
Z. Zainal
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600, Bangi, Selangor Darul Ehsan, Malaysia
E. Marouf
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600, Bangi, Selangor Darul Ehsan, Malaysia
I. Ismail
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600, Bangi, Selangor Darul Ehsan, Malaysia
C.K. Fei
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600, Bangi, Selangor Darul Ehsan, Malaysia
American Journal of Plant Physiology
Year: 2009 | Volume: 4 | Issue: 2 | Page No.: 70-79







ABSTRACT
Plant defensins are a group of pathogenesis-related defense mechanism proteins. Transgenic tomato plants expressing the chili defensin gene under the control of a Cauliflower Mosaic Virus 35S (CaMV 35S) promoter were generated. The 5 kDa peptide, corresponding to the chili defensin protein was detected in the total protein fraction extracted from the transgenic plants. When compared to partially-purified peptide extracts from untransformed tomato plants, those from transgenic plant possessed the ability to reduce the growth of several fungi in vitro. T2 transgenic plants were selected and tested for resistance against Fusarium sp. and Phytophthora infestans. The transgenic lines were more resistant to infection by these pathogens than the control plants.
PDF Abstract XML References Citation
How to cite this article
Z. Zainal, E. Marouf, I. Ismail and C.K. Fei, 2009. Expression of the Capsicuum annum (Chili) Defensin Gene in Transgenic Tomatoes Confers Enhanced Resistance to Fungal Pathogens. American Journal of Plant Physiology, 4: 70-79.
DOI: 10.3923/ajpp.2009.70.79
URL: https://scialert.net/abstract/?doi=ajpp.2009.70.79
DOI: 10.3923/ajpp.2009.70.79
URL: https://scialert.net/abstract/?doi=ajpp.2009.70.79
INTRODUCTION
Plants have evolved a variety of different mechanisms to cope with the constant threat posed by phytopathogenic microorganisms. For example, they have developed physical barriers and antimicrobial compounds that form in advance of a pathogenic attack (Van Loon et al., 2006). After infection by pathogens, these constitutive defenses are then supported by other induced mechanisms. For example, the cell wall can be reinforced by oxidative cross-linking. Additionally, hypersensitive cell death is triggered to isolate the pathogens from the healthy part of the plant causing the production of antimicrobial compounds such as phytoalexins and Pathogenesis-Related (PR) proteins (Epple et al., 1997; Gara et al., 2003). During recent years, it has become evident that, in addition to the above-mentioned defense mechanisms, several families of small, cysteine-rich, basic polypeptides play a role in the defense of plants against microorganisms (Stec, 2006). These are similar to the variety of antimicrobial polypeptides in epithelial tissues and macrophages that support the vertebrate immunoglobulin system and antimicrobial polypeptides located in the fat body that serve as part of the insect immune response (Aerts et al., 2006). In plants, this group of polypeptides has been classified into eight families that encompass thionins, defensins, lipid transfer proteins, hevein-type peptides, knottin-like peptides and snakins (Lay et al., 2003).
Plant defensins were originally termed γ-thionins because they have a similar size (5 kDa) and the same number of disulfide bridges (four) as α- and β-thionins. However, further studies revealed that γ-thionins were structurally different from those of α- and β-thionins and rather similar to insect and mammalian defensins in terms of its structure and function. Thus, this class of plant peptides was termed plant defensins (Aerts et al., 2007). The first members of this protein family have been isolated from wheat and barley and named γ-hordothionin and γ-purothionin. Since then, a variety of plant defensins have been isolated and characterized from many plant species including monocotyledons and dicotyledons. Plant defensins have 45-54 amino acid residues containing eight conserved cysteine residues linked to disulfide bridges. The global structure of plant defensins is composed of a Cysteine-Stabilized αβ motif (CSαβ motif) consisting of an α-helix and a triple-stranded β-sheet, organized in a βαββ architecture and stabilized by four disulfide bridges (Thevissen et al., 2003; Liu et al., 2006).
