
Heshmat Omidi
College of Agricultural Sciences, Shahed University, P.O. Box 18151/159, Tehran, Iran
LiveDNA: 98.1127
ORCID: 0000-0002-2728-1469
American Journal of Plant Physiology
Year: 2010 | Volume: 5 | Issue: 6 | Page No.: 338-349







ABSTRACT
The aim of present study was to get further information on the ability of drought stress applied to retard senescence and to frustrate inhibitory effects of drought-stress via comparing two canola genotypes differing in drought tolerance. The objective of this work was to evaluate the effects of the drought stress (control, -0.5, -1 and -1.5 Mpa) on two canola genotypes (Okapi and RGS). The experiment was factorial with four replications at experimental greenhouse at Center of Agriculture Research-Shahed, Iran, during 2006-2007. Morphological parameters of root and shoot dry weight, proline content osmolyte and enzyme activities of Ascorbate peroxidase (APx), Guaiacol peroxidase (GAx) and Catalase (Cat) in shoot and root of tissues canola genotypes were highly significant differences for main and interaction effects of two factors (Drought (D) and Genotype (G)) studied. Drought decreased the roots and shoots dry weight of canola plants and this effect was particularly significant at high level of stress. So, results indicated that proline content, ascorbate peroxidase (APx) and Guaiacol peroxidase (GAx) and Catalase (Cat) activity in root and shoot increased with enhancement of drought stress in the two genotypes. Of course, the proline content and antioxidant activities in Okapi genotype were more than RGS genotype. These results showed that Okapi genotype tolerated drought stress than RGS with increasing of proline content and antioxidant activities including GAx and APx. Shoot always maintained higher level of free Catalase (CAT) contents than the root. Therefore, the evaluation of both antioxidant enzymes and proline content can be suggested for Mechanisms of drought stress tolerance.
PDF Abstract XML References Citation
Received: May 17, 2010;
Accepted: August 12, 2010;
Published: October 13, 2010
How to cite this article
Heshmat Omidi, 2010. Changes of Proline Content and Activity of Antioxidative Enzymes in Two Canola Genotype under Drought Stress. American Journal of Plant Physiology, 5: 338-349.
DOI: 10.3923/ajpp.2010.338.349
URL: https://scialert.net/abstract/?doi=ajpp.2010.338.349
DOI: 10.3923/ajpp.2010.338.349
URL: https://scialert.net/abstract/?doi=ajpp.2010.338.349
INTRODUCTION
Plants are often exposed to environmental stresses through a life cycle. Several environmental factors adversely affect plant growth and development and final yield performance of a oilseed crop. Antioxidants enzyme and organic osmolytes such as proline are known to occur widely in higher plants and normally accumulates in large quantities in response to environmental stresses.
Among these stresses, in particular drought is the most serious problem that limits plant growth and crop productivity in agriculture. Drought stress is one of the major factors in limiting crop production, particularly in arid and semi-arid locations (Ashraf and Foolad, 2007). In such regions, germination and seedling establishment constitute are the most critical periods in the life-cycle of plants (Omidi et al., 2008). Consequently, adaptations during these stages can determine their natural distribution.
Canola is one of the most important oil seeds growing in many parts of Iran. It is very important to grow canola with high oil levels for agronomical benefits (Omidi et al., 2008). A large part of vegetable oil for human consumption in Iran is imported to the country and hence cultivation and appropriate management of oil seeds to increase yield is very important (Omidi et al., 2010). In recent years, due to the adaptation of canola to different climatic conditions, its production has been the center of attention and its cultivation area has increased from 19 000 ha in 2000 to 300 000 ha in 2008 (Omidi et al., 2010).
Drought stress lowers plant production worldwide. Drought stress results in a wide variety of physiological and biochemical changes in plants. Among them, the accumulation of low molecular weight solutes, such as proline commonly referred to as compatible solutes. Proline accumulation in plant cells exposed to water stress is a widespread phenomenon and is often considered to be involved in stress resistance mechanisms, although it’s precise role continues to be controversial (2-5). Water scarcity has been considered a limiting factor to crop production in arid and semiarid regions of the world (Hsu et al., 2003; Mudgal et al., 2010). Under environmental stress conditions, many plants accumulate several kinds of compatible solutes such as proline, glycinebetaine (from now on betaine), sugars and polyols (Ashraf and Foolad, 2007).
