
Shukdeb Sen
Department of Biology, School of Science, Engineering and Mathematics,Bethune Cookman University, 640 Dr. Mary McLeod Bethune Boulevard,Daytona Beach, Florida 32114-3099, USA
American Journal of Plant Physiology
Year: 2010 | Volume: 5 | Issue: 3 | Page No.: 122-132







ABSTRACT
Seed germination is a complex process involving various physical and biochemical cues such as water, light and phytohormones. This study focused on the cross talk between Nitric Oxide (NO) and hemoglobin in the regulation of the seed germination process in wheat (Triticum durum). The levels of NO, hemoglobin, catalase and S-nitrosylated proteins increased during sensu stricto germination but rapidly dropped off within 48 h of imbibition. At least thirteen S-nitrosylated proteins were detected by using modified Biotin switch method. S-nitrosylated protein bands with molecular mass 93, 80, 71, 52, 45.7, 38, 32, 27.5, 25, 20, 15, 12 and 10 kD were observed during the 24 h period. The levels of these proteins decreased during 48 h. The NO scavenger c-PTIO-treated tissue exhibited suppressed levels of synthesis for 93, 80 and 71 kD proteins during 24 h when radicles emerged. It drastically lowered the NO levels in the germinating seeds but did not inhibit the germination process, suggesting the possibility that NO is not directly involved in the regulation process. The S-nitrosylation process is the switch which regulates the seed germination process. Ideas along these lines are discussed in the study.
PDF Abstract XML References Citation
How to cite this article
Shukdeb Sen, 2010. S-Nitrosylation Process Acts as a Regulatory Switch for Seed Germination in Wheat. American Journal of Plant Physiology, 5: 122-132.
DOI: 10.3923/ajpp.2010.122.132
URL: https://scialert.net/abstract/?doi=ajpp.2010.122.132
DOI: 10.3923/ajpp.2010.122.132
URL: https://scialert.net/abstract/?doi=ajpp.2010.122.132
INTRODUCTION
Seed germination is a complex process where the emergence of radicle from the structures surrounding the embryo after water uptake by the seeds is considered as the completion of germination. The events that are part of this process involve various physical and biochemical interactions. The researcher proposed that Nitric Oxide (NO) in conjunction with phytochrome regulate the process of seed germination (Sen and Cheema, 1995). After the publication of this report, a large number of papers followed documenting the presence of nitric oxide in the plants and various regulatory roles played by the No. Exogenous application of NO donors have significantly enhanced the germination and/or break dormancy in seeds (Beligni and Lamattina, 2000; Bethke et al., 2004a; Sarath et al., 2006). Seed dormancy can be effectively broken by the addition of cyanide, nitrite, nitrate and NO, which resulted in higher rates of germination in Arabidopsis seeds (Bethke et al., 2006).
The 2-Phenyl-4,4,5,5,-tetramethylimidazoline-1-oxyl 3-oxide (PTIO), is a potent and specific chemical scavenger of NO (Goldstein et al., 2003) and it has been extensively used to block NO-responsive effects in various plant tissues (Lamattina et al., 2003). In warm-season C4 grass seeds, PTIO blocked NO-responsive stimulation of germination, indicating that adequate levels of endogenous NO were required for germination (Sarath et al., 2006).
Nitric Oxide (NO) can be produced in plants by non-enzymatic as well as enzymatic pathway systems. Plant tissues synthesize NO via the non-enzymatic reduction of apoplastic nitrite. The NO is produced rapidly when nitrite was added to the medium in the barley (Hordeum vulgare) aleurone layers. The NO production requires an acid apoplast and phenolic compounds in the medium could increase NO production (Bethke et al., 2004b). The NO-producing enzymes identified in plants are nitrate reductase, (Yamasaki and Sakihama, 2000; Rockel et al., 2002; Desikan et al., 2002; Toon et al., 2001). Nitrate reductase activity increases during anoxic condition and a concomitant release of nitrite by the tomato roots in the culture medium (Allegre et al., 2004). The L-arginine-dependent Nitric Oxide Synthase (NOS) enzyme in plants have been reported in olive leaves (Valderrama et al., 2007) and sunflower hypocotyls (Chaki et al., 2009). Evidences indicate that NO can also be generated by other enzymes such as xanthine oxidoreductase, peroxidase, cytochrome P450 and some heme proteins (Del Rio et al., 2004).