Plant defensins have been detected in different organs of plants such as leaves, flowers, pods, seeds and tubers, although, most plant defensins isolated to date are seed-derived (Wong and Ng, 2005; Song et al., 2005; Abre and Melane, 2008). Furthermore, the expression of some defensin genes is developmentally regulated, while others are greatly elevated in response to biotic and abiotic external stimuli. Defensin genes induced upon pathogen infection have been identified in peas, pepper and Arabidopsis (Mirouze et al., 2006. In Arabidopsis, at least five different defensin genes (pdf) have been identified; for example pdf 2.3, expressed constitutively in seedling, rosettes and flowers and pdf 1.2, induced by methyl jasmonate as well as by a fungal pathogen (Mirouze et al., 2006; Oh et al., 1999; Hanks et al., 2005; Manners et al., 1998). In addition, transgenic tobacco plants expressing the radish seed antifungal protein 1 (Rs-AFP1) exhibited enhanced resistance to fungal pathogens. Thus, plant defensins are considered to play an important role in plant defense mechanisms due to their antimicrobial activity in vitro and enhanced resistance of transgenic plants against phytopathogens. However, it has not been clearly demonstrated whether the variety of plant defensin isoforms differ in biological significance (Park et al., 2002). In this study, we report the generation of transgenic tomato plants (Lycopersicon esculentum cv. Alisa craig) constitutively expressing the chili defensin (cdef1) gene, which exhibit enhanced resistance against fungi.
MATERIALS AND METHODS
Cloning
The chili defensin coding sequence was amplified by PCR from the corresponding cDNA with following primers:
cd.1: 5´ GGG GGG GGA TCC ATG GCT GGC TTT TCC AAA GTA 3´ cd.2: 5´ GCC GCC GAG CTC TTA AGC ACA GGG CTT CGT GCA 3´ |
Primer cd.1 introduced a BamHI site at the start codon without altering the nucleotide sequence of the coding region and primer cd.2 introduced an SstI site just behind the stop codon. The PCR product was digested with BamHI and SstI (Promega), gel-purified, then cloned into the vector pBI121 (Jefferson, 1987). The correct sequence was verified by DNA sequencing and the recombinant vector was designated pBI-CD.
Generation of Transgenic Tomato Plants
The recombinant plasmid was transferred to Agrobacterium tumefaciens strain LBA 4404 using the freeze-thaw method and used for tomato transformation. Transformation of tomatoes (Lycopersicon esculentum cv. Alisa craig) was carried out according to the methods of Ling et al. (1998). Cotyledons and hypocotyls (10 days after germination) were used as the source of explant material for co-cultivation with A. tumefaciens. Tissue was selected on 2ZI (Murashige-Skoog (MS) salts, R3 vitamins, 2 mg L-1 zeatin, 0.1 mg L-1 indole-3-acetic acid (IAA) medium containing 150 mg L-1 gentamycin and 150 mg L-1 (timentin) and transformed calluses were excised and transferred to regeneration medium. Upon regeneration, plantlets were rooted with antibiotic selection and transferred to soil.
Southern Blots
Genomic DNA from transformed and non-transformed plants was isolated following the method described by Tabaeizadeh et al. (1999). DNA (30 μg) was digested overnight with HindIII, electrophoresed on a 0.7% agarose gel and transferred to nylon membranes (Hybond N+, Amersham Biosciences). Membranes were hybridized with a 613 bp 32P-labeled probe derived from the purified DNA fragment encompassing the CaMV 35S promoter and the chili defensin sequence. Hybridization and autoradiography procedures were carried out according to Sambrook et al. (1989).