Oilseed crop studies published to date have concentrated on the various species in response to different biotic and abiotic factors and many functional proteins have been identified, e.g., many kinds of proteins up-regulated in response to stress in Suaeda aegyptiaca leaves (Askari et al., 2006), rice leaves (Kim et al., 2005a, b) and rice roots (Yan et al., 2005) and proteins involved in responses to osmotic stress in Arabidopsis (Ndimba et al., 2005). The responses of plants to stress conditions have evolved a variety of physiological and biochemical processes, e.g., solute accumulation and the development of enzymatic antioxidant systems (Ashraf and Foolad, 2007). The amino acid proline (Pro) is known to occur widely in higher plants and normally accumulates in large quantities in response to environmental stresses (Khedr et al., 2003). Plants employ antioxidant defense mechanisms against oxidative damage of reaction oxygen species. Proline and betaine enhance antioxidant defense systems in plant responses to various oxidative stresses (Khedr et al., 2003; Demiral and Turkan, 2004; Okuma et al., 2004; Park et al., 2006; Molinari et al., 2007).
Some of the proteins made in response to such stresses are enzymes that synthesize compounds believed to serve as osmoprotectants, including proline, polyamines, glycine, betaine and soluble sugars. Soluble sugars accumulate in the leaves of many plant species when they experience water stress (Udomchalothorn et al., 2009). Earlier reports have shown that both proline and betaine improve salt tolerance in tobacco BY-2 cells by increasing the activity of enzymes involved in the antioxidant defense and methylglyoxal detoxification systems (Hoque et al., 2007a, b). Recent research has focused the protective roles of proline and betaine in cell death against various oxidative stresses (Chen and Dickman, 2005). The expressions of antioxidant enzymes are up-regulated upon some abiotic stress, including, e.g., responses in shoot and root tissues, cells and organelles in the model plant Arabidopsis (Hsu et al., 2003; Dixon et al., 2005; Schurmann, 2003), responses to drought stress in maritime pine (Costa et al., 1998), sugar beet (Hajheidari et al., 2005), rice (Ali and Komatsu, 2006), Elymus elongatum (Gazanchian et al., 2007) and wheat (Hajheidari et al., 2007), responses to salt stress in rice (Ashraf and Foolad, 2007) and S. aegyptiaca (Kim et al., 2005a, b) and responses in poplar to heat stress (Ferreira et al., 2006) and ozone exposure (Ozden et al., 2009). The association between drought stress damage and senescence, on one hand and osmotic adjustment mechanisms and biochemical processes on the other, are little considered. Several recent studies focus on evaluation of water deficit on oilseed establishment adaptation. However, we hypothesized that solute accumulation and allocation under drought conditions depend on aging process of plant tissue.
In many researches, drought tolerance in plants have been studied in relation to regulatory mechanisms of osmotic (Ashraf and Foolad, 2007). drought stress, like other abiotic stresses, can also lead to oxidative stress through the increase in reactive oxygen species (ROS), such as superoxide (O2-), hydrogen peroxide (H2O2) and hydroxyl radicals (OH), which are highly reactive and may cause cellular damage through oxidation of lipids, proteins and nucleic acids (Apel and Hirt, 2004; Mudgal et al., 2010; Nojavan and Khorshidi, 2006).
To be able to endure oxidative damage under conditions which favors increased oxidative stress such as drought, plants must possess efficient antioxidant system. Plant cells have evolved a complex antioxidant system, which is composed of lowmolecular mass antioxidants (glutathione, ascorbate and carotenoids) as well as ROS-scavenging enzymes, such as superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX), guaiacol peroxidase (GPX) and glutathione reductase (GR) (Apel and Hirt, 2004; Bhardwaj et al., 2007).
Canola (Brassica napus L.) species is considered to be a relatively moderately drought-sensitive genus within which there is certain variability towards drought tolerance. Because the canola species used in this study were cultivated under various conditions throughout the world, comparisons between varietals are questionable; nonetheless, identification of varietal differences in response to drought between some canola species was the aim of the present study.
The recent study was conducted in order to assess two canola varieties for their drought tolerance and to give more information on the significance of osmotic adjustment and osmolytes accumulation under drought stress. In this case, the following questions were therefore addressed: (1) how much these processes are influenced by Root-Shoot tissue senescence and (2) what is the degree of tolerance of the two tested varieties? For this urpose we investigated the changes in Enzyme assays status and solutes accumulation especially proline, in canola roots and shoots and to relate them to defense strategies in terms of osmotic adjustment of each variety against drought stress.
MATERIALS AND METHODS
Experimental Design
The experiments were carried out at the Basic Science Experimental Station (BSES), 25 km, South of Tehran, Iran in 2006-2007 and. The experimental design was Factorial of drought stress and canola genotypes with four replications. The four drought stress, were including: No stress-(control), minimum stress (-0.5 Mpa), moderate (-1 Mpa) and High stress (-1.5 Mpa). The canola autumn genotypes were consist of RGS and Okapi.