Hemoglobins are ubiquitous in all living systems. Three distinct types of hemoglobin have been found in plants and they are classified as symbiotic, non-symbiotic and truncated hemoglobin. The symbiotic hemoglobins are found in the nodules of leguminous plants and they regulate oxygen supply to the nitrogen-fixing bacteria (Appleby, 1992). The non-symbiotic hemoglobin (nsHbs) can be categorized into two groups based on their oxygen-binding properties. The class 1 hemoglobin is stress-induced hemoglobin that is activated during hypoxic stress and has different oxygen-binding properties whereas; class 2 type hemoglobin has similar oxygen-binding properties as symbiotic hemoglobin (Dordas et al., 2003). The truncated hemoglobin share some characteristics with non-symbiotic hemoglobin but the functions of this hemoglobin is unknown at this time (Watts et al., 2001).
The S-nitrosylation is a process where the covalent attachment of a nitrogen monoxide group to the thiol side chain of the amino acid cysteine occurs and it has emerged as an important mechanism by which proteins undergo post-translation modification. In animal system, various S-nitrosylated proteins have been extensively investigated. These proteins control various processes through the redox-based regulation mechanisms. The S-nitrosylation of a cysteine residue of the beta globin chain of hemoglobin has been reported recently. In birds and mammals these cysteine residues have been conserved and can be S-nitrosylated by NO to form S-nitrosothiol-hemoglobin (SNO-Hb). In human beta chain the cysteine residue is positioned at 93 (Gow and Stamler, 1998; Stamler et al., 1997b). According to these reports, the S-nitrosothiols do not react with Fe (II)-Hb and thus SNO-Hb can deliver the bioactive NO to the tissue and can activate the guanylyl cyclase pathway to render transduction cascade. In plants, at least three proteins have been experimentally shown to be regulated by S-nitrosylation. Arabidopsis hemoglobin 1 can scavenge NO through the formation of S-nitroso-hemoglobin (Perazzolli et al., 2004). Glyceraldehyde 3-phosphate dehydrogenase (Mohr et al., 1999) and methionine adenosyltransferase (Lindermayr et al., 2005) have been regulated by NO via S-nitrosylation.
In durum wheat (Triticum durum Desf.), the researcher observed that the Nitric Oxide (NO) scavenger 2-(4-carboxyphenyl)-4,4,5, 5-tetramethylimidazoline-1-oxyl-3 oxide (cPTIO) neither induces dormancy nor suppresses the seed germination. Instead, it stimulated the germination process. This unexpected finding lead us to propose that NO regulates the process of germination indirectly in durum wheat seeds. We hypothesize that NO exerts its influence through the chemical modification of various targets such as the covalent modification of amino acid cysteine (S-nitrosylation). The rationale behind this idea is supported by the fact that cPTIO is an effective scavenger of NO, as such, exposure to a high concentration of cPTIO (1 mM) should scavenge all of the free NO available in plant tissue and eliminate the direct participation of NO in the regulation of seed germination. S-nitrosylation of the target proteins, however, could escape the scavenging of NO in the tissue and subsequently, at an appropriate physiological condition, can deliver NO to the tissue as needed for the signaling cascade.
In this study, we have evaluated the influence of cross-talk between hemoglobin and nitric oxide during the seed germination process. Furthermore, we have presented a hypothetical model to explain how these important biomarkers influence the processes of seed germination in durum wheat.
MATERIALS AND METHODS
Carboxy PTIO potassium salt (2-(4-carboxy phenyl)-4,4,5,5--tetramethylimidazoline-1-oxyl-3 oxide (cPTIO) was obtained from MP Biomedicals, Solon, Ohio. Nitric oxide assay kits were purchased from Assay Designs, Ann Arbor, Michigan. All electrophoretic supplies such as polyacrylamide, SDS, ammonium persulfate, protein standards, nitrocellulose papers, Alkaline phosphatase Immun-Blot assay kits, buffers and other chemicals were purchased from Bio-Rad Laboratories, Hercules, California. Protease inhibitor cocktail, Sodium nitroprusside (SNP) and other chemicals were purchased from Sigma chemicals, St. Louise, Missouri. Wheat seeds (Triticum durum) were procured from Great Harvest Organics, Atlanta, Indiana. Polyclonal Anti rice hemoglobin (Hb1) antibodies received from Dr. G. Sarath, University of Nebraska-Lincoln, Nebraska. Anti SNO-Cys antibody was obtained from Alpha Diagnostic Company, Texas. S-nitrosylated protein detection kits were purchased from Cayman chemical company, Ann Arbor, Michigan. Prepared gradient gels were purchased from Jules, Inc, Milford, Cincinnati. This study was conducted during June 2006 through July 2009.