Northern Blots
Total RNA was prepared from leaf tissues as described by Chomczynski and Sacchi (1987). To start, 20 μg of total RNA was separated on a denaturing 1.2% agarose gel (Ausubel et al., 1994). Ethidium bromide was included in the gel to verify equal loading of RNA. After transfer to membranes (Hybond N+, Amersham Biosciences), filters were hybridized with a 228 bp 32P-labeled probe, prepared from the corresponding cDNA. Filters were washed with 2xSSC, 0.1% SDS at room temperature for 10 min each, then once with 1xSSC, 0.1% SDS at room temperature for 15 min. Finally, the membranes were washed at high stringency with 0.1xSSC, 0.1% SDS at 65°C for 15 min. Filters were exposed to film (Kodak) at -70°C for 2 to 6 days.
Western Blots
Total protein from transgenic and control plants were extracted according to Andresen et al. (1992). Protein extracts were subjected to a polypeptide SDS-PAGE (Schagger and Von Jagow, 1987) and transferred to a nitrocellulose membrane (Sigma, 0.2 μm pore size) by electroblotting using a Hofer miniVE Blotter (Amersham Biosciences). Membranes were incubated with an antibody raised in rabbits against chili defensin and the bands were visualized with a goat anti-rabbit IgG conjugated with alkaline phosphatase (Promega). Western blot and immunodetection conditions were set according to the methods described by Sambrook et al. (1989).
Production of Antibody
Polyclonal antibodies were raised in rabbits using a purified CD obtained from the expression of the chili defensin gene cloned in plasmid pMALc2x (New England Biolabs) and expressed in E. coli strain TB1. The recombinant fusion protein was purified by amylose-affinity chromatography. All procedures were performed according to manufacturer’s protocols.
Disease Resistance Analysis
Seeds from the control (non-transgenic plant) and transgenic plants (T2 generation) were surface-sterilized and grown on ½ MS, 2% w/v sucrose and 75 μg mL-1 gentamycin and grown at 25°C in a 16 h light/8 h dark regime. After 1 month, surviving plants were transferred to soil and grown for 1 month at 25°C. Disease tests were performed by applying three needle-prick wounds to the leaves and covering the fresh wounds with 5 μL drops of a mixture of Phytophthora infestans and Fusarium sp. spores (conidial concentration was adjusted to 5x105 spores per mL using a hemocytometer and Tween 20 was added to a concentration 0.05% just before inoculation). Plants were covered with a plastic bag and kept at high humidity for 2 days to stimulate infection. Disease symptoms were checked 7 days after infection (Thomma and Broekaert, 1998).
For the in vitro assay, crude protein was extracted according to Francois et al. (2004). Thirty grams of tomato leaves (2 month-old plants, T2 generation plants and control plants) were frozen in liquid nitrogen, homogenized in extraction buffer (10 mM KCl, 9.6 mM NaH2PO4.2H2O, 15.2 mM Na2HPO4.2H2O, 150 mM NaCl, pH 7.0, 50 mL) and total protein extracts were boiled at 100°C for 10 min. After centrifugation at 13,000 rpm for 30 min, the supernatant was precipitated with ammonium sulfate at 70% relative saturation. The precipitated protein fraction was collected by centrifugation at 13,000 rpm for 30 min at 4°C. The pellet dissolved in 2 mL of dH2O and dialyzed overnight at 4°C against distilled water using 2000 kDa cut-off dialysis tubing (Sigma) and filter sterilized (0.2 μm). Protein concentration was quantified using the Bradford assay with bovine serum (BSA) as standard.
Antifungal activity of crude protein isolated from leaves was determined according to Lai et al. (2002). Plant extracts (serial two fold dilution, 2, 4, 8, 32, 64 and 128 mg L-1) were pipetted into the wells of a 96 well microtiter plate (TPP, Immonumaxi) containing 50 μL of one quarter strength Potato Dextrose Yeast extract (PD medium). After mixing, 50 μL of fungal spores (Phytophthora infestans, Fusarium sp., Colletotrichum gloeosporioides and Curvularia sp.; 40,000 spores mL-1 of water) were added to each well and the plates incubated in the dark at room temperature. As a control, protein extracted from a control plant (non-transgenic) was applied to all wells. Fungal spore germination and growth were measured with a microplate reader at 595 nm from 3 days to 1 week after inoculation.