Plant Material and Growth Conditions
The one hundred seeds of both genotypes were surface sterilized with 5% sodium hypochlorite for 5 min and then thoroughly rinsed with distilled water before further experimentation. Seeds of canola genotypes were grown in the containers with Hoagland nutrient solution. Surface-sterilized seeds were germinated in the dark on sand, moistened with distilled water. Ten day old seedlings with the uniform size were transferred to hydroponic culture in plastic container with 2 L of nutrient solution. Plants were grown under greenhouse conditions with a 14 h photoperiod of natural daylight, maximum and minimum temperatures were 26 and 18°C, respectively and relative humidity was 60-80% on average and the average of photosynthetically active radiation was 500 mol/m2/sec. Canola seedlings were grown under controlled conditions as previously described for 3 weeks. Four stress treatments were imposed by adding D0 = 0 (control), D1 = 20.33, D2 = 29.61 and D3 = 36.80 g/100cc PEG 6000 to the nutrient solution after 10 day old plants. The Osmotic Potential (OP) of the treatments was calculated using (Michel and Kaufmann, 1973). Twenty days after drought treatment, the plants were harvested. The Shoot and root samples collected from both varieties were frozen in liquid nitrogen and stored at-20°C in order to disrupt cell membranes. Shoot tissues were thawed and centrifuged at 1200xg for 25 min at 4°C to extract the cell sap. The analysis of biochemical solutes were performed in roots and Shoot stages during the experiment. Moreover, the proline concentrations were determined from the primary shoot and root apex.
Proline Content
Proline content was estimated using ninhydrin reaction method (Bates et al., 1973), except that phosphoric acid was omitted to avoid interference with concentrated sugars (Magne and Larher, 1992). Shoot discs were placed into tubes (five discs per tube) and 2 mL of distilled water were added. The tubes were kept for 30 min in a boiling water bath to extract hot water soluble compounds and then cooled to room temperature. One hundred microliter of the water extract was mixed with 400 mL of distilled water and 1 mL of ninhydrin reagent (0.5 g of ninhydrin in 30 mL of glacial acetic acid and 20 mL of distilled water). The mixture was boiled for 20 min in a water bath and then cooled on ice-water. The chromophore formed was extracted with 3 mL of toluene by vigorous shaking and tubes were placed in the dark for a minimum of 2 h. Absorbance of the resulting organic layer was measured at 520 nm. Calibrations were made with 1 mM L-proline as a standard.
Enzyme Assays
Fresh shoot samples (0.4 g) were ground in liquid nitrogen using a mortar and pestle and the ground samples were homogenized in an ice bath in 4 mL single extraction solution containing 50 mM Tris-HCl (pH 7.0), 1 mMEDTA, 20% glycerin, 1 mMascorbic acid (AsA), 1 mM dithiothreitol (DTT), 1 mM glutathione (GSH) and 5 mM MgCl2 and extracted at 48°C. The homogenate was centrifuged at 8,000 g at 48°C for 15 min. This method was modified as described by Knorzer et al. (1996). The supernatant was stored in volumes of 0.4 mL at 70°C for the analysis of soluble protein and the following determination of antioxidant enzymes. All experiments were performed at 258°C and completed within a day.
Catalase (EC 1.11.1.6, CAT)
A modification of the procedure of Aebi (1984) was used to analyze the CAT activity. The CAT activity was determined by directly measuring the decomposition of H2O2 at 240 nm. The reaction mixture contained 50 mM Tris-HCl buffer (pH 7.0) and 0.1 mM EDTA. The reaction mixture and 750 mM H2O2 were warmed up at 25°C. Then, 50 μL enzyme solution was mixed with 2.9 mL reaction mixture and 50 μL 750 mM H2O2 was added to start the reaction. The absorbance at 240 nm was read every 30 sec. The CAT activity was followed by a decrease of absorbance between 0.5 and 3 min. The CAT activity was calculated and expressed as nmol H2O2 mg-1 protein min-1.
Guaiacol Peroxidase (EC 1.11.1.7, GPx)
The Guaiacol peroxidase (GPx) activity was measured using a modification of the method of Chance and Maehly (1955). The assay mixture contained 50 mM Tris-HCl (pH 7.0), 0.1 mMEDTA, 10 mM guaiacol and 5 mM H2O2. Then, 50 μL enzyme solution was added to start the reaction in a total volume of 3.0 mL. Changes in the absorbance of the brown guaiacol at 470 nm between 0.5 and 3.5 min were recorded for calculating the GPx activity, which was expressed as micromol guaiacol mg-1 protein min-1.
Ascorbate Peroxidase (EC 1.11.1.11, APx)
The APx activity was measured using a modification of the procedure of Nakano and Asada (1981). The reaction mixture contained 50 mM Tris-HCl buffer (pH 7.0), 0.1 mM EDTA, 0.1 mM H2O2. Then, 2.93 mL of the reaction mixture was homogenized with 50 μL enzyme solution and the reaction was started by 20 μL 30 mM AsA (a total volume of 3 mL). The H2O2-dependent oxidation of ascorbate was followed by a decrease in the absorbance at 290 nm within 1 min. The APx activity was calculated and expressed as μmol AsA mg-1 protein min-1.
Statistical Analysis
The results of the morphological and biochemical parameters were statistically analyzed by one-way ANOVA and Least Significant Difference (LSD) test to determine significant differences among group means (SAS Institute, 1988). A p-value of 0.05 was considered statistically significant. The statistical package SAS, version 9.1 (SAS/STAT Software for PC. SAS Institute Inc., Cary, NC, USA) was used for all the applied statistical analyses (Steel and Torrie, 1980).
RESULTS
Effects of the four Drought stress (D) treatments on the growth of canola genotypes is shown in Table 1. The application of stress significantly affected plant growth components such as root and shoot dry weight (DW), proline content and antioxidant enzymes activity of canola genotypes (Table 1-2, Fig. 1-4). Drought decreased the roots and shoots dry weight of canola plants and this effect was particularly significant at high level of stress.
| Table 1: | Results of variance analysis (ANOVA) of Drought stress (D), genotype (G) and their interaction for shoot and root dry weight, proline concentration and antioxidant activity |
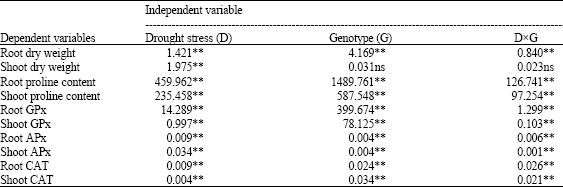 | |
| Numbers represent F-values at 5% level.. * and **significant at p<0.05 and p<0.01; ns: Non-significant. GPx: The guaiacol peroxidase; APx: Ascorbate peroxidase, CAT: Catalase | |
| Table 2: | The interaction effects of drought stress and varieties on canola antioxidant enzymes activity |
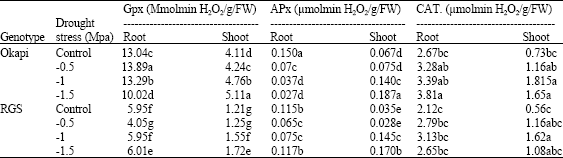 | |
| Means within each column followed by the same letter are not statistically different at 0.05 | |
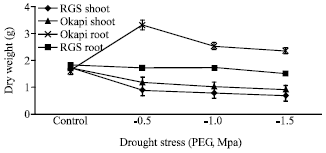 | |
| Fig. 1: | Change of Dry Weight (DW) of root and shoot canola genotypes under drought stress |
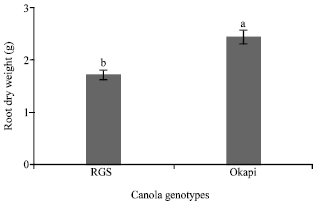 | |
| Fig. 2: | Root dry weight of canola genotypes |
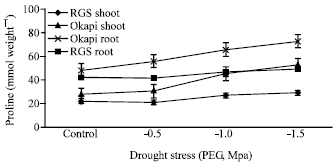 | |
| Fig. 3: | Effect of drought stress on changes in root and shoot proline content of canola genotypes |
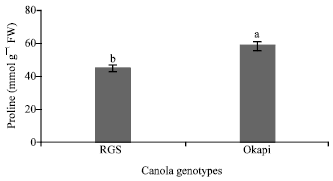 | |
| Fig. 4: | Proline content under canola genotypes |
The growth of shoot canola seedling was decreased by drought levels of -0.5 Mpa and above; this decrease was severe at -1.5 Mpa, whereas a level of -1 Mpa was lethal to the plants (Fig. 1). At all levels of drought, the shoot dry weight decrease more and this showed the sensitivity of shoot than root to drought stress.
The root dry matter in the Okapi genotype was higher than in the RGS genotype (Fig. 2). This study showed that canola genotypes differ in their capacity to utilize water content under drought stress. At higher water availability, Okapi also had the greatest root dry weight but there were no differences in shoot growth of the two canola genotypes. When water availability was low, the canola RGS genotype achieved the lowest shoot growth and Okapi the highest root growth, respectively. Okapi was characterized by lower water content concentrations in the roots than the RGS canola genotypes which suggest that this genotype can achieve high growth rates at low plant drought stress content (Fig. 1-2). The interaction of genotypes and drought stress on dependent variable of Dry Weight (DW) showed the highest significant reduction of RGS shoot among the genotypes tested in drought treatment compared to the unstressed plants.