Extraction of Embryonic Axes from Wheat Seeds
Wheat (Triticum durum) seeds were soaked in 10% bleach for 5 min and then rinsed with sterile distilled water for five to seven times. These surface sterilized seeds were placed on water saturated sterilized filter papers for 4 h in dark. After 4 h of imbibition, seeds were treated with various reagents. Various concentrations of sodium nitroprusside (SNP) and c-PTIO [100 μM, 1 mM and 10 mm] were sprayed once to see the effects of SNP and c-PTIO. In some experiments, 3 h after SNP spray (100 μM c-PTIO) containing solution was sprayed. All untreated control and treated seeds were kept in dark at room temperature (24°C). Embryonic axes were excised from germinating seeds at different time period ranging from 12, 18, 24, 36 and 48 h after imbibition. Embryonic axes were excised in such a way that the most endosperm and other parts of the caryopsis were discarded.
Preparation for Protein Extracts for Gel Electrophoresis
One hundred embryonic axes were excised from each batch and homogenized in 750 μL of extraction buffer containing the protease inhibitor cocktail from Sigma (125 mM Tris, pH 6.8; 2% SDS, 20% glycerol) with ice-cold mortar and pestle Homogenized tissue was centrifuged in a Fisher microcentrifuge for 5 min and supernatants were filtered through Millipore Ultrafree-MC centrifugal filtration device. Filtrates were collected from each group of samples and 5% 2-mercaptoethanol was added to each sample and boiled for 5 min. These samples were used for gel electrophoresis experiments.
Extraction of S-nitrosylated Proteins from Embryonic Axes
The S-nitrosylated proteins were extracted by the method described in the S-nitrosylated protein detection assay kit (Cayman Chemicals, Ann Arbor, Michigan) which is based on modified Biotin switch method of Jaffrey et al. (2001). Briefly, the embryonic axes (0.5-1.0 g) were homogenized in 1 mL WASH buffer and centrifuged in a microcentrifuge for 5 min to remove the particulates. The supernatants (500 μL) were collected and mixed with equal volume of WASH buffer and then centrifuged to pellet the tissue (500x g, 5 min). Then to each cell pellet 5 volumes (up to 0.5 mL total volume) of Buffer A containing Blocking reagent, was added. Samples were incubated for 30 min at 4°C on a shaker with gentle agitation. Lysates were collected by centrifugation of the samples for 10 min at 4°C with a tabletop microcentrifuge. Supernatants were transferred into separate cold, 15 mL conical polypropylene tubes and 4 volumes of ice-cold acetone to each sample were added. All samples were mixed thoroughly by inversion followed by incubation at -20°C for 1 h. Samples were removed from -20°C and centrifuged at 3000x g for 10 min at 4°C to produce compact protein pellets. Carefully acetone was decanted from each sample pellets. The samples were resuspended with 0.5 mL of buffer B containing reducing and S-nitrosylation Labeling reagents. Samples were mixed well by inversion and incubated for 1 h at room temperature. All these steps were carried out in dark.
Four volumes (2 mL) of ice-cold acetone was added to each sample, mixed each thoroughly by inversion followed by incubation at -20°C for 1 h. Samples from -20°C were removed and centrifuged at 3000x g for 10 min at 4°C. Sample pellets were collected and each pellet was resuspended in a minimal volume (100-200 μL) of cold WASH buffer. Protein concentrations were determined of each samples and the remaining samples were stored at -20°C for further analysis.
Nitric Oxide Assay
Embryonic axes (0.5-1.0 g) from control and various treated seeds were homogenized with 1 mL of phosphate buffered saline (10 mM sodium phosphate, 150 mM NaCl, pH 7.4) and centrifuged at 13,000 rpm for 5 min to remove particles. The supernatants were collected and centrifuged again at 13,000 rpm using Microcon Ultracel YM-10 (Millipore Corporation) filtration device for 40 min. Filtrates were collected and stored in refrigerator for future use. These filtrates were used to measure total nitrate/nitrite levels in the sample by assay designs total nitric oxide assay kits.