RESULTS AND DISCUSSION
The pBI-CD plasmid, harboring 228 bp of the chili defensin cDNA under the control of a CaMV 35S promoter, was used for Agrobacterium tumefaciens mediated transformation and selected for gentamycin resistance (Fig. 1). For further characterization, 49 plants (T0 generation) originating from independent transformation events were grown.
In order to confirm the integration of cdef1 in the putative transgenic lines, Southern blots of genomic DNA digested with HindIII were performed. The DNA blot is shown in Fig. 2 indicates that selected lines were derived from independent transformation events. In line T5 and T13, two signals were prominent, reflecting the integration of two copies of the transgene in the genome of the transgenic plants. In lines T2, T9 and T12, only one signal was observed, which shows the presence of one transgene in the genomes of transgenic plants. No hybridization signal was detected in control plants.
Transcription of the integrated cdef1 was analyzed by Northern blot by hybridization with a chili defensin-specific probe. This indicated that the selected lines analyzed (T2, T5, T9, T12 and T13) stably expressed the cdef1 transgene as shown in Fig. 3, which confirms that cdef1 has been integrated into the tomato genome and expresses mRNA in transgenic plants. Expression of cdef1 was also analyzed by immuno-blot analysis of leaf crude protein fractions using the raised antibody. Protein extracts of leaves from transformants and non-transgenic control plants were electrophoresed on a SDS-PAGE gel, transferred to a nitrocellulose membrane and incubated with antibodies raised against chili defensin. The raised antibody detected a protein band with an estimated molecular mass 5 kDa as compared to a pre-stained molecular mass standard (Fig. 4). No correlation could be found between the number of inserts and transcript or protein levels.
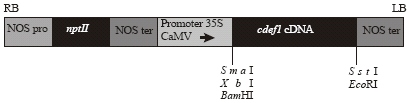 | |
| Fig. 1: | Schematic diagram of the pBI-CD vector constructed for transformation of cdef1 in transgenic tomato plants. LB: Left border, RB: Right border, nptII: Neomycin phosphotransferase gene conferring kanamycin resistance, hptII: Conferring hygromycin resistance, Nos pro: Nophaline synthase promoter, Nos ter: Terminator, LacZ: β-D-galactosidase gene |
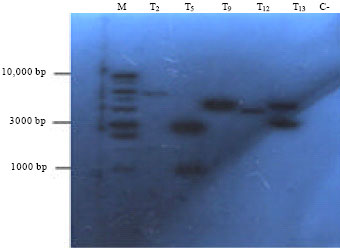 | |
| Fig. 2: | Autoradiogram of the Southern hybridization. The different signal patterns suggest the multiple integration of cdef1 in plant genome. Lane M: 1 kb marker; Lane-1-5: Transgenic plants; Lane C-: Non-transformed control |
 | |
| Fig. 3: | Autoradiogram of the Northern hybridization. Total RNA isolated from leaves oftransgenic tomato was separated on a 1.2% denaturing agarose gel, blotted onto a nylon membrane and hybridized with radio-labeled probes as indicated. Lane T2-T13: Transgenic plants; Lane C-: Non-transformed plant |
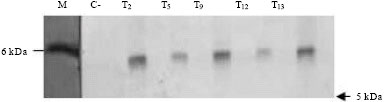 | |
| Fig. 4: | Western blot analysis of plant defensin in the leaves of transgenic tomato. Lane M: Protein marker; Lane C-: Non-transformed plant; Lane T2-T13: Transgenic tomato plants |
| Table 1: | Progeny segregation of survived lines |
 | |
| The inheritance of the transgene was tested by estimating the percentage of the seedlings that grew on gentamycin | |
| Table 2: | Antifungal activity of leaf extracts from transgenic tomato plants (T) expressing chili defensin |
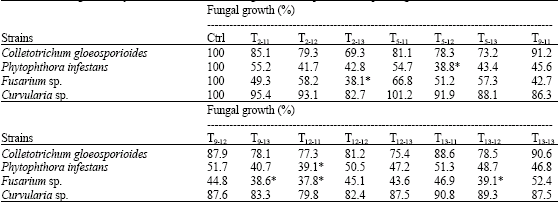 | |
| Result are summarized from four replicated experiments and expressed as the percentage of fungal growth over extracts from untransformed (Ctrl) tomato plants. *Indicate significant differences between leaf extracts from transgenic plants and control extracts | |
Primary transformants (line T2, T5, T9, T12 and T13) were allowed to self-fertilize and seeds of the subsequent (segregation) T1 generation were selected with gentamycin(75 mg L-1). Among the transgenic plants tested, the progeny population from 4 lines (T2, T5, T9, T12) showed conventional Mendelian inheritance (3:1), while line T13 showed a 5:1 ratio (Table 1). Gentamycin and resistant T1 seedlings were grown in a greenhouse to produce T2 seeds.