Proline Content
There were highly significant differences for root and shoot proline content for two factors (drought and genotype) studied. In this study, proline content in root was substantially higher than in shoot genotypes. Proline content increased in root than shoot significantly when exposed to drought stress level (Fig. 3). The content of proline was higher in the Okapi genotype than in the RGS genotype (Fig. 4). With a drought concentration of -0.5 Mpa treatment in the nutrient solution, the proline concentration in root and shoot in canola genotypes increased to twofold values compared to the control treatment (Fig. 3) and drought stress had a significant effect on it (Table 1). Among the genotypes, Okapi had the maximum concentration of proline in root and shoot than RGS in all drought treatment (Table 1, Fig. 3-4).
Antioxidant Enzymes Activity
Water deficit increased CAT activity about 1.26 multiple more than the control plants and Apx activity increased in shoot of stressed canola plants about at least 1.98 multiple higher than the control plants, as shown in Table 2. Moderate stress (-1 Mpa) notably increased the specific GAx peroxidase and CAT activities in the canola plants. In contrast, these antioxidant enzymes were increased under high stress conditions, particularly changed in the plants treated with high level of stress (Table 2). By increasing drought levels from 0 to -1.5 Mpa, the activity of two antioxidant enzymes (APX and CAT) in shoot and root increased but GPX in two genotypes decreased. The enzymatic activity (APx and GAx) of shoot and root plants treated in combination with drought stress was increased by all drought levels, as shown in Table 2. The result showed that canola genotypes in case of antioxidant enzyme activities responses, in which Okapi genotype had more Guaiacol peroxidase activity than RGS genotype. The enzymatic activity (APx and CAT) of canola genotypes were different and Okapi genotype (as tolerant) in shoot and roots organs had the most enzymatic activity (Table 2).
Correlations Coefficients
Correlation coefficients among canola seedling traites and antioxidant enzymes activity shown in Table 3. In this study, results showed a negative relationship between root dry weight and the shoot dry weight (r2 = -0.44**), GPx in shoot (r2 = -0.576***) and GPx in canola root (r2 = -0.672***).
| Table 3: | Correlation coefficients among canola seedling traites and antioxidant enzymes activity |
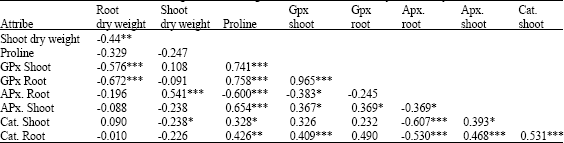 | |
| *, **and ***significant at the 5, 1 and 0.01 % levels of probability, respectively | |
Additionally, we found a strong negative correlation between proline accumulation and Apx root (r2 = -0.600***) under stress. The Proline content was significantly correlated with Gpx. shoot content (r2 = 0.741***), Gpx root activity (r2 = 0.758***) and APx. shoot (r2 = 0.654***) of the plants.
DISCUSSION
Drought is a major factor in reducing the growth and productivity of plants and involves different responses: dehydration of the cells as a result of the low water potential and nutritional imbalance caused by reducing of nutrition absorb elements by unavailable moisture in the uptake and translocation of essential nutrients (Ehsanpour and Fatahian, 2003; Zhu, 2002). The physiological and biochemical responses of B. Napus genotypes were investigated under drought stress. The shoot and root dry weight of each genotype were significantly affected by stress (Fig. 1-2). Shoot and root and dry weight of stressed plants was reduced compared to unstressed plants. There are several reports to show contradictory results on the effect of drought and salt stress on biomass accumulation (Hsu et al., 2003). Some studies have reported that biomass accumulation increased (Demiral and Turkan, 2004) while others have found that it decreased (Ali and Komatsu, 2006) or remained unchanged (Zhu, 2002) during stress condition.
The increase in drought stress, increased proline concentration in both root and shoot tissues of canola genotypes (Fig. 3-4). The accumulation of free proline is a common response to a wide range of biotic and abiotic stresses (Bohnert et al., 1995). A positive correlation between the proline content and salt-stress tolerance has been previously observed in a wide range of plant species (Bohnert et al., 1995). Many studies have suggested that proline is involved in intracellular osmotic adjustments between the cytoplasm and vacuole. It has also been proposed that proline can stabilize the cellular structure and scavenge free radicals or act as a storage compound for carbon and nitrogen to allow recovery of lipids from stress (Delauney and Verma, 1993). The synthesis, accumulation and degradation of proline in plants are highly regulated in response to environmental changes. When the synthesis of proline is induced by osmotic stress, the P5CS gene has a role as a rate-limiting enzyme in the salt-stress pathway in plants. A correlation between levels of transcription of the P5CS gene and proline content has been shown in a number of plants. This result suggests that up to a certain level of drought stress (-1 Mpa) treatment, canola plants can produce higher amount of proline in protecting cells against drought stress. However, under control and almost low stress condition (up to -0.5 Mpa of drought) proline content was not increased (Fig. 4), which might be the possible reason of not a success of intracellular osmotic adjustments, thus resulting unreduced in plant growth and decrease of growth plantlet. This has been previously observed by findings of Bohnert et al. (1995).
The mechanisms of Pro action are not fully understood, but it has been suggested that in addition to its role as an osmolyte for osmotic adjustment, Pro contributes to stabilizing sub-cellular structures (e.g., membranes and proteins), scavenging free radicals and buffering cellular redox potential under stress conditions (Ashraf and Foolad, 2007). The variation in drought tolerance between the Okapi and the genotypes’ of RGS could be due to differences in the level of soluble components e.i. proline content and thus drought tolerance may be partly attributed to an increased level of solutes, which protect and support cellular structures under stress (Nojavan and Khorshidi, 2006).
Water deficit could cause oxidative damage. Therefore, plant cells need different mechanisms, which enable the detoxification of excess ROS and keep the balance of the formation and removal of ROS. The increased activities of CAT, APx and GPx, detected in this study are presumed to limit cellular damage and enhance the plant’s antioxidative capacity to defend stress. Some proteins, a part of the plant antioxidant system, are rapidly activated and up regulated in response to oxidative stress generated by drought stress in canola seedling, including, ascorbate, glutathione (Bhardwaj et al., 2007). The most important reducing substrate for H2O2 detoxification in plant cells is ascorbate. Ascorbate peroxidase reduces H2O2 to water, with the concomitant generation of monodehydroascorbate (Aebi, 1984). Thioredoxin peroxidase is a part of the thioredoxin antioxidant system, acting after the oxidative stress is generated in the electron transport chain through salt exposure and inducing redox changes in different compartments (Delauney and Verma, 1993). It has been indicated that drought stress affects the physiology and biochemistry of plant cells under in vivo and in vitro conditions. In this context, increased proline and activities of CAT, APx, GPx has been reported in plants grown under stress (Hoque et al., 2007a). Drought stimulated the accumulation of the ROS including H2O2 in plants cells. The metabolism of H2O2 is dependent on various functionally interrelated antioxidant enzymes such as catalases and peroxidases. These enzymes are involved in climination of H2O2 from stressed cells (Kim et al., 2005a, b; Nojavan and Khorshidi, 2006).
Present results demonstrated that catalase (CAT) and ascorbate peroxidase (APX) activities were significantly increased (Table 2) and guiacual peroxidase (GPX) decreased in shoot and root of canola genotypes under drought stress. These results about guiacual peroxidase activity were supported by the findings of Hoque et al. (2008). The reductions of GPX activity suggest that these enzymes were unable to completely neutralize H2O2 resulted from the oxidative stress. Thus, results suggest that CAT and APX activities coordinated with SOD activity play a central protective role in the O2-and H2O2 scavenging process which has been reported similar by findings (Hoque et al. 2007b). When the activity values of the H2O2 scavenging enzymes were compared (Table 2) it was observed that CAT and APX had a much higher scavenging activity than GPX in shoot and root of stressed canola plants.
It has been demonstrated that drought can limit growth, fresh and dry weight of canola genotypes and lead to an increase in ROS generation, which are responsible for a secondary oxidative stress that can damage cellular structure and metabolism. The efficiency of the latter process is dependent upon the plant antioxidant defense mechanisms. the obtained results showed that the differences in the antioxidative enzyme activities of shoot and root may, at least in part, explain the greater tolerance of Okapi to drought stress when compared to other genotype. RGS however had the highest decreasing in fresh and dry weight but it showed the highest CAT activity at high drought level in root, the highest activity of GPx I in drought treatments and the highest proline content in shoot when subjected to drought stress. It can be deduced that antioxidant activity especially CAT and APX activities play a protective role to drought tolerance in canola genotypes. Among the genotypes, Okapi genotype used these two mechanisms to drought tolerance.
CONCLUSION
As conclusion, the different organs of canola plant treated (roots or leaves) are considered in drought, the accumulation and remobilization of the major solutes contributing to the osmotic adjustment were shown to be affected by shoot and root tissue senescence. The later growth stages, of both organs seem to be more protected but sometimes more reactive toward the application of drought stress. Results of this research suggest also that osmolytes accumulation such as proline and antioxidant enzymes activity as GPx, CAT and APX could be a part of the drought resistance mechanisms developed by canola tolerant genotype and could be exploited in breeding programs for improved drought stress tolerance.
ACKNOWLEDGMENTS
The authors would like to thank Dr. Tahmasebi and Mr. Kazemi for their assistance in the laboratory and field, respectively. In addition Dr. Karami and Dr. Habibi are thanked for their helpful correspondence.
REFERENCES
- Ashraf, M. and M.R. Foolad, 2007. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot., 59: 206-216.
CrossRefDirect Link - Omidi, H., Z. Tahmasebi, H. Torabi and M. Miransari, 2008. Soil enzymatic activities and available P and Zn as affected by tillage practices, canola (Brassica napus L.) cultivars and planting date. Eur. J. Soil Biol., 44: 443-450.
Direct Link - Kim, S., J. Lim, M. Park, Y. Kim, T. Park, Y. Seo and S. Yun, 2005. Enhanced antioxidant enzymes are associated with reduced hydrogen peroxide in barley roots under saline stress. J. Biochem. Mol. Biol., 38: 218-224.
Direct Link - Omidi, H., Z. Tahmasebi, H.A.N. Badi, H. Torabi and M. Miransari, 2010. Fatty acid composition of canola (Brassica napus L.), as affected by agronomical, genotypic and environmental parameters. Comptes Rendus Biologies, 333: 248-254.
CrossRef - Hsu, S.Y., Y.T. Hsu and C.H. Kao, 2003. The effect of polyethylene glycol on proline accumulation in rice leaves. Biol. Plant, 46: 73-78.
CrossRefDirect Link - Yan, S.P., Z.C. Tang, W. Su and W. Sun, 2005. Proteomic analysis of salt stress-responsive proteins in rice root. Proteomics, 5: 235-244.
Direct Link - Askari, H., J. Edqvist, M. Hajheidari, M. Kafi and G.H. Salekdeh, 2006. Effects of salinity levels on proteome of Suaeda aegyptiaca leaves. Proteomics, 6: 2542-2554.
Direct Link - Kim, D.W., R. Rakwal, G.K. Agrawal, Y.H. Jung and J. Shibato et al., 2005. A hydroponic rice seedling culture model system for investigating proteome of salt stress in rice leaf. Electrophoresis, 26: 4521-4539.
PubMed - Ndimba, B.K., S. Chivasa, W.J. Simon and A.R. Slabas, 2005. Identification of arabidopsis salt and osmotic stress responsive proteins using two-dimensional difference gel electrophoresis and mass spectrometry. Proteomics, 5: 4185-4196.
Direct Link - Khedr, A.H.A., M.A. Abbas, A.A. Abdel Wahid, W.P. Quick and G.M. Abogadallah, 2003. Proline induces the expression of salt-stress-responsive proteins and may improve the adaptation of Pancratium maritimum L. to salt-stress. J. Exp. Bot., 54: 2553-2562.
CrossRefDirect Link - Demiral, T. and I. Turkan, 2004. Does exogenous glycinebetaine affect antioxidative system of rice seedlings under NaCl treatment? J. Plant Physiol., 161: 1089-1100.
CrossRefDirect Link - Park, E.J., Z. Jeknic and T.H.H. Chen, 2006. Exogenous Application of glycinebetaine increases chilling tolerance in tomato plants. Plant Cell Physiol., 47: 706-714.
Direct Link - Molinari, H.B.C., C.J. Marur, E. Daros, M.K.F. Campos and J.F.R.P. Carvalho et al., 2007. Evaluation of the stress-inducible production of proline in transgenic sugarcane (Saccharum spp.): Osmtic adjustment, chlorophyll fluorescence and oxidative stress. Physiol. Plant., 130: 218-229.
CrossRefDirect Link - Udomchalothorn, T., S. Maneeprasobsuk, E. Bangyeekhun, P. Boon-Long and S. Chadchawan, 2009. The role of the bifunctional enzyme, fructose-6-phosphate-2-kinase/fructose-2, 6-bisphosphatase, in carbon partitioning during salt stress and salt tolerance in Rice (Oryza sativa L.). Plant Sci., 176: 334-341.
CrossRef - Hoque, M.A., M.N.A. Banu, E. Okuma, K. Amako, Y. Nakamura and Y. Shimoishi, 2007. Exogenous proline and glycinebe-taine increase NaCl-induced ascorbate-glutathione cycle enzyme activities and proline improves salt tolerance more than glycinebetaine in tobacco Bright Yellow-2suspension-culturedcells. J. Plant Physiol., 164: 1457-1468.
- Hoque, M.A., M.N.A. Banu, Y. Nakamura, Y. Shimoishi and Y. Murata, 2008. Proline and glycinebetaine enhance antioxidant defense and methylglyoxal detoxification systems and reduce NaCl-induced damage in cultured tobacco cells. J. Plant Physiol., 165: 813-824.
CrossRefPubMedDirect Link - Chen, C. and M.B. Dickman, 2005. Proline suppresse sap optosisin the fungal pathogen Colletotrichum trifolii. Proc. Natl. Acad. Sci. USA., 102: 3459-3464.