Polyacrylamide Gel Electrophoresis and Western Blotting Analysis
Treated and non-treated samples were subjected to SDS gel electrophoresis as described (Sen and Cheema, 1995). The mini gels were transferred immediately on nitrocellulose membrane. Protein transfer was carried out at 30 volts, 90 mA for 18 h at room temperature in a Hoefer Mighty small transphor unit. After completion of transfer, the membranes were washed in TTBS (20 mM Tris, 500 mM NaCl, 0.05% Tween-20, pH 7.5) throughly and blocked by 5% non-fat dry milk for 2 h. After TTBS wash, the membranes were incubated with rice anti-hemoglobin antibody (generous gift from Dr. Gautam Sarath, USDA-ARS, University of Nebraska, Lincoln, Nebraska) for 2 h in room temperature. Blots were washed with TBS-Tween buffer and then incubated with a 1/3000 dilution of the anti-rabbit IgG conjugated with alkaline phosphatase for 2 h at room temperature. After washing with TBS-Tween, blots were developed using the chromogenic mix nitroblue tetrazolium/5-bromo-4-chloro-3-indolyl phosphate according to the direction given in Amplified alkaline phosphatase Immun-Blot assay kit (Bio-Rad Corporation).
For the identification of S-nitrosylated proteins, slight modification of the blotting procedures was undertaken. For blocking step, 2% bovine serum albumin in PBS was utilized instead of dry milk. Anti biotin antibody conjugated with HRP (Bethyl Laboratories, Texas) was used. For the color development we used 4-chloro-1-naphhol (4CN) to identify S-nitrosylated proteins. For catalase studies, rabbit polyclonal anti catalase antibody was used. Anti-catalase antibody was obtained from Rockland, Pennsylvania.
RESULTS
Sequence Profiles of Non-Symbiotic Hemoglobin
Analysis of sequence profile searches revealed that non-symbiotic hemoglobin contains a conserved cysteine at position# 79 in all groups of plants (Fig. 1). This cysteine might be a target for S-nitrosylation that would result S-nitrosocysteine (SNO-Cys). This could act as a reservoir of NO for future use by the plant tissue. Leg-hemoglobins do not have cysteines in them. The acid-base catalysis of nitrosylation explained how some proteins containing a thiol reaction site have a C(D,E) sequence-an acidic amino acid following cysteine This cysteine is bioregulatory targets of nitrogen oxides (Stamler, 1994; Stamler et al., 1997a). The non-symbiotic hemoglobin contain amino acid sequence VMTCE/D where, C(D,E) sequence-an acidic amino acid following cysteine is present. This cysteine may be S-nitrosylated in hemoglobin forming SNO-Cys Hb.
When S-nitrosoglutathione (GSNO) and L-cysteine is mixed in a phosphate buffer (pH 7.0 or 7.4), the degradation of GSNO takes place within 3 h. This might suggest that the degradation may be due to direct release of NO from GSNO, which is enhanced by L-cysteine. Furthermore, it is possible that trans-nitrosation of the NO-group from GSNO to L-Cysteine have occurred (Kuo et al., 2003, 2005). This information supports the idea that cysteine residue in hemoglobin might form SNO-Cys.
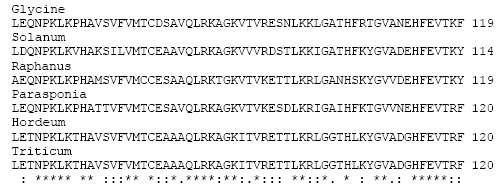 | |
| Fig. 1: | Multiple sequence alignment of the non-symbiotic hemoglobin was constructed by using ClustalW program. Partial amino acid sequences are presented here to show the segments where cysteine is conserved. Accession Numbers are given as follows: Hordeum Q42831, Triticum AAN85432, Raphanus AAP37043, Glycine AAA97887, Solanum AAN85431 and Parasponia AAB86653 |
Tissue Levels of NO in Embryonic Axes
Levels of nitric oxide measured by Griess reactions from germinated wheat seeds are shown in Table 1. The results indicate that there is a copious amount of nitric oxide is produced in the germinating seeds after 24 h of H2O imbibition. The levels of NO drops as embryonic axes grow.
| Table 1: | Levels of nitric oxide observed in control and SNP-treated embryonic axes of germinating wheat seeds per gram of tissue |
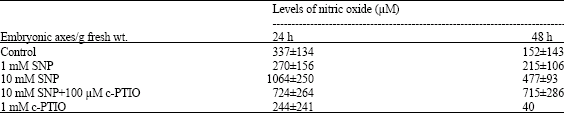 | |
| *Data presented here reflect the mean values±SD of four replicates with the exception of 48 h c-PTIO treated samples | |
The NO donor sodium nitroprusside (SNP) increased the NO levels in treated seeds. The NO scavenger 2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide (c-PTIO) treatment reduced the level of NO drastically. Germinated seeds exposed to 100 μM SNP produced similar amount of NO per gram of fresh embryonic axes tissue. A 2-fold increase in the levels of NO in seeds treated with 10 mM SNP was observed. Similarly, combination of 10 mM SNP and 100 μM c-PTIO lowered drastically the levels of No. The seeds treated with 1 mM c-PTIO alone resulted lowest levels of NO indicating active scavenging/suppression of NO synthesis in the tissue. However, it did not inhibit the germination process because 80% seeds germinated. Higher concentration of SNP (10 mM), drastically suppressed the germination (7-10%) and blocked root differentiation.
Tissue Levels of Hemoglobin by Western Blot Analysis
Tissue hemoglobin levels were measured by western blot analysis (Fig. 2). After 24 h of water imbibitions, the emergence of embryonic root (radicle) take place indicating the completion of sensu stricto germination. At this juncture, extensive synthesis of hemoglobin occurred in the tissue that last for a short period of time. After 48 h of imbibitions, the levels of hemoglobin decreased drastically and completely eliminated at 72 h (Fig. 3).
 | |
| Fig. 2: | Western blot of proteins from germinating wheat seeds after 24 h of imbibitions. Each lane consisted of 80 mg crude protein extracts and the blot was probed with polyclonal rice Hb antibody. Lane 1: Rice Hb-1, Lane 2: Blank, Lane 3: 10 Mm SNP, Lane 4: 1 mM c-PTIO , Lane 5: 10 mM SNP + 100 mM c-PTIO, Lane 6: 10 mM SNP, Lane 7: 1 mM SNP, Lane 8: Control, Lane 9: Blank, Lane 10: Protein standard |
 | |
| Fig. 3: | Western blot of proteins from germinating wheat seeds after 48 h of imbibition. Each lane consisted of 80 μg crude protein extracts and the blot was probed with polyclonal rice Hb-1 antibody. Lane 1: Control, Lane 2: 100 μM SNP, Lane 3: 1mM SNP, Lane 4: 10 mM SNP, Lane 5: 10 mM SNP+100 μM c-PTIO, Lane 6: 1 mM c-PTIO, Lane 7: Blank, Lane 8: Rice Hb-1 protein |
S-Nitrosylated Protein Profile
Biotin switch method of Jaffrey et al. (2001) was used to visualize S-nitrosylated proteins by western blot analysis (Fig. 4). This method blocks free SH groups first and any S-NO bonds present in the samples are then cleaved. Biotinylation of the newly formed SH groups provide the basis for visualization using streptavidin-based colorimetric detection. Jaffrey et al. (2001) detected 13 proteins bands. The method involves the substitution of a biotin group at every cysteine sulfur residues modified by nitrosylation. The biotinylated proteins are then detected by immunoblotting using anti-biotin antibody. Immunoreactive bands corresponding to 10, 12, 15, 20, 25, 27.5, 32, 38, 45.7, 52, 71, 80 and 93 kDa were observed in germinating wheat seeds. Control and sodium nitroprusside (SNP)-treated seeds showed 93, 80, 71, 52, 27.5, 25, 20, 15 and 10 kDa protein bands indicating that these proteins are expressed during 24 h after water imbibition, when radicle emergence is completed.
Seeds treated with NO scavenger 2-(4-carboxyphenyl)-4,4,5, 5-tetramethylimidazoline-1-oxyl-3 oxide (c-PTIO) showed protein bands with molecular weight 52, 27.5, 15 and 10 kDa indicating that these proteins are expressed only during 24 h. Bands with 93, 80 and 71 kD were less prominent with c-PTIO treatment, indicating suppressed synthesis of these proteins.
During 48 h, 93, 80, 71, 52 kDa protein levels are reduced drastically indicating that these groups of proteins are expressed during the radicle emergence stages and are essential in the early phases of seed germination. It appears that these proteins may not be essential during post germination stages. It is important to note that 10 mM SNP strongly induce the expression of 52 kDa protein and it retains its activity even after 48 h. This observation suggests that 52 kDa protein is an important protein that play a very important role in pre and post germination stages in wheat. The 10 mM SNP treated seeds show poor germination and inhibit root differentiation.
 | |
| Fig. 4: | Western blot images of proteins from germinating wheat seeds after 24 and 48 h of imbibition in various solutions. Each lane consisted of 60 μg of crude protein extracts and the blot was probed with polyclonal anti-biotin antibody. Lane 1: 1 mM c-PTIO [48 h], Lane 2: 10 mM SNP [48 h], Lane 3: 1 mM SNP [48 h], Lane 4: Control [48 h], Lane 5: 10-3M 2,4,5-T [24 h], Lane 6: 1 mM c-PTIO [24 h], Lane 7: 10 mM SNP [24 h], Lane 8: 1 mM SNP [24 h], Lane 9: Control [24 h], Lane 10: Protein standard |
DISCUSSION
Growth analysis revealed that in both control and 100 μM SNP treatment 95% seeds germinated after one-day incubation but at higher concentration (10 mM) SNP only 5-10% seeds germinated indicating severe inhibitory effects on germination. Furthermore, those seeds that were germinated in 10 mM SNP treatment, showed stunted growth in the primary and seminal roots indicating an inhibitory influence in the root differentiation process. The coleoptile growth was normal in 10 mM SNP treatment. Ten millimolar SNP-treated tissue exhibited three-fold more NO than control tissue (Table 1), suggesting that excess NO suppressed the normal root differentiation and directly regulate the root differentiation process. The cPTIO-treatment on the other hand, showed 90% germination as well as normal primary and seminal root growth. The coleoptile growth was also normal. The levels of NO reduced drastically in 1 mM cPTIO treated tissue but it did not affect the germination process. This result contradicts other published reports where, cPTIO inhibits the germination by initiating dormancy. This contradictory findings can be resolved through the hypothesis that NO influence germination via S-nitrosylation.
The data presented here indicate that the levels of NO, class-1 non-symbiotic hemoglobin (nsHb) and S-nitrosylated proteins increased during early stages of seed germination (24 h), whereas levels of each of these biomarkers dropped off during 48 h. Similarly, catalase levels increased during 24 h of growth, which then dropped off significantly during 48 h. Seed dormancy inducer c-PTIO, effectively scavenge NO, but it does not inhibit the seed germination process. At a relatively high concentration (1 mM) of c-PTIO, the levels of NO reduced drastically but did not inhibit germination. This observation raises the question, is NO necessary for the germination process? Sodium nitroprusside (SNP), a NO donor reduces seed dormancy in Arabidopsis, as well as other seeds. Treatment with high concentration of SNP (10 mM) increased NO levels, but when exposed with 100 μM c-PTIO, it reduced the NO but did not increase the rate of seed germination.
To answer the question, whether NO is necessary for germination, the following facts should be emphasized. The basal levels of NO remain same in all tissue regardless of treatment (Table 1) during the 24 h period. The effects of c-PTIO at this early stage is blocked by a mechanism yet to be discovered, or NO might have reacted with sulfhydryl group (s) of hemoglobin and other proteins, yielding S-nitrosothiols which are unaffected by c-PTIO. However, the NO levels drop off drastically in all treated tissue during 48 h. Reduction of NO levels correlates well with the reduction of S-nitrosylated proteins during 48 h when the completion of germination occurred. However, moderate levels of S-nitrosylated proteins remain, indicating some residual NO might have been present in the tissue during post-germination period.
During germination, the activation of NO and hemoglobin indicate that there is a regulatory cross-talk between these two agents that control the early phases of the germination process. Hemoglobin plays a dual role in this regulatory switch. On one hand, it can scavenge NO to regulate the NO levels; on the other hand, it could play a significant role as the substrate for S-nitrosylation process. Furthermore, hemoglobin can act as a source for oxygen by autoxidation in hypoxic environment. Data presented here as well as by Ishibashi et al. (2008) who indicated that activation of NO, H2O2 and catalase occurred during germination process. A similar activation of these agents was also reported during the disease response process. This parallel relationship points to the fact that during germination, seeds are exposed to an environment where various disease causing microbial agents exists and the seeds are vulnerable to these pathogenic attack. It has been documented that NO play a major role in the regulation of Hypersensitive Reaction (HR), which modulates the activities of Systemic Acquired Resistance (SAR) and defense-related genes (Delledonne et al., 1998; Wendehenne et al., 2004; Zaninotto et al., 2006). Similarly, S-nitrosothiols also play a key role in plant disease resistance (Feechan et al., 2005). For this reason, it could be suggested that a part of increased NO levels observed during sensu stricto germination is due to preventive action against disease causing agents rather than direct influence on the germination process. In durum wheat, c-PTIO did not inhibit germination nor did it extend dormancy. This observation suggests strongly that seeds store S-nitrosylated proteins which could be cleaved during imbibition when pH level drops and donate NO to the germinating seeds.
This observation suit well to the hypothesis that NO does not activate germination process directly, instead, NO exerts its influence through S-nitrosylation process. The NO can easily modify various proteins through S-nitrosylation, forming S-nitrosothiols which are short-lived and controlled directly by the redox state of the cell. This mechanism of protein S-nitrosylation can act as a switch in the regulation of various biochemical pathways that controls the seed germination process. Characterization and identification of S-nitrosylated proteins that were observed at different stages of growth during the seed germination process is necessary so that we can get a better picture of the regulatory roles of these S-nitrosylated proteins.
ACKNOWLEDGMENTS
This study was supported in part by the National Science foundation (NSF-UP), grant# HRD-0310317. Bethune Cookman University Research Foundation also supported this project. Author is grateful to Dr. Gautam Sarath, University of Nebraska-Lincoln, Nebraska for sharing rice hemoglobin antibody.
REFERENCES
- Allegre, A., J. Silvestre, P. Morard, J. Kallerhoff and E. Pinelli, 2004. Nitrate reductase regulation in tomato roots by exogenous nitrate: A possible role in tolerance to long-term root anoxia. J. Exp. Bot., 55: 2625-2634.
CrossRefDirect Link - Bethke, P.C., M.R. Badger and R.L. Jones, 2004. Apoplastic synthesis of nitric oxide by plant tissues. Plant Cell, 16: 332-341.
Direct Link - Bethke, P.C., F. Gubler, J.V. Jacobson and R.L. Jones, 2004. Dormancy of Arabidopsis seeds and barley grains can be broken by nitric oxide. Planta, 219: 847-855.
CrossRefDirect Link - Bethke, P.C., I.G. Libourel, V. Reinohl and R.L. Jones, 2006. Sodium nitroprusside, cyanide, nitrite and nitrate break Arabidopsis seed dormancy in a nitric oxide-dependent manner. Planta, 223: 805-812.
CrossRefDirect Link - Chaki, M., A.M. Ferna'ndez-Ocana, R. Valderrama, A. Carreras and F.J. Esteban et al., 2009. Involvement of reactive nitrogen and oxygen species (RNS and ROS) in sunflower-mildew interactions. Plant Cell Physiol., 50: 265-279.
CrossRefDirect Link - Del Rio, L.A., F.J. Corpas and J.B. Barroso, 2004. Nitric oxide and nitric oxide synthase activity in plants. Phytochemistry, 65: 783-792.
Direct Link - Delledonne, M., Y. Xia, R.A. Dixon and C. Lamb, 1998. Nitric oxide functions as a signal in plant disease resistance. Nature, 394: 585-588.
Direct Link - Desikan, R., R. Griffiths, J. Hancock and S. Neill, 2002. A new role for an old enzyme: Nitrate reductase-mediated nitric oxide generation is required for abscisic acid-induced stomatal closure in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA., 99: 16314-16318.
CrossRefDirect Link - Dordas, C., R. Jean and R.D. Hill, 2003. Plant haemoglobins, nitric oxide and hypoxic stress. Ann. Bot., 91: 173-178.
Direct Link - Feechan, A., E. Kwon, B.W. Wang, Y. Wang, J.A. Pallas and G.J. Loake, 2005. A central role for S-nitrosothiols in plant disease resistance. Proc. Nat. Acad. Sci. USA., 102: 8054-8059.
PubMedDirect Link - Goldstein, S., A. Russo and A. Samuni, 2003. Reactions of PTIO and carboxy-PTIO with *NO, *NO2 and O2-*. J. Biol. Chem., 278: 50949-50955.
PubMedDirect Link - Gow, A.J. and J.S. Stamler, 1998. Reactions between nitric oxide and haemoglobin under physiological conditions. Nature, 391: 169-173.
PubMedDirect Link - Ishibashi, Y., K. Yamamoto, T. Tawaratsumida, T. Yuasa and M. Iwaya-Inoue, 2008. Hydrogen peroxide scavenging regulates germination ability during wheat (Triticum aestivum L.) seed maturation. Plant Signaling Behav., 3: 183-188.
PubMedDirect Link - Jaffrey, S.R., H. Erdjument-Bromage, C.D. Ferris, P. Tempst and S.H. Snyder, 2001. Protein S-nitrosylation: A physiological signal for neuronal nitric oxide. Nat. Cell Biol., 3: 193-197.
CrossRefDirect Link - Kuo, W.N., J.M. Robinson and J. Nibbs, 2003. Nitrosation of cysteine and reduced glutathione by nitrite at physiological pH. Front. Biosci. 8: 62-69.
PubMedDirect Link - Lamattina, L., C. Garcia-Mata, M. Graziano and G. Pagnussat, 2003. Nitric oxide: The versatility of an extensive signal molecule. Annu. Rev. Plant Biol., 54: 109-136.
PubMed - Lindermayr, C., G. Saalbach and J. Durner, 2005. Proteomic identification of S-nitrosylated proteins I Arabidopsis. Plant Physiol., 137: 921-930.
CrossRefDirect Link - Mohr, S., H. Hallak, A. Boitte, E.G. Lapetina and B. Brune, 1999. Nitric oxide induced S-glutathionylation and inactivation of glyceraldehydes 3-phosphate dehydrogenase. J. Biol. Chem., 274: 9427-9430.
CrossRefDirect Link - Perazzolli, M., P. Dominici, M Romero-Puertas, E. Zago and J. Zeier et al., 2004. Arabidopsis non-symbiotic hemoglobin Ahb1 modulates nitric oxide bioactivity. Plant Cell, 16: 2785-2794.
CrossRefDirect Link - Rockel, P., F. Strube, A. Rockel, J. Wildt and W.M. Kaiser, 2002. Regulation of nitric oxide (NO) production by plant nitrate reductase in vivo and in vitro. J. Exp. Bot., 53: 103-110.
Direct Link - Sarath, G., P.C. Bethke, R. Jones, L.M. Baird, G. Hou and R.B. Mitchell, 2006. Nitric oxide accelerates seed germination in warm-season grasses. Planta, 223: 1154-1164.
CrossRefDirect Link - Sen, S. and I. Cheema, 1995. Nitric oxide synthase and calmodulin immunoreactivity in plant embryonic tissue. Biochem. Arch., 11: 221-227.
Direct Link - Stamler, J.S., 1994. Redox signaling: Nitrosylation and related target interactions of nitric oxide. Cell, 78: 931-936.
PubMed - Stamler, J.S., L. Jia, J.P. Eu, T.J. McMahon and I.T. Demchenko et al., 1997. Blood flow regulation by S-nitrosohemoglobin in the physiological oxygen gradient. Science, 276: 2034-2037.
CrossRefDirect Link - Stamler, J.S., E.J. Toone, S.A. Lipton and N.J. Sucher, 1997. (S)NO signals: Translocation, regulation and a concensus motif. Neuron, 18: 691-696.
PubMedDirect Link - Toon, N.N., A. Holk and G.F. Scherer, 2001. Rapid increase of NO release in plant cell cultures induced by cytokinin. FEBS Lett., 509: 174-176.
PubMedDirect Link - Valderrama, R., F.J. Corpas, A. Carreras, F. Luque and A. Fernandez-Ocana et al., 2007. Nitrosative stress in plants. FEBS Lett., 581: 453-461.
Direct Link - Watts, R.A., P.W. Hunt, A.N. Hvitved, M.S. Hargrove, W.J. Peacock and E.S. Dennis, 2001. A hemoglobin from plants homologous to truncated hemoglobin of microorganisms. Proc. Natl. Acad. Sci. USA., 98: 10119-10124.
CrossRefDirect Link - Wendehenne, D., J. Durner and D.F. Klessig, 2004. Nitric oxide: A new player in plant signaling and defense responses. Curr. Opin. Plant Biol., 7: 449-455.
PubMedDirect Link - Yamasaki, H. and Y. Sakihama, 2000. Simultaneous production of nitric oxide and peroxynitrite by plant nitrate reductase in vitro evidence for the NR-dependent formation of active nitrogen species. FEBS Lett., 468: 89-92.
Direct Link - Zaninotto, F., S. La Camera, S. Polverari and M. Delledonne, 2006. Cross talk between reactive nitrogen and oxygen species during hypersensitive disease resistance response. Plant Physiol., 141: 379-383.
Direct Link