
Shahidul Islam
Department of Agriculture, University of Arkansas at Pine Bluff, 1200 North University Drive, Mail Slot 4913, Pine Bluff, AR 71601, USA
R. Carvajal R. Carmen
Mississippi State University, Mississippi, USA
Jr. James O. Garner
Department of Agriculture, University of Arkansas at Pine Bluff, 1200 North University Drive, Mail Slot 4913, Pine Bluff, AR 71601, USA
American Journal of Plant Physiology
Year: 2009 | Volume: 4 | Issue: 1 | Page No.: 9-17







ABSTRACT
Twenty five cultivars were screened for germination at low (10°C), moderate (30°C) and at stress (40°C) temperatures. Three cultivars were chosen such as Texas Cream 40 was able to germinate at very high and low temperatures. Black Crowder demonstrated acceptable germination at high temperatures but negatively affected at low temperature. The main sugars present in cowpea seed were sucrose, raffinose and stachyose. Sugar contents were affected by cultivar, type of tissue and temperature. Sucrose contents were higher in embryo tissue of cultivars with low percent germination and reduced in the cultivar with higher percent germination suggesting the use of sucrose for germination. Sucrose decreased greatly at 30°C and increased again at 40°C. Raffinose and stachyose contents were higher in ungerminated seed. In germinated seed, raffinose and stachyose contents were found only in cotyledon tissues at 10°C. Peroxidase activities was affected by cultivars, type of tissue and temperature. The highest peroxidase activity was found at low temperature (10°C) in embryo tissue of the cultivar with the highest germination. High peroxidase activity was related to ability of seed to germinate at low temperature.
PDF Abstract XML References Citation
How to cite this article
Shahidul Islam, R. Carvajal R. Carmen and Jr. James O. Garner, 2009. Carbohydrate Compositions and Peroxidase Activity in Ungerminated,
Cotyledon and Embryo Tissues of Vigna unguiculata L. Walp Seed
Grown Under Stress Temperatures. American Journal of Plant Physiology, 4: 9-17.
DOI: 10.3923/ajpp.2009.9.17
URL: https://scialert.net/abstract/?doi=ajpp.2009.9.17
DOI: 10.3923/ajpp.2009.9.17
URL: https://scialert.net/abstract/?doi=ajpp.2009.9.17
INTRODUCTION
Cowpea (Vigna unguiculata L. Walp) is an annual, herbaceous legume, indigenous to Africa (Craufurd et al., 1996; Hall et al., 1997). It is a high protein (25%), fiber (6.3%) and low fat (1.9%) vegetable crop having good nutritional qualities (Ricardo, 1985; Fery, 1990, 2002). It is a short-day crop sensitive to chilling temperatures (Ehlers and Hall, 1997; Hall et al., 1997) and is adapted to warm weather and humid conditions (Craufurd et al., 1996; Hall et al., 1997). It is more tolerant to drought, water logging, infertile soils and acid stress, than common beans.
Cowpeas have been reported to have greater adaptation to high temperatures than any other crop species. Germination greater than or equal to 80% has been observed in cowpea at temperatures ranging from 10 to 40°C (Wolk and Herner, 1982; Ismail et al., 1997, 1999). Temperatures outside this range adversely affect germination of cowpea. Variation in germination percentage and rate of germination under stress temperatures have been observed among cowpea cultivars (Marsh, 1993; Craufurd et al., 1996).
Peroxidases (donor: H2O2 oxidoreductase; EC 1.11.1.7), a ubiquitous class of plant proteins, are enzymes whose primary function is to oxidize a variety of hydrogen donors at the expense of hydrogen peroxide (Dunford and Stillman, 1976; Gaspar et al., 1982). It has been implicated in a variety of physiological process in plants including seed vigor and viability. Seed vigor has also been associated with carbohydrate content. In corn (Zea mays L.), the decline of sucrose and raffinose contents during aging was closely correlated with the loss of ability to germinate and rate of germination (Bernal-Lugo and Leopold, 1992). As a chilling-sensitive crop, cowpea is affected during germination and early seedling development if planted late in the spring when temperatures are still low in subtropical areas. Good stand establishment is necessary to avoid yield loss, thus it would be beneficial to plant a cultivar able to tolerate low temperatures. The objectives of this research were to determine the carbohydrate composition of the cowpea seed prior to and during germination and to evaluate the peroxidase enzyme activity in cowpea seed and its effect on chilling tolerance.
MATERIALS AND METHODS
Cultivar/Genotype
Trials were carried out over two years using 25 cowpea cultivars/genotypes grown to initially screen for germination and coefficient of variability of germination tests at 10, 30 and 40°C (Islam et al., 2005, 2006). Three cultivars were chosen from 25 for further study for biochemical characteristics such as Texas Cream 40 which showed ability to germinate at very high (40°C) and low (10°C) temperature; Black Crowder which had acceptable high germination at 40°C, but reduced germination at 10°C and Mississippi Purple which exhibited lower germination at all temperatures tested 40OC (Islam et al., 2005, 2006). The seeds were obtained from Native Seed Search, Tucson, AZ, USA. The experiment was conducted during 2000 to 2002 at the Mississippi State University, USA. To get seeds from the same lot, cultivars were grown in the same year, under a common set of environmental conditions, handled using the same harvesting/seed processing equipment and stored under the same conditions.
Environmental Conditions
The germination tests were conducted in germination chambers each of which was set at a different temperature: 10°C (low temperature), 30°C (moderate temperature) and 40°C (high temperature) (Islam et al., 2005, 2006). The control treatment having a constant temperature of 24°C. All chambers were maintained at 100%±1 relative humidity. Light/dark periods were maintained at 8/16 h day-1 at all three temperatures.
Extraction and Analysis of Carbohydrates
Seed carbohydrates were extracted and identified following the methodology by Connor and Sowa (2000). To determine the initial content of carbohydrates in cowpea seed, whole dry seed were ground using a handy mill machine and 0.5 g of the resulting flour was used for carbohydrate extraction. For extraction from the germinating seeds, the seeds were dissected into cotyledons and axes after radical protrusion and before first leaf expansion. Tissues from each cultivar germinated at each temperature were separately dried in a freeze-dry system (Labconco Freezone 4.5 LABCONCO Corporation, Kansas City, MO). A 0.2 to 0.5 g dry tissue sample was used for each carbohydrate extraction. The tissue samples were placed in test tubes with 10 mL of an 80% ethanol solution and then boiled in a 75°C water bath for 1 h then filtered. Additional ethanol solution was used to rinse the samples. The extracts were poured into evaporation flasks and then roto-evaporated to dryness in a flask-evaporator (Buchler Instruments, Fort Lee, NJ). The evaporation flasks were rinsed with 10 mL of distilled water, and the samples were freeze-dried overnights in a stoppered vacuum flask attached to the freeze-dry apparatus. The samples were dissolved in 1 mL of Trimethylsilylimidiazole (TMSI), then heated in a 75°C water bath for 30 min and aspirated to dryness. The samples were then resuspended in 2 mL of chloroform and stored refrigerated until analysis. The analysis of carbohydrates was performed in a Hewlett Packard (HP) Gas Chromatograph (GC) equipped with a flame ionization detector and using a 125-5037 DB-5 column (30 m length x 0.53 mm I.D. and 0.5 μm film thickness) (J and W Scientific Folsom, CA). The program in the GC preset a detector and injector temperature of 230°C. An initial oven temperature of 210°C was held for 7 min and then increased 15°C min-1 until a temperature of 270°C was reached and held for 25 min. The flow was kept at 1.76 nn mL-1. Data were collected and printed in a HP printer attached to the GC. A graph and numerical map with the respective percentage peak area of each sugar was obtained. The carbohydrates were identified by comparing with standards of pure sugars prepared in a similar manner to the tissue samples and injected in the GC.
Peroxidase Extraction
Peroxidase was extracted using a slight modification of the methodology of Silva et al. (1990). Seed were separated into cotyledons and axes and 3.0 g of each tissue were used for extraction. Each sample was ground using a pre-chilled mortar and pestle and 3.0 mL of citrate phosphate buffer (pH 6.0) was added as a grinding solution. This material was then placed in a centrifuge tube. An additional 3.0 mL of the buffer was used to wash the mortar and pestle and this plus a small amount of Polyvinyl-polypyrrolidone (PVP) was added to the tube. The samples were centrifuged at 6000 x g at 5°C for 10 min and the supernatant was saved and maintained at freezing temperature for the next step of enzyme analysis.
Peroxidase Activity
Peroxidase activity was measured using the methodology of Silva et al. (1990). The enzymatic activity of peroxidase was measured spectrophotometreically in a Spectronic Genesys 5 spectrophotometer (Spectronic Instruments Inc., Rochester, NY). One milliliter of crude peroxidase extract was added to a reaction mixture consisting of 2 mL of 0.05 M citrate phosphate buffer (pH 6.0) and 2 mL of 4.0 x 10-4 guaiacol. The reaction was initiated by adding 2 mL of 3% H2O2. The change of the absorbance at 470 nm was used to measure the enzyme activity.
Statistics
The data were analyzed as a combined series of CRDs. Data were subjected to Analysis of Variance (ANOVA) using the General Linear Models (GLM) procedure of SAS version 8.1 (SAS Instit., Inc. Cary, North Carolina, USA). Mean separations were done using Fisher′s protected Least Significant Difference (LSD) test.
RESULTS AND DISCUSSION
Carbohydrate compositions were determined in whole ungerminated seed and in cotyledon and embryo tissues of germinating seed at low (10°C), moderate (30°C) and high (40°C) temperature. The main sugars present in cowpea seed were sucrose, raffinose and stachyose. The sugar content was affected by temperature, cultivar and types of tissue.
Sucrose Content
There was a significant temperature x cultivar x tissue interaction effect for sucrose content of cowpea seed (Table 1). The initial sucrose content in ungerminated seed was highest in BC, followed by TC-40 and lowest in MP. At low temperature (10°C), the higher mean sucrose content was in the embryo tissue of MP followed by BC and the lowest sucrose content was in TC-40. In cotyledon tissue, the highest mean sucrose content was in TC-40 and the lowest in MP and BC (Table 1). At moderate temperature (30°C), the highest sucrose content was in cotyledon of TC-40 (53.51 mg g-1) and the lowest was in MP. In embryo tissues, highest mean sucrose content was in TC-40 and the lowest in MP and BC. At high temperature (40°C), the highest mean sucrose content in cotyledon tissues was in TC-40 and MP, followed by BC. In embryo tissue there were no significant differences among cultivars for mean sucrose content. Sucrose is primary nutrient in cells of higher plants (Xu et al., 1989). During seed development changes in carbohydrates occur in such way that monosaccharide content decreases whereas oligosaccharides content increases, but during the germination process this pattern is reversed (Vertucci and Farrant, 1995). At low temperature TC-40 seed had faster and higher percent germination than MP and BC (Islam et al., 2006); thus, the higher sucrose content in the cotyledon and lower sucrose contents in embryos of the above cultivar suggest faster and more efficient use of sucrose in germination process and seedling development in this cultivar. Xu et al. (1989) indicated that during germination seeds have a dual role: first as a source of sucrose produced in cotyledons and secondly as sink when the young seedling uses sucrose for development.
| Table 1: | Sucrose content (mg g-1) of whole seed, cotyledon (Cot.) and embryo (Emb.) tissues of seed of three cultivars of cowpea germinated at three different temperatures |
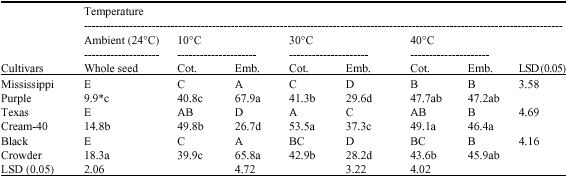 | |
| * Data are means of five replications of 20 seed each foe each cultivar at each temperature. a-d: Means within a temperature not followed by a common lowercase letter are significantly (p<0.05) different according to Fisher′s protected LSD test.A-E: Means within a cultivar not followed by a common capital letter are significantly (p<0.05) different according to Fisher′s protected LSD test | |
| Table 2: | Raffinose content (mg g-1) of whole seed, cotyledon (Cot.) and embryo (Emb.) tissues of seed of three cultivars of cowpea germinated at three different temperatures |
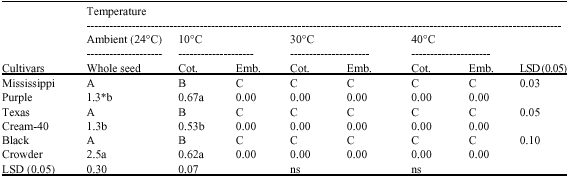 | |
| * Data are means of five replications of 20 seed each foe each cultivar at each temperature. a-b: Means within a temperature not followed by a common lowercase letter are significantly (p<0.05) different according to Fisher′s protected LSD test.A-C: Means within a cultivar not followed by a common capital letter are significantly (p<0.05) different according to Fisher′s protected LSD test | |
Raffinose Content
There was a significant temperature x cultivar interaction for raffinose content (Table 2). In whole ungerminated seed, the highest raffinose content mean was in BC (2.5 mg g-1) and lowest in TC-40 (1.5 mg g-1) and MP (1.3 mg g-1). In germinating seed at low temperature (10°C), raffinose contents in cotyledon tissues were 0.67 mg g-1 (MP), 0.62 mg g-1 (BC) and 0.35 mg g-1 (TC-40). No raffinose was detected in embryo tissues at low temperature or in any tissue at 30 and 40°C.
| Table 3: | Stachyose content (mg g-1) of whole seed, cotyledon (Cot.) and embryo (Emb.) tissues of seed of three cultivars of cowpea germinated at three temperatures |
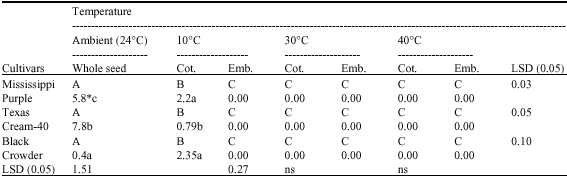 | |
| * Data are means of five replications of 20 seed each foe each cultivar at each temperature. a-b: Means within a temperature not followed by a common lowercase letter are significantly (p<0.05) different according to Fisher′s protected LSD test.A-C: Means within a cultivar not followed by a common capital letter are significantly (p<0.05) different according to Fisher′s protected LSD test | |
Stachyose Content
There was a significant temperature x cultivar interaction for stachyose content (Table 3). The highest stachyose content mean in the whole ungerminated seed was in BC (10.5 mg g-1), followed by TC-40 (7.8 mg g-1) and MP (5.8 mg g-1). Stachyose was also detected in cotyledon tissues of germinating seed at low temperature (10°C) in BC (2.4 mg g-1), MP (2.2 mg g-1) and TC-40 (0.8 mg g-1). No stachyose was detected in tissues of germinating seeds at moderate or high temperatures, which suggest that larger chain sugars had been reduced to sucrose or other monosaccharides. Koster and Leopold (1988) demonstrated that raffinose as well as stachyose have an important role helping in dessication tolerance of pea, corn and soybean seed, probably helping to prevent sucrose crystallization. The increase in sugar content when comparing the dry ungerminated seed to the sugar content in cotyledon tissue of germinating seed at low temperature indicated either de novo synthesis of or convection of other products to sucrose. Miquel and Browse (1995) pointed out that seed storage components such as lipids, proteins and carbohydrates are converted to sucrose in order to feed the developing embryo.
Peroxidase Activity
At low temperature (10°C) which is considered a chilling temperature for cowpea cultivars, the highest peroxidase activity was in embryo tissue of TC-40 (3 ΔA m-1 g-1), followed by embryo tissue of BC (2.7 ΔA m-1 g-1) (Fig. 1). The lowest peroxidase activity was in cotyledon tissues of TC-40 (1.4 ΔA m-1 g-1) and MP (1.9 ΔA m-1 g-1) (Fig. 2). At moderate temperature (30°C) the higher peroxidase activity was in embryo tissues of BC (2.9 ΔA m-1 g-1) and TC-40 (2.5 ΔA m-1 g-1) and the lowest in cotyledon tissue of MP (0.8 ΔA m-1 g-1). As at low temperature, higher peroxidase activity was observed in embryo tissue than cotyledon tissue. At high temperature (40°C) the highest peroxidase activity was in cotyledon tissue of BC (2.2 ΔA m-1 g-1), followed by embryo tissues of TC-40 (1.9 ΔA m-1 g-1) and MP (1.8 ΔA m-1 g-1). The lower peroxidase activity was in cotyledon tissues of MP (1.38 ΔA m-1 g-1) and TC-40 (1.4 ΔA m-1 g-1) and in embryo tissues of BC (1.4 ΔA m-1 g-1). There were a significant (p<0.01) positive correlation between sucrose contents at different temperatures and peroxidase activity of cowpea seeds (Fig. 3). In the case of cotyledon tissues, sucrose contents at 40°C has the most intimate relation (r = 89) with peroxidase activity of cowpea seeds followed by the treatment 30°C (r = 0.88). Like cotyledon, the sucrose contents of the embryo tissues were highly correlated (r = 0.85) with peroxidase activity of cowpea seeds at 40°C followed by the treatment 30°C (r = 0.49). The results indicate that sucrose de novo synthesis was higher in cowpea seeds and it played a vital role in the biochemical process under stress temperatures. Peroxidase activity in plant tissue increase as a response to stress treatment (Wagih and Coutts, 1982). Marafon et al. (2008) reported that in Prunus persica L. Batsch, the high peroxidase activity during dormancy may be attributed to cold tolerance due to oxidative stress reduction caused by low temperatures. The enzyme activity of cowpea leaves was increased by mechanical abrasion or as a response to be infection by Tobacco Necrosis Virus. Chilling temperatures is thought to increase the levels of active oxygen species, which could increase the risk of chilling injury, thus increase in the activity of the active oxygen species scavenging system is desirable to increase tolerance to chilling temperature. The peroxidase activity frequently increased in response to stress and one of the main functions of this enzyme in the cellular protection against oxidative reactions observed in several tissues of the plants (Siegel, 1993). The cowpea cultivar with the higher germination percentage, at low temperature, had higher peroxidase activity, suggesting the peroxidase acted in scavenging free radicals, which if accumulated could cause damage to cellular component and alters metabolic function (Siegel, 1993). This is in agreement with Arderson et al. (1995) who mentioned that peroxidase activity increased in chilling acclimated maize seedling.
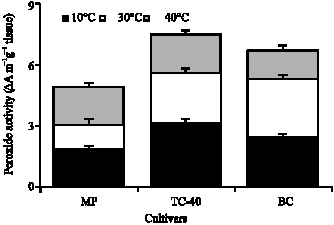 | |
| Fig. 1: | Peroxide activity (ΔA m-1 g-1 tissue) embryo (Emb.) tissues of seed of three cultivars of cowpea germinated at three temperatures. Data are means of five replications. Bars indicate ±SE |
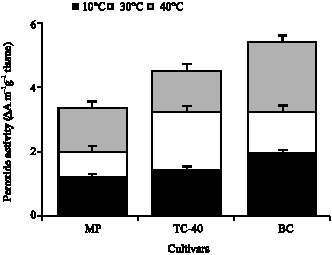 | |
| Fig. 2: | Peroxide activity (ΔA m-1-g1 tissue) cotyledon (Cot.) tissues of seed of three cultivars of cowpea germinated at three temperatures. Data are means of five replications. Bars indicate ±SE |
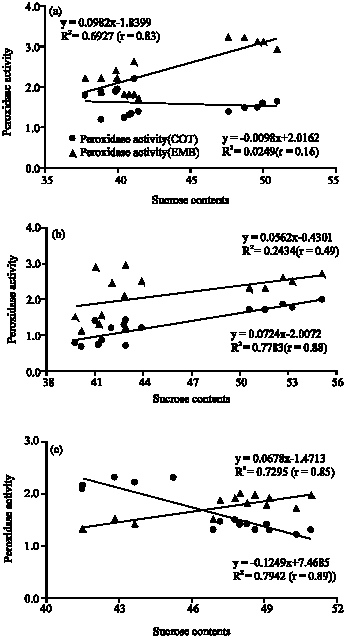 | |
| Fig. 3: | Correlation between peroxidase activity and sucrose contents at (a) 10°C, (b) 30°C and (c) 40°C of cotyledon (COT) and embryo (EMB) tissues of seed cowpea seeds (n = 15) |
CONCLUSION
In conclusion, it is apparent that the main sugars present in cowpea seed were sucrose, raffinose and stachyose. Raffinose and stachyose are stored carbohydrate present only in dry ungerminated seed and in cotyledon tissues of germinating seed at low temperature. No de novo synthesis of these sugars was detected. Sucrose de novo synthesis was found at higher temperatures. An accumulation of this sugar was evident in embryo tissues of cultivars with reduced ability to germinate at low temperature. The results also indicated that high peroxidase activity was related to ability of seed to germinate at low temperature. The cowpea cultivars with high germination percentage showed higher sucrose, raffinose and stachyose, as well as peroxidase activity in ungerminated seed. The results provided by this research will facilitate future plant physiological and genetic studies of cowpea as well as other plant species.
REFERENCES
- Anderson, M.D., T.K. Prasad and C.R. Stewart, 1995. Changes in isozyme profiles of catalase, peroxidase and glutathione reductase during acclimation to chilling in mesocotyls of maize seedlings. Plant Physiol., 109: 1247-1257.
CrossRefPubMedDirect Link - Bernal-Lugo, I. and A.C. Leopold, 1992. Changes in soluble carbohydrate during seed storage. Plant Physiol., 98: 1207-1210.
PubMedDirect Link - Craufurd, Q., H. Ellis, J. Summerfield and L. Menin, 1996. Development in cowpea (Vigna unguiculata). I. The influence of temperature on seed germination and seedling emergence. Exp. Agric., 32: 1-12.
CrossRefDirect Link - Connor, K.F. and S. Sowa, 2000. Sucrose accumulation as a response to desiccation in Quercus alba L. Acorn: A natural glycoprotectant. Proceedings of the Ecological Society of America 85th Annual Meeting, August 6-10, 2000, Snowbird, Utah, pp: 235-235.
Direct Link - Ehlers, J.D. and A.E. Hall, 1997. Cowpea (Vigna unguiculata L. Walp.). Field Crops Res., 53: 187-204.
CrossRefDirect Link - Fery, L., 1990. The cowpea: Production, utilization and research in the United States. Horticul. Rev., 12: 197-222.
CrossRefDirect Link - Islam, S., C.R. Carmen and J.O. Garner, 2006. Screening for tolerance of stress temperature during germination of twenty five cowpea (Vigna unguiculata L. Walp) cultivars. J. Food Agric. Environ., 4: 191-195.
Direct Link - Ismail, A., A. Hall and T. Close, 1997. Chilling tolerance during emergence of cowpea associated with a dehydrin and slow electrolyte leakage. Crop Sci., 37: 1270-1277.
Direct Link - Ismail, A., A. Hall and T. Close, 1999. Allelic variation of a dehydrin gene cosegregates with chilling tolerance during seedling emergence. Proc. Natl. Acad. Sci. USA., 96: 13566-13570.
PubMedDirect Link - Koster, K.L. and A.C. Leopold, 1988. Sugar and dessication tolerance in seeds. Plant Physiol., 88: 829-832.
PubMedDirect Link - Marsh, L., 1993. Moisture affects cowpea and okra seed emergence and growth at low temperatures. HortSci., 28: 774-777.
Direct Link - Wagih, E.E. and R.H.A. Coutts, 1982. Peroxidase, poliphenoloxidase and ribonuclease in tobacco necrosis virus infected or mannitol osmotically-stressed cowpea and cucumber tissue. J. Phytopathol., 104: 124-137.
CrossRefDirect Link - Xu, D.P., S. Shi-Jean and C.C. Black, 1989. Sucrose metabolism in Lima been seeds. Plant Physiol., 89: 1106-1116.
PubMedDirect Link - Marafon, A.C., M.A. Bacarin, F.G. Herter and A.C. Rodrigus, 2008. Carbohydrate level, peroxidase activity and vascular connection in tissues of peach trees (Prunus persica L. Batsch) cv. Jubileu with peach tree short life symptoms during the dormancy period. Acta Hortic., 774: 397-402.
Direct Link
anuj k. ahlavat Reply
the paper is good enough to increase the knowledge about biochemical status of cotyledons and embryo during germination.