PubMed - Ali, G.M. and S. Komatsu, 2006. Proteomic analysis of rice leaf sheath during drought stress. J. Proteome Res., 5: 396-403.
CrossRef - Dixon, D.P., M. Skipsey, N.M. Grundy and R. Edwards, 2005. Stress-induced protein sglutathionylation in Arabidopsis. Plant Physiol., 138: 2233-2244.
Direct Link - Schurmann, P., 2003. Redox signaling in the chloroplast: The ferredoxin/thioredoxin system. Antioxid. Redox Signal, 5: 69-78.
PubMed - Costa, P., N. Bahrman, J.M. Frigerio, A. Kremer and C. Plomion, 1998. Water-deficit-responsive proteins in maritime pine. Plant Mol. Biol., 38: 587-596.
Direct Link - Hajheidari, M., M. Abdollahian-Noghabi, H. Askari, M. Heidari and S.Y. Sadeghian et al., 2005. Proteome analysis of sugar beet leaves under drought stress. Proteomics, 5: 950-960.
CrossRef - Gazanchian, A., M. Hajheidari, N.K. Sima and G.H. Salekdeh, 2007. Proteomeresponse of elymus elongatum to severe water stress and recovery. J. Exp. Bot., 58: 291-300.
PubMed - Hajheidari, M., A. Eivazi, B.B. Buchanan, J.H. Wong, I. Majidi and G.H. Salekdeh, 2007. Proteomics uncovers a role for redox in drought tolerance in wheat. J. Proteome Res., 6: 1451-1460.
PubMed - Ferreira, S., K. Hjerno, M. Larsen, G. Wingsle and P. Larsen et al., 2006. Proteome profiling of populus euphratica Oliv. upon heat stress. Ann. Bot., 98: 361-377.
PubMed - Ozden, M., U. Demirel and A. Kahraman, 2009. Effects of proline on antioxidant system in leaves of grapevine (Vitis vinifera L.) exposed to oxidative stress by H2O2. Sci. Hort., 119: 163-168.
CrossRef - Michel, B.E. and M.R. Kaufmann, 1973. The osmotic potential of polyethylene glycol 6000. Plant Physiol., 51: 914-916.
CrossRefPubMedDirect Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Magne, C. and F. Larher, 1992. High sugar content of extracts interferes with colorimetric determination of amino acids and free proline. Anal. Biochem., 200: 115-118.
CrossRefPubMedDirect Link - Knorzer, O.C., J. Durner and P. Boger, 1996. Alterations in the antioxidative systerm of suspension-cultured soybean cells (Glycine max) induced by oxidative stress. Physiol. Plant., 97: 388-396.
Direct Link - Aebi, H., 1984. Catalase in vitro. In: Methods in Enzymology, Packer, L., Academic Press, Cambridge, Massachusetts, United States, ISBN: 9780121820053, pp: 121-126.
CrossRefDirect Link - Chance, B. and A.C. Maehly, 1955. Assay of catalases and peroxidases. Methods Enzymol., 2: 764-775.
CrossRefDirect Link - Nakano, Y. and K. Asada, 1981. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in Spinach chloroplasts. Plant Cell Physiol., 22: 867-880.
CrossRefDirect Link - Bohnert, H.J., D.E. Nelson and R.G. Jensen, 1995. Adaptations to environmental stresses. Plant Cell., 7: 1099-1111.
CrossRefPubMedDirect Link - Delauney, A.J. and D.P.S. Verma, 1993. Proline biosynthesis and osmoregulation in plants. Plant J., 4: 215-223.
CrossRef - Ehsanpour, A.A. and N. Fatahian, 2003. Effect of salt and proline on Medicago sativa callus. Plant Cell Tissue Organ. Cult., 73: 53-56.
Direct Link - Zhu, J.K., 2002. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol., 53: 247-273.
CrossRefPubMedDirect Link - Apel, K. and H. Hirt, 2004. Reactive oxygen species: Metabolism, oxidative stress and signal transduction. Annu. Rev. Plant Biol., 55: 373-399.
CrossRefPubMedDirect Link - Mudgal, V., N. Madaan and A. Mudgal, 2010. Biochemical mechanisms of salt tolerance in plants: A review. Int. J. Bot., 6: 136-143.
CrossRefDirect Link - Nojavan, A.M. and M. Khorshidi, 2006. An investigation of vanillin imposed oxidative stress in corn (Zea mays L.) and the activities of antioxidative enzymes. Pak. J. Biol. Sci., 9: 34-38.
CrossRefDirect Link - Bhardwaj, R., N. Arora, P. Sharma and H.K. Arora, 2007. Effects of 28-homobrassinolide on seedling growth, lipid peroxidation and antioxidative enzyme activities under nickel stress in seedlings of Zea mays L. Asian J. Plant Sci., 6: 765-772.
CrossRefDirect Link