SD). LSD: Least significant difference; T0, Control group; T1-T3, Treatment groups. (T1-100 mM, T2-200 mM and T3-300 mM) | |
| Table 2: | Comparison of means of salinity treatments for salt tolerance in Morinda pubescens J.E. Smith |
 | |
| Small letter indicates statistical difference among different treatments (values Mean±SD). LSD: Least significant difference; T0, Control group; T1-T3, Treatment groups. (T1-100 mM, T2-200 mM and T3-300 mM) | |
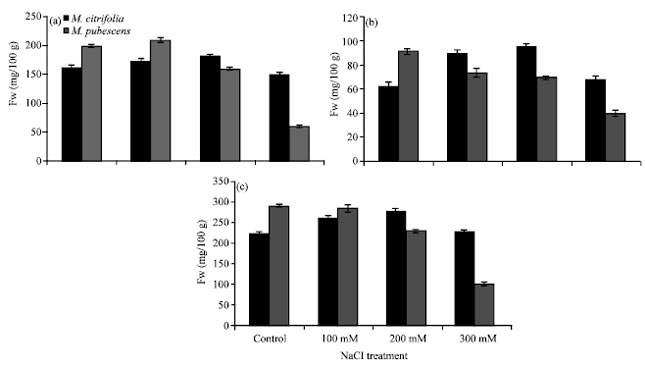 | |
| Fig. 1(a-c): | Effect of NaCl salinity on photosynthetic pigments in Morinda leaves (a) Chlorophyll a, (b) Chlorophyll b and (c) Total chlorophyll |
The average leaf area of M. citrifolia increases slightly at 100 and 200 mM salinity treatments, whereas average leaf area of M. pubescens increases slightly at 100 mM NaCl treatment which further decreases at higher levels (200 and 300 mM).
Effect of salt stress on the photosynthetic pigments in M. citrifolia and M. pubescens is shown in Fig. 1(a-c). It is clear from the Fig. 1(a-c) that chl. a, chl. b and total chlorophylls decreases with increasing salinity treatments in leaves of M. pubescens while, in leaves of M. citrifolia chl. a and total chlorophylls slightly increases under saline conditions whereas, chl. b exhibits quite noticeable increasing trend with increasing salt doses.
Influence of NaCl salinity on the contents of different mineral elements in leaves of M. citrifolia and M. pubescens are depicted in Table 1 and 2. The level of sodium content in leaves of M. citrifolia increases with increasing salinity treatment while it is slightly elevated in leaves of M. pubescens. Further it is also evident that the leaves of M. citrifolia accumulate more sodium than M. pubescens. The potassium content increases with increasing salinity treatments in leaves of M. pubescens and M. citrifolia and this increase is more significant at 100 mM salinity treatment. The Ca content is elevated at 100 and 200 mM salinity doses in M. citrifolia leaves while it is slightly increased in M. pubescens leaves at 100 mM NaCl treatment. The Ca content is significantly reduced at 200 and 300 mM NaCl concentrations in M. pubescens leaves.
The least significance differences at 5% level is depicted in Table 1. The growth attributes in M. citrifolia shows greater significant difference between the treated (T1 and T2) and control, the mineral content (Na+ and K+) also has quite significant differences among the treatments but the Cl- content has not that much significant difference.
In halophytic species, the growth is not affected or even stimulated by salinity but in glycophytic species opposite response is observed. It is now well established that adverse effect of salinity on plant growth in both due to osmotic stress as well as toxicity of ions like sodium and chloride. As salt concentration increases above a threshold level both growth rate and ultimate size of most plant species decreased progressively (Mass and Hoffmann, 1977). Rajpar et al. (2006) reported that increase in soil salinity decreases the height of wheat. Ghoulam et al. (2002) reported that high NaCl concentrations caused a great reduction in growth parameters of sugar beet cultivars such as leaf area, fresh and dry weight of leaves and roots but the leaf number was less affected. Amirjani (2010) reported reduction in soybean plant height with increasing salinity levels. De Pascale et al. (2003) reported decline in leaf area of pepper (Capsicum annum) subjected to salt stress. Various factors contribute to salinity induced reduction in plant growth is due to osmotic and specific ion effects. Salinity creates physiological drought and affects water relations in plants which in turn can lead to decline in water status and fresh weight of salt stressed plants. At the same time some halophytes belonging to chenopodiaceae develop succulent nature to bring about salt dilution (Waisel, 1972). The reduction in leaf growth may be due to ion toxicity, ion deficiency, ion imbalance, changes in growth regulators and/or other processes such as hardening of cell walls that limit cell expansion (Neumann et al., 1994; Nabil and Coudret, 1995).
Rozema and van Diggelen (1991) compared the physiological responses of 4 halophytes (Aster tripolium, Spartina anglica, Puccinellia maritima and Elymus pycnanthus) and noticed that the decrease of net photosynthesis per unit of leaf surface area. The same result was observed by Abdelly et al. (1995) in Medicago species. Fifty percent reduction in shoot DW, leaf surface area and leaf number at 150 mM NaCl, of Crithmum maritimum have been noticed by Hamed et al. (2004). In the present study we noticed more than 50% decreases in height of Morinda pubecsens with increase in salinity treatments (200 and 300 mM). This decrease reflects its salt sensitive nature. We observed some sign of toxicity, such as scorching of leaves at the highest levels of salt doses in M. pubescens. In case of M. citrifolia and M. pubescens a significant reduction in leaf area is evident at higher levels (300 mM) of NaCl salinity. It is evident that the number of leaves in M. pubescens is higher than the M. citrifolia while the leaf area of M. pubescens is lower than the M. citrifolia. Though the number of leaves increases in M. pubescens the leaves becomes lanceolate and linear than the M. citrifolia which may results in the reduction of leaf area in M. pubescens leaves than the M. citrifolia.
The plants are exposed to both osmotic stress and specific ion toxicities during the salinity stress. Entry of sodium and chloride ions in large amount in chloroplasts leads to several disorders. According to Strogonov et al. (1970), salinity might be affecting the strength of the forces binding the pigment-protein-lipid complex of chlorophyll in the thylakoid membrane. The decrease in the chlorophyll content under saline conditions is further attributed to increased chlorophyllase activity (Reddy and Vora, 1986). A decline in total chlorophylls under saline conditions has been noticed in several salt sensitive species such as greengram, asparagus, chickpea, sorghum and salvadora. (Azooz et al., 2004; Dagar et al., 2004). Parvaiz and Riffat (2005) also observed a decrease in chlorophyll content in Pisum sativum with increase in salt concentration from 50 to 200 mM. Djanaguiraman et al. (2006) noticed a decrease in chlorophyll content in rice under saline conditions. A significant decrease in the content of pigment fractions and consequently of the total chlorophyll content as compared with control plants with increasing salinity in Calotropis procera have been reported by Al-Sobhi et al. (2006). Jaleel et al. (2008) noticed a decrease in photosynthetic pigments of Catharanthus roseus under salt stress which caused a decrease of 11 and 38% in chlorophyll a, respectively at 50 and 100 mM NaCl treatment. Tantawy et al. (2009) also observed the decrease in total chlorophyll content in tomato with the increasing level of salinity. Khan et al. (2009) also reported a decrease in total chlorophyll content in six different genotypes of wheat where decrease was significant in sensitive genotypes in comparison to tolerant genotypes. In the present investigation a decline in total chlorophyll content beyond 100 mM NaCl salinity in M. pubescens indicates its salt sensitive nature relative to M. citrifolia. Strogonov (1973) proposed a significant correlation between salinity tolerance and maintenance of chlorophyll level. In the year 1994, Salama et al. (1994) observed that chlorophylls in the salt tolerant wheat cultivars the more effectively protected against the deleterious effects of Na because such plants show higher accumulation of vacuolar Na+ and osmolytes like to putrescine and quaternary ammonium compounds in plants. M. citrifolia leaves shows increase in chlorophyll content with increasing salinity treatments reflecting its salt tolerant halophytic nature and this increase in chlorophyll content might be due to the development of mechanism for osmotic adjustment as indicated by Strogonov et al. (1970). As the M. pubescens is found to grow in dry xeric conditions while M. citrifolia is common along the seashore showing halophytic habitat thus, in our experimental studies these two species also reflects differential response to salt stress, showing salt sensitive nature (M. pubescens) and salt tolerant nature (M. citrifolia).
The distribution of essential nutrients in plants mainly influnced by NaCl. The nutrient essenensiality varies from plant to plant and regulated by internal ionic status of root. Sodium is dominant cation in saline soil which is essential micronutrient. In different plant parts it is passively accumulated. Sodium has been reported to act as a substitute for potassium in some metabolic processes (Marschner et al., 1981). The role of sodium in maintaining favorable water balance has been suggested by Rains (1972). Gaikwad (1995) reported accumulation of sodium in leaf tissue of Amaranthus caudatus while in Amarnthus paniculatus Na accumulates in the root zone in response to salt stress. Sharma (1995) noticed accumulation of Na in all parts of wheat plant under saline condition. According to Loupassaki et al. (2002), in all cultivars of olive plant the higher Na% dm was recorded in the roots followed by shoot, mature and young leaves. According to Schachtman and Munns (1992), an ability to limit Na accumulation in the leaves might be an important mechanism in salt tolerance of Triticum species.
Pak et al. (2009) studied effect of salt stress on Na+ and K+ content in rape plants and noticed that Na+ ions significantly increases under salinity stress. Guan et al. (2010) noticed that as salt intensity increases the Na+ content also increases in Suaeda seedlings. Similar trend of results were also obtained during present study in Morinda. Sodium is the most essential microelement for halophytes (Marschner, 1986) and it also plays an important role in osmotic adjustment in halophytes. In light of above results, since M. citrifolia is halophytic species besides playing a positive role in metabolism, sodium might be also contributing to osmotic adjustment under saline conditions. Potassium is considered as a most essential macronutrient and its uptake is highly selective and closely coupled with metabolic activity, in cells, tissue as well as long distance transport via xylem and phloem. The tips and margin of the leaves scorch stem become weak and lodge under K deficiency. The critical concentration of K is in the range of 0.5 to 2% in dry matter and K requirement for optimum growth of plant is 2-5% (Marschner, 1986). High K concentration in cytoplasm and chloroplast is necessary to neutralize the soluble and insoluble macromolecule anion and to stabilize the pH 7-8 that is optimum for enzyme reaction (Patil, 2008). Accumulation of K+ in the leaves was studied by number of workers in monocotyledon halophyte (Spratina anglica) by Albert and Popp (1977) and Gorham et al. (1980). A preferential potassium uptake mechanism has been considered as one of the important mechanism of salt tolerance in marine algae and mangroves (Joshi, 1976). In the present investigation it is noticed that M. citrifolia plants are able to maintain potassium level above an optimal level (1%) in leaves over the entire range of the salt treatment. The leaf potassium appears to contribute to salinity tolerance, since, in this plant there is elevation in potassium content in response to salinity. Calcium is an essential element contributing in cell wall stabilization. Calcium plays important role in cell division and extension, Ca++ regulates the spindle activity. Clarkson and Hanson (1980) also reported that Ca is important in pectin synthesis. Fundamental role of Ca++ is to maintain membrane stability and cell integrity. According to Caldwell and Haug (1981), Calcium stabilizes cell membrane by binding phosphate and carbonate group of phospholipids.
According to Rains (1972), Ca protects injurious effects of H+ and other toxic ions. A decline in calcium content under saline conditions is reported to create a condition of calcium deficiency in salt sensitive plants (Bernstein, 1975) on the other hand in many plants salinity is found to promote calcium uptake. There are several reports of increasing Ca++ content in some plant species under saline conditions (Karadage and Chavan, 1983). Ramoliya et al. (2006) observed that calcium content increased in leaves of Prosopis cineraria subjected to salt stress. The accumulation of Ca2+ is considered for the protective effects of calcium on salt stress and sodium toxicity (Bressan et al., 1998). In case of M. citrifolia also calcium might be playing a similar role since calcium nutrition is not at all affected by salt stress and the calcium level in leaves is elevated under lower salt doses. On the other hand decrease in Ca content in M. pubescence at higher levels of salinity (200 and 300 mM) is particularly associated with the reduction of growth which indicates the salt sensitive nature of this species.
CONCLUSION
The two species of Morinda reflects differential morphological and physiological response to salt stress The morphological and physiological attributes of M. citrifolia under salinity doses clearly indicates its salt resistance nature whereas, M. pubescens shows its salt sensitive nature.
ACKNOWLEDGMENT
Authors are highly acknowledged to UGC-DRS-I-SAP and Head of Botany Department, Shivaji University, Kolhapur.
REFERENCES
- Abdelly, C., C. Lachaal, C. Grignon, A. Soltani and M. Hajji, 1995. Episodic association of strict halophytes and glycophytes in a saline, hydromorphic ecosystem in semi-arid zones. Agronomie, 15: 557-568.
Direct Link - Albert, R. and M. Popp, 1977. Chemical composition of halophytes from the Neusiedter Lake region in Austria. Oecologia, 27: 157-170.
CrossRef - Al-Sobhi, O.A., H.S. Al-Zahrani and S.B. Al-Ahmadi, 2006. Effect of salinity on chlorophyll and carbohydrate contents of Calotropis procera seedlings. Sci. J. King Faisal Univ., 7: 105-115.
Direct Link - Amirjani, M.R., 2010. Effect of salinity stress on growth, mineral composition, proline content, antioxidant enzymes of soybean. Am. J. Plant Physiol., 5: 350-360.
CrossRefDirect Link - Arnon, D.I., 1949. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol., 24: 1-15.
CrossRefPubMedDirect Link - Ashraf, M., 2004. Some important physiological selection criteria for salt tolerance in plants. Flora-Morphol. Distrib. Funct. Ecol. Plants, 199: 361-376.
CrossRefDirect Link - Azooz, M.M., M.A. Shaddad and A.A. Abdel-Latef, 2004. The accumulation and compartmentation of proline in relation to salt tolerance of three sorghum cultivars. Ind. J. Plant Physiol., 9: 1-8.
Direct Link - Bressan, R.A., P.M. Hasegawa and J.M. Pardo, 1998. Plants use calcium to resolve salt stress. Trends Plant Sci., 3: 411-412.
Direct Link - Caldwell, C.R. and A. Haug, 1981. Temperature dependence of the barley root plasma membrane-bound Ca2+ and Mg2+ dependant ATPase. Physiol. Plant, 53: 117-124.
CrossRef - Chinnusamy, V., A. Jagendorf and J.K. Zhu, 2005. Understanding and improving salt tolerance in plants. Crop Sci., 45: 437-448.
Direct Link - Clarkson, D.T. and J.B. Hanson, 1980. The mineral nutrition of higher plants. Ann. Rev. Plant Physiol., 31: 239-298.
CrossRefDirect Link - De Pascale, S., C. Ruggiero, G. Barbieri and A. Maggio, 2003. Physiological responses of pepper to salinity and drought. J. Am. Soc. Horticult. Sci., 128: 48-54.
Direct Link - Dagar, J.C., H. Bhagwan and Y. Kumar, 2004. Effect on growth performance and biochemical contents of Salvadora persica when irrigated with water of different salinity. Indian J. Plant Physiol., 9: 234-238.
Direct Link - Djanaguiraman, M., J.A. Sheeba, A.K. Shanker, D.D. Devi and U. Bangarusamy, 2006. Rice can acclimate to lethal level of salinity by pretreatment with sublethal level of salinity through osmotic adjustment. Plant Soil, 284: 363-373.
CrossRefDirect Link - Ghoulam, C., A. Foursy and K. Fares, 2002. Effects of salt stress on growth, inorganic ions and proline accumulation in relation to osmotic adjustment in five sugar beet cultivars. Environ. Exp. Bot., 47: 39-50.
CrossRefDirect Link - Gorham, J., L.L. Hughes and R.G.W.Y.N. Jones, 1980. Chemical composition of salt marsh plants from Ynys Mon (Anglesey): The concept of physiotypes. Plant Cell Environ., 3: 309-318.
CrossRef - Guan, B., J. Yu, Z. Lu, W. Japhet, X. Chen and W. Xie, 2010. Salt tolerance in two Suaeda species: Seed germination and physiological responses. Asian J. Plant Sci., 9: 194-199.
CrossRefDirect Link - Hamed, K.B., A. Debez, F. Chibani and C. Abdelly, 2004. Salt response of Crithmum maritimum, an oleagineous halophyte. Trop. Ecol., 45: 151-159.
Direct Link - Jaleel, C.A., B. Sankar, R. Sridharan and R. Panneerselvam, 2008. Soil salinity alters growth, chlorophyll content and secondary metabolite accumulation in Catharanthus roseus. Turk. J. Biol., 32: 79-83.
Direct Link - Karadage, B.A. and P.D. Chavan, 1983. Physiological studies in salinity tolerance of Sesbania aculeata POIR. Biol. Plant., 25: 412-418.
CrossRef - Khan, M.A., M.U. Shirazi, M.A. Khan, S.M. Mujtaba and E. Islam et al., 2009. Role of proline, K/NA ratio and chlorophyll content in salt tolerance of wheat (Triticum aestivum L.). Pak. J. Bot., 41: 633-638.
Direct Link - Lee, S.H., A.P. Singh and G.C. Chung, 2004. Rapid accumulation of hydrogen peroxide in cucumber root due to exposure to low temperature appears to mediated decreases in water transport. J. Exp. Bot., 55: 1733-1741.
CrossRef - Loupassaki, M.H., K.S. Chartzoulakis, N.B. Digalaki and I.I. Androulakis, 2002. Effects of salt stress on concentration of nitrogen, phosphorus, potassium, calcium, magnesium, and sodium in leaves, shoots, and roots of six olive cultivars. J. Plant Nutr., 25: 2457-2482.
CrossRefDirect Link - Marschner, H., P.J.C. Kuiper and A. Kylin, 1981. Genotypic differences in the response of sugar beet plants to replacement of potassium by sodium. Physiol. Plant., 51: 239-244.
CrossRef - Maas, E.V. and G.J. Hoffman, 1977. Crop salt tolerance-current assessment. J. Irrig. Drainage Div., 103: 115-134.
Direct Link - Munns, R., 2002. Comparative physiology of salt and water stress. Plant Cell Environ., 25: 239-250.
CrossRefDirect Link - Munns, R. and M. Tester, 2008. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol., 59: 651-681.
CrossRefDirect Link - Munns, R., R.A. James and A. Lauchli, 2006. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot., 57: 1025-1043.
CrossRefPubMedDirect Link - Nabil, M. and A. Coudret, 1995. Effects of sodium chloride on growth, tissue elasticity and solute adjustment in two Acacia nilotica subspecies. Physiol. Plant., 93: 217-224.
CrossRef - Neumann, P.M., H. Azaizeh and D. Leon, 1994. Hardening of root cell walls: A growth inhibitory response to salinity stress. Plant Cell Environ., 16: 303-309.
CrossRefDirect Link - Pak, V.A., M. Nabipour and M. Meskarbashee, 2009. Effect of salt stress on chlorophyll content, fluorescence, Na + and K + ions content in rape plants (Brassica napus L.). Asian J. Agric. Res., 3: 28-37.
CrossRefDirect Link - Rajpar, I., Y.M. Khanif, F.M. Soomro and F.M. Soomro, 2006. Effect of NaCl salinity on the growth and yield of inqlab wheat (Triticum aestivum L.) variety. Am. J. Plant Physiol., 1: 34-40.
CrossRefDirect Link - Rains, D.W., 1972. Salt transport by plant in relation to salinity. Annu. Rev. Plant Physiol., 23: 367-388.
CrossRef - Ramoliya, P.J., H.M. Patel, J.B. Joshi and A.N. Pandey, 2006. Effect of salinization of soil on growth and nutrient accumulation in seedlings of Prosopis cineraria. J. Plant Nutr., 29: 283-303.
CrossRefDirect Link - Reddy, M.P. and A.B. Vora, 1986. Changes in pigment composition, Hill reaction activity and saccharides metabolism in Bajra (Pennisetum typhoides S & H) leaves under NaCl salinity. Photosynthetica, 20: 50-55.
Direct Link - Rozema, J. and J. van Diggelen, 1991. A comparative study of growth and photosynthesis of four halophytes in response to salinity. Acta Oecol., 12: 673-681.
Direct Link - Roychoudhury, A., S. Basu, S.N. Sarkar and D.N. Sengupta, 2008. Comparative physiological and molecular responses of a common aromatic indica rice cultivar to high salinity with non-aromatic indica rice cultivars. Plant Cell Rep., 27: 1395-1410.
CrossRefPubMedDirect Link - Salama, S., S. Trivedi, M. Busheva, A.A. Arafa, G. Garab and L. Erdei, 1994. Effects of NaCl salinity on growth cation accumulation, chloroplast structure and function in wheat cultivars differing in salt tolerance. J. Plant Physiol., 144: 241-247.
Direct Link - Schachtman, D.P. and R. Munns, 1992. Sodium accumulation in leaves of Triticum species that differ in salt tolerance. Aust. J. Plant Physiol., 19: 331-340.
Direct Link - Tantawy, A.S., A.M.R. Abdel-Mawgoud, M.A. El-Nemr, Y.G. Chamoun, 2009. Alleviation of salinity effects on tomato plants by application of amino acids and growth regulators. Eur. J. Scientific Res., 30: 484-494.
Direct Link - Toth, S.J., A.L. Prince, A. Wallace and S.D. Mikkelsen, 1948. Rapid quantitative determination of eight mineral elements in plant tissue by a systematic procedure involving use of a flame photometer. Soil Sci., 66: 459-466.
Direct Link - Bernstein, L., 1975. Effects of salinity and sodicity on plant growth. Ann. Rev. Phytopathol., 13: 295-312.
CrossRefDirect Link - Holmberg, N. and L. Bulow, 1998. Improving stress tolerance in plants by gene transfer. Trends Plant Sci., 3: 61-66.
CrossRefDirect Link - Joseph, B. and D. Jini, 2011. Development of salt stress-tolerant plants by gene manipulation of antioxidant enzymes. Asian J. Agric. Res., 5: 17-27.
CrossRefDirect Link