To test the antifungal activity of cdef1, partially purified extracts from five independent transgenic plants (T2 generation of lines T2, T5, T9, T12 and T13, three plants for each line) were prepared. Crude protein extract fractions were tested for in vitro antifungal activity against a pathogenic fungus using a quantitative microplate assay. Crude extracts were added to cultures of Phytophthora infestans, Fusarium sp., Colletotrichum gloeosporioides and Curvularia sp. and their effects on fungal growth were assessed. When compared to similar extracts from untransformed tomato leaves (control), the transformed leaves expressing the chili cdef1 resulted in significant growth inhibition of the four fungi tested. In some lines (line T2-13, T5-12, T9-13, T12-11 and T13-12), up to 60% fungal growth inhibition was observed (Table 2), while in other samples (lines T2-12, T5-11, T5-13, T12-12, T12-13 and T13-13), growth decreased up to 50%. This result indicates that the chili defensin (cdef1) possesses potent antifungal activity.
To assess whether transgenic tomato plants could acquire enhanced resistance to fungal pathogens, 30 transgenic tomato plants (T2 generation, 6 plants from each line) were selected using gentamycin. Phytophthora infestans and Fusarium sp. fungal pathogens were used to infect transgenic and control plants and ten days after infection, disease symptoms started to appear on the control plants, but not the transgenic plants. Fourteen days after infection, the control plants displayed disease symptoms such as brown lesions, necrotic spots and leaf wilting. However, transgenic lines remained relatively healthy without appreciable disease symptoms, with the exception of a slight yellowing of the bottom leaves that had contacted the infecting fungus (Fig. 5a-c).
Plants have developed various defense mechanisms against environmental stresses such as infection by foreign pathogens. Upon contact with pathogens, a signal is transmitted and a number of antimicrobial proteins are expressed to fight off pathogens.
 | |
| Fig. 5: | Disease symptoms on leaves of control and transgenic plants. Picture were taken 14 days after infection. (a and b) The control plants showed disease symptoms such as brown lesions, necrotic spots and leaf wilting. (c) The transgenic lines remained relatively healthy without appreciable disease symptoms |
Plant defensins were suggested to be involved in plant defense mechanisms because these proteins have either antifungal or antibacterial properties (Moreno et al., 1994; Terras et al., 1995). In addition, the steady-state levels of several plant defensin transcripts increase upon infection (Chiang and Hadwiger, 1991; Terras et al., 1995).
To validate the presumed role of chili defensin in host defense, we generated transgenic tomato plants containing cdef1 by Agrobacterium mediated transformation. The CaMV promoter was used in this study as it is extremely strong and expected to drive high levels of chili defensin expression in transgenic tomato plants. Southern blots were generated out to further confirm T-DNA integration and results clearly show variation in pattern of the integration (one to two copies) among transgenic plants, indicating independent transformation events. The 5 kDa peptide, which corresponds to the mature form of the chili defensin peptide was detected in the total protein fraction extracted from transgenic tomato leaves by western blot analysis. These chili-defensin extracts significantly reduced the growth of phytopathogenic fungi. However, due to the use of partially-purified plant extracts rather than purified peptides, it remains possible that some of the enhanced antifungal activity associated with the transgenic extracts was due to synergistic interactions between the chili defensin and endogenous tomato compounds. In this context, the plant extracts tested in this study had been partially purified to remove enzymes through protein denaturation and size fractionation (> 2000 Da). Therefore, it is unlikely that these extracts contained hydrolytic enzymes such as chitinases and glucanases, which have been speculated to interact synergistically with defensins (Lai et al., 2002). We cannot rule out the possibility of interactions between the chili defensin and other small, heat-stable compounds from tomato that may have been in the extracts. Because of the difficulties associated with obtaining proper disulfide linkages, chemical synthesis of defensin peptides is not a viable option (Lai et al., 2002).
Present analysis of the antifungal activities of the chili defensin gene is consistent with previous suggestions that plant defensins participate in host defense responses against pathogens. For example, extracts from tobacco plants expressing pea defensin genes significantly reduced the growth of several phytopathogenic fungi including Ascochyta pinodes, A. pisi, Fusarium oxysporum, F. solani, Alternaria alternate, Ascochyta lentis, Leptosphaeria maculans and Trichoderma reesei (Lai et al., 2002). Meyer et al. (1996) also showed that purified chili fruit-specific defensin was effective in suppressing the growth of the fungi Fusarium oxysporum and Botrytis cinerea. Florack et al. (1994) demonstrated that the hordothionin (HTHs) from transgenic tobacco plants were biologically active and inhibited the growth of Clavibacter michiganesis subsp. michiganesis. However, considerable variation was observed in the effectiveness of chili defensin against different fungal strains. Chili defensin had a higher antifungal activity against Phytophthora infestans and Fusarium sp. compared to Colletotrichum gloeosporioides and Curvularia sp. This shows that the antifungal activity of plant defensins is strongly dependent on the target fungus. Park et al. (2002) showed that antifungal activities of the BSD1 (Brassica Stamen-specific plant Defensin 1) protein by radial growth-inhibition assays were most effective against Neurospora crassa, Fusarium oxysporium, Phytophthora parasitica and Alternaria solani and did not show growth inhibition against Botrytis cinrea, Pythium apharudermatum and Rhizoctonia solani.
The transgenic T2 lines also exhibited enhanced resistance to the fungal pathogen. Different levels of fungi resistance in T2 transformants may be caused by two factors. First, transcriptional or post-transcriptional silencing of the transgenes could have affected the level of disease resistance. The silencing of transgenes has been reported for many plant species in transgenic rice plants harboring a chitinase gene, silencing of the transgene was reported in T2 and T3 progeny (Chareonpornwattana et al., 1999; Hart et al., 1992). Second, positional effects within the genome may cause differences in transgene expression (Meyer, 1995).
Additional evidence for a role of plant defensins in plant defense came from the analysis of the disease tolerance of transgenic tobacco plants constitutively expressing Rs-AFP2. The leaves of such Rs-AFP2-expressing plants displayed a significantly decreased susceptibility to Alternaria longipes infection relative to untransformed control plants (Terras et al., 1995). It is already known that transgenic rice plants overproducing wasabi defensin are expected to possess a durable and wide-spectrum resistance (i.e., field resistance) against various rice blast races (Kanzaki et al., 2002). A possible role for plant defensins in fungal pathogen defense is also suggested by the finding that the over expression of a pea defensin gene (DRR230-a) in canola reduced disease symptoms following infection with Leptosphaeria maculans (Wang et al., 1999). However, additional research will be required to determine whether expression of chili defensin is an effective means of increasing disease resistance.
Finally, conclusive evidence for the involvement of chili defensins in the plants defense response could be obtained by inactivating these genes or expressing different transgenes under the control of a single promoter sequence. With the development of effective genetic transformation methods, this is now feasible in plants through antisense or gene tagging strategies.
ACKNOWLEDGMