
S. Rezvani
Young Researchers Club, Department of Plant Biology
LiveDNA: 98.13110
S. Shariati
Young Researchers Club, Department of Chemistry, Islamic Azad University, Rasht Branch, Rasht, Guilan, Iran
American Journal of Plant Physiology
Year: 2010 | Volume: 5 | Issue: 1 | Page No.: 1-6







ABSTRACT
In this study, one Arabidopsis thaliana L. in the stress conditions, growthing was analysed for the presence of trehalose. Using as anion-exchange high performance liquid chromatography (HPLC) analysis, trehalose was 0.63 mg-1 wd highest in flower and root 0.4 mg-1 wd. Furthermore, trehalose metabolizing enzymes, Trehalose-6-Phosphate Synthase (TPS) and trehalase enzyme activities were measured in flower and root. The TPS activity sharply increased under stress conditions growth. In addition, enzymes and metabolism trehalose 6-phosphate synthase enzyme activity Trehalase TPS and also review and determined that TPS activity in terms of drought growth increased the final conclusion was that some plants in stress conditions trehalose synthesis are also clear that this sugar can prevent the accumulation of proteins such as β-Amyloidal (in Alzheimer's disease), Huntington (Huntington disease) and Prion (Krotzfild-Jakob disease). Thus, this can be as sugar drug used against these diseases.
PDF Abstract XML References Citation
How to cite this article
S. Rezvani and S. Shariati, 2010. Effect of Trehalose Arabidopsis thaliana L. on Huntington's Disease. American Journal of Plant Physiology, 5: 1-6.
DOI: 10.3923/ajpp.2010.1.6
URL: https://scialert.net/abstract/?doi=ajpp.2010.1.6
DOI: 10.3923/ajpp.2010.1.6
URL: https://scialert.net/abstract/?doi=ajpp.2010.1.6
INTRODUCTION
Trehalose is a disaccharide that occurs naturally in insects, plants, fungi and bacteria. The major dietary source is mushrooms. Trehalose is used in bakery goods, beverages, confectionery, fruit jam, breakfast cereals, rice and noodles as a texturizer, stabilizer, humectant, or formulation aid with a low sweetening intensity. Trehalose is a naturally occurring disaccharide with known protein and membrane stabilizing capability. Although, trehalose absorption in humans has not been well studied, a small fraction (0.5%) is likely to be absorbed by passive diffusion, as has been demonstrated for other dissacharides (Van Elburg et al., 1995). In mammalian cell culture, trehalose is moved from the extracellular to intracellular compartment via a fluid phase endocytotic mechanism and is dependent on extracellular concentration (Oliver et al., 2004).
The mechanisms by which trehalose metabolism alters plant development are largely unknown. Trehalose itself could affect development by acting as signal molecule in carbohydrate metabolism. For example, trehalose induces enzymes of fructan synthesis in barley (Wagner et al., 1986; Mulleret al., 2000) and Sucrose synthase activity in soybean (Muller et al., 1998). In general, sugars such as Sucrose and Glucose act as signals in the regulation of gene expression (Koch, 1996). Whereas the expression of several source-specific genes is probably regulated by hexoses in a hexokinase-dependent signaling pathway (Jang and Sheen, 1994; Jang et al., 1997; Dai et al., 1999), the regulation of the expression of some other genes appears to be directly mediated by Sucrose without prior cleavage to hexoses (Yokoyama et al., 1994; Chiou and Bush, 1998; Rook et al., 1998). It is conceivable that trehalose, which is structurally similar to Sucrose, might act as an analog of Sucrose in sugar sensing. In addition to the trehalose that may be produced by the plants themselves, plants are also exposed to trehalose formed by microorganisms in mutualistic, as well as in pathogenic interactions. Trehalose formed by rhizobia during nodulation appears to have a strong effect on the carbohydrate contents in root nodules (Muller et al., 1995b). It is possible that trehalose-producing plant symbionts and/or pathogens can exploit the effects of trehalose to gain control over the sugar-sensing system of the plant. If this is the case, the trehalose-degrading enzyme trehalase, which is widespread among higher plants and is found in multiple tissues, may provide a safeguard against potentially deleterious effects of trehalose on carbohydrate allocation in plant-microbe interactions (Muller et al., 1995a; Aeschbacher et al., 1999).
Trehalose is thought to act by altering or replacing the water shell that surrounds lipid and protein macromolecules (Colaco et al., 1995). Trehalose is a naturally occurring disaccharide with known protein and membrane stabilizing capability. Because of these unique chemical properties, this molecule has been the focuses of study in several neurodegenerative diseases, which are associated with the misfolding of disease-specific proteins. These conditions, include Alzheimers Disease (AD) an amyloid proteinopathy, Huntington’s Disease (HD), an expanded polyglutamine proteinopathy and oculopharyngeal muscular dystrophy (OPMD), an expanded polyalanine proteinopathy.
In each disease, specific misfolded aggregate-prone proteins are resistant to the normal cellular processes of protein turnover and accumulate in insoluble inclusions in regions specific to each disease. While insoluble aggregates correlate with disease progression, there is increasing evidence that the initiating and most toxic events are caused by soluble protein oligimers or microaggregates. Trehalose is thought to work by interfering with production or enhancing destruction of toxic fragments.
One of the fascinating aspects of trehalose is its presence in various organisms that can survive at the extremes of temperature and dehydration. This observation led to work which showed that trehalose is a naturally occurring reducer of cell stress, protecting these organisms from extremes in heat shock and osmotic stress (Crowe, 2002). Trehalose is thought to act by altering or replacing the water shell that surrounds lipid and protein macromolecules (Colaco et al., 1992). It is thought that its flexible glycosidic bond allows trehalose to conform to the irregular polar groups of macromolecules. In so doing, it is able to maintain the three-dimensional structure of these biologic molecules under stress, preserving biologic function.
As an extension of its natural capability to protect biological structures, trehalose has been used for the preservation and protection of biologic materials. It stabilizes bioactive soluble proteins such as monoclonal antibodies and enzymes for medical use (Colaco et al., 1992). It stabilizes proteins for inhaled use (Strickley and Anderson 1997). It is used to preserve cellular blood products for transfusion and greatly extends the shelf life of platelets (Crowe et al., 2003) and cord blood (Zhang et al., 2003). It is used to preserve embryos during freeze-drying where it increases viability (Suzuki et al., 1996). It is used in cryopreservation of transplant cells and tissue where it has been shown to increase viability (Beattie et al., 1997) and decrease host immune response (Erdag et al., 2002). Building on extensive study in multiple biologic systems that describe its ability to inhibit lipid and protein misfolding (Singer and Lindquist, 1998), trehalose has become an attractive molecule for study in neurodegenerative disease characterized by protein misfolding and aggregate pathology. Such diseases include Alzheimer's and Parkinson's disease and the less common triplet repeat diseases. Recent scientific publications describe trehalose benefit in model systems that recapitulate aggregate pathology that characterize Alzheimer's (AD), Huntington's (HD) and occulopharyngeal muscular dystrophy (OPMD). The mechanisms by which trehalose metabolism alters plant development are largely unknown. Trehalose itself could affect development by acting as signal molecule in carbohydrate metabolism. For example, trehalose induces enzymes of fructan synthesis in barley (Wagner et al., 1986; Mulleret al., 2000) and Sucrose synthase activity in soybean (Muller et al., 1998). In general, sugars such as Sucrose and Glucose act as signals in the regulation of gene expression (Koch, 1996). Whereas, the expression of several source-specific genes is probably regulated by hexoses in a hexokinase-dependent signaling pathway (Jang and Sheen, 1994; Jang et al., 1997; Dai et al., 1999), the regulation of the expression of some other genes appears to be directly mediated by Sucrose without prior cleavage to hexoses (Yokoyama et al., 1994; Chiou and Bush, 1998; Rook et al., 1998). It is conceivable that trehalose, which is structurally similar to Sucrose, might act as an analog of Sucrose in sugar sensing. In addition to the trehalose that may be produced by the plants themselves, plants are also exposed to trehalose formed by microorganisms in mutualistic, as well as in pathogenic interactions. Trehalose formed by rhizobia during nodulation appears to have a strong effect on the carbohydrate contents in root nodules (Muller et al., 1995b). It is possible that trehalose-producing plant symbionts and/or pathogens can exploit the effects of trehalose to gain control over the sugar-sensing system of the plant. If this is the case, the trehalose-degrading enzyme trehalase, which is widespread among higher plants and is found in multiple tissues, may provide a safeguard against potentially deleterious effects of trehalose on carbohydrate allocation in plant-microbe interactions (Muller et al., 1995a; Aeschbacher et al., 1999).
MATERIALS AND METHODS
In May 2009 the 100 seeds were surface sterilized by immersion in sodium hypochloride (40% (v/v)) for 20 min, rinsed with distilled water and transferred into plastic pots (8 cm 42 diameter) filled with perlite. The Seeds were planted in to sterile soil under conditions at 20°C with 4- 6 weeks light. Then extraction by coffee machine differents parts of plant: such as flowers, roots, shoots and leaves. The insoluble pellets remaining from the carbohydrate extraction were resuspended in 0.2 mL of NaOH (0.5 m) and incubated at 60°C for 1 h. HCl (0.2 mL, 0.5 m) was subsequently added. After cooling down to room temperature, 0.6 mL of acetate/Na+ buffer (0.2 m, pH 4.5) containing 1 unit of amyloglucosidase (special quality for starch determination, Boehringer Mannheim, Germany) was added and the samples was incubated overnight at 37°C. The reaction was stopped by boiling for 2 min. The samples were centrifuged (10 min at 10,000 g); the supernatants were 10 times diluted and were analyzed for Glc formation using HPLC.
RESULTS AND DISCUSSION
Trehalose benefit was first shown in Huntington's model systems (Tanka et al., 2004). Huntington's is an autosomal dominant neurodegenerative disease, which presents with cognitive impairment, involuntary choreiform movements and psychiatric manifestations. Onset is generally in midlife, but can occur in childhood and old age. It inexorably progresses to disability and death over a 10-25 year period. Huntington's is characterized by an expanded CAG repeat within the first exon of the Huntington gene. The mutant protein generated has an expanded polyglutamine (polyGN) tract. The pathologic hallmark of this and other polyGN diseases is the formation of aggregates, containing misfolded mutant protein in both cytoplasm and nucleus of affected cells.
Although, trehalose absorption in humans has not been well studied, a small fraction (0.5%) is likely to be absorbed by passive diffusion, as has been demonstrated for other dissacharides (Van Elburg et al., 1995). Tanaka et al. (2004) demonstrated that trehalose inhibits polyglutamine-mediated protein aggregation of a model polyGN protein in vitro solution and that it decreases aggregate formation and prolongs viability in a model cell culture. This same group went on to show that trehalose ameliorated motor symptoms, decreased aggregate number and size and prolonged life by 20% in the R6/2 transgenic mouse model of Huntington's.
In this study, Arabidopsis thaliana L. in the stress conditions, growthing was analysed different kinds of Arabidopsis for the presence of trehalose.
In Arabidopsis, inhibition of trehalase causes the accumulation of trehalose and a strong reduction in starch and sucarus contents, suggesting a role for trehalose and trehalase in carbon allocation (Muller et al., 2001). In addition, trehalose has been show to inhibit Arabidopsis seedling root elongation and cause starch accumulation in shoots. Furthermore, trehalose increases AGPase (ADP-Glc pyrophosphorylase) activity and induces APL3 gene expression (Wingler et al., 2000; Fritzius et al., 2001). In soybean, trehalose also affects Suc synthase and invertase activities (Muller et al., 1998). How trehalose affects plant gene expression, enzyme activities, photosynthetic activity and carbon allocation is not clear, but trehalose-6-phosphate does not appear to have any effect on plant hexose phosphorylation (Wiese et al., 1999). However, transgenic tobacco plants expressing Escherichia coli homologs of TPS and trehalose-6-phosphate phosphatase show a positive correlation between trehalose-6-phosphate levels and photosynthetic activity, suggesting a regulatory role for trehalose-6-phosphate in plant carbohydrate metabolism (Paul et al., 2001). The distribution and activity of trehalase was measured in mature Arabidopsis grown. In these plants, a strong trehalase activity was found in mature flowers and roots, whereas leaves, stems, had significantly lower activities (Fig. 1).
Accordingly, its structure/activity benefits would be expected to persist for relatively extended times. If planned studies demonstrate brain or cerebral spinal fluid absorption, trehalose will open a new avenue of potential therapy for the prevention and treatment of multiple neurodegenerative diseases. This review summarized evidence for protective benefit in models of Huntingtin's disease, oculopharyngeal muscular dystrophy and Alzheimer's.
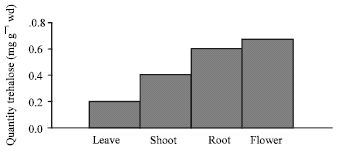 | |
| Fig. 1: | Using as anion-exchange High Performance Liquid Chromatography (HPLC) analysis, trehaloses was highest in flower and root a balance of changes quantity trehalose in Arabidobsis thaliana L. that is most quantity 0. 63 mg-1 wd in flowers and least quantity 0.2 mg-1 wd in leave |
Although not studied, Parkinson's disease and amyotrophic lateral sclerosis display aggregate pathology that may be amenable to similar response.
ACKNOWLEDGMENTS
The author is grateful to Dr. Shahab Shariati, Dr. Akbar Islam Nezhad, Mr. Assghar Islam Nezhad, Dr. Alireza Eslami and grateful to support Department of Plant Biology, Islamic Azad University, Rasht Branch, Guilan, Iran and Young Researchers Club Rasht Branch.
REFERENCES
- Aeschbacher, R., J. Muller, T. Boller and A. Wiemken, 1999. Purification of the trehalase GmTRE1 from soybean nodules and cloning of its cDNA: GmTRE1 is expressed at a low level in multiple tissues. Plant Physiol., 119: 489-496.
Direct Link - Beattie, G.M., J.H. Crowe, A.D. Lopez, V. Cirulli, C. Ricordi and A. Hayek, 1997. Trehalose: A cryoprotectant that enhances recovery and preserves function of human pancreatic islets after long-term storage. Diabetes, 46: 519-523.
Direct Link - Chiou, T.J. and D.R. Bush, 1998. Sucrose ifs a signal molecule in assimilate partitioning. Proc. Natl. Acad. Sci. USA., 95: 4784-4788.
Direct Link - Colaco, C., J. Kampinga and B. Roser, 1995. Amorphous stability and trehalose. Scence, 268: 788-788.
Direct Link - Colaco, C., S. Sen, M. Thangavelu, S. Pinder and B. Roser, 1992. Extraordinary stability of enzymes dried in trehalose: Simplified molecular biology. Biotechnology, 10: 1007-1011.
CrossRefDirect Link - Crowe, J.H., F. Tablin, W.F. Wolkers, K. Gousset, N.M. Tsvetkova and J. Ricker, 2003. Stabilization of membranes in human platelets freeze-dried with trehalose. Chem. Phys. Lipids, 122: 41-52.
CrossRef - Crowe, L.M., 2002. Lessons from nature: The role of sugars in anhydrobiosis. Comp. Biochem. Physiol. A Mol. Integrative Physiol., 131: 505-513.
CrossRef - Dai, N., A. Schaffer, M. Petreikov, Y. Shahak and Y. Giller et al., 1999. Overexpression of arabidopsis hexokinase in tomato plants inhibits growth, reduces photosynthesis and induces rapid senescence. Plant Cell, 11: 1253-1266.
Direct Link - Erdag, G., A. Eroglu, J. Morgan and M. Toner, 2002. Cryopreservation of fetal skin is improved by extracellular trehalose. Cryobiology, 44: 218-228.
PubMedDirect Link - Fritzius, T., R. Aeschbacher, A. Wiemken and A. Wingler, 2001. Induction of ApL3 expression by trehalose complements the starch-deficient Arabidopsis mutantadg2-1 lacking ApL1, the large subunit of ADP-glucose pyrophosphorylase. Plant Physiol., 126: 883-889.
CrossRefDirect Link - Oliver, A.E., K. Jamil, J.H. Crowe and F. Tablin, 2004. Loading human mesenchymal stem cells with trehalose by fluid-phase endocytosis. Cell Preserv. Technol., 2: 35-49.
CrossRefDirect Link - Paul, M., T. Pellny and O. Goddijn, 2001. Enhancing photosynthesis with sugar signals. Trends Plant Sci., 6: 197-200.
CrossRefDirect Link - Jang, J.C., P. Leon, L. Zhou and J. Sheen, 1997. Hexokinase as a sugar sensor in higher plants. Plant Cell, 9: 5-19.
Direct Link - Koch, K.E., 1996. Carbohydrate-modulated gene expression in plants. Ann. Rev. Plant Physiol. Plant Mol. Biol., 47: 509-540.
PubMedDirect Link - Muller, J., R.A. Aeschbacher, A. Wingler, T. Boller and A. Wiemken, 2001. Trehalose and trehalase in Arabidopsis. Plant Physiol., 125: 1086-1093.
Direct Link - Muller, J., R. Aeschbacher, N. Sprenger, T. Boller and A. Wiemken, 2000. Disaccharide-mediated regulation of sucrose: Fructan-6-fructosyltransferase (6-SFT), a key enzyme of fructan synthesis in barley leaves. Plant Physiol., 123: 265-274.
Direct Link - Muller, J., T. Boller and A. Wiemken, 1995. Effects of validamycin A, a potent trehalase inhibitor and phytohormones on trehalose metabolism in roots and nodules of soybean and cowpea. Planta, 197: 362-368.
CrossRefDirect Link - Muller, J., T. Boller and A. Wiemken, 1998. Trehalose affects sucrose synthase and invertase activities in soybean (Glycine max L. Merr.) roots. J. Plant Physiol., 153: 255-257.
Direct Link - Rook, F., N. Gerrits, A. Kortstee, M. van Kampen, M. Borrias, P. Weisbeek and S. Smeekens, 1998. Sucrose-specific signalling represses translation of the Arabidopsis ATB2 bZIP transcription factor gene. Plant J., 15: 253-263.
Direct Link - Singer, M.A. and S. Lindquist, 1998. Multiple effects of trehalose on protein folding in vitro and in vivo. Mol. Cell, 1: 639-648.
Direct Link - Strickley, R.G. and B.D. Anderson, 1997. Solid-state stability of human insulin. II. Effect of water on reactive intermediate partitioning in lyophiles from pH 2-5 solutions: Stabilization against covalent dimer formation. J. Pharm. Sci., 86: 645-653.
Direct Link - Suzuki, T., A. Boediono, M. Takagi, S. Saha and C. Sumantri, 1996. Fertilization and development of frozen-thawed germinal vesicle bovine oocytes by a one-step dilution method in vitro. Cryobiology, 33: 515-524.
CrossRefDirect Link - Tanaka, M., Y. Machida, S. Niu, T. Ikeda and N.R. Jana et al., 2004. Trehalose alleviates polyglutamine-mediated pathology in a mouse model of Huntington disease. Nat. Med., 10: 148-154.
CrossRefDirect Link - Van Elburg, R.M., J.J. Uil, F.T. Kokke, A.M. Mulder, W.G. van de Broek, C.J. Mulder and H.S. Heymans, 1995. Repeatability of the sugar-absorption test, using lactulose and mannitol, for measuring intestinal permeability for sugars. J. Pediatr. Gastroenterol. Nutr., 20: 184-188.
Direct Link - Wagner, W., A. Wiemken and P. Matile, 1986. Regulation of fructan metabolism in leaves of barley (Hordeum vulgare L. cv Gerbel). Plant Physiol., 81: 444-447.
Direct Link - Wiese, A., F. Groner, U. Sonnewald, H. Deppner and J. Lerchl et al., 1999. Spinach hexokinase I is located in the outer envelope membrane of plastids. FEBS Lett., 461: 13-18.
Direct Link - Wingler, A., T. Fritzius, A. Wiemken, T. Boller and R.A. Aeschbacher, 2000. Trehalose induces the ADP-glucose pyrophosphorylase gene, APL3 and starch synthesis in Arabidopsis. Plant Physiol., 124: 105-114.
CrossRefDirect Link - Yokoyama, R., T. Hirose, N. Fujii, E.T. Aspuria, A. Kato and H. Uchimyia, 1994. The rolC promoter of Agrobacterium rhizogenes Ri plasmid is activated by sucrose in transgenic tobacco plants. Mol. General Genet., 244: 15-22.
Direct Link - Zhang, X.B., K. Li, K.H. Yau, K.S. Tsang and T.F. Fok et al., 2003. Trehalose ameliorates the cryopreservation of cord blood in a preclinical system and increases the recovery of CFUs, long-term culture-initiating cells and nonobese diabetic-SCID repopulating cells. Transfusion, 43: 265-272.
Direct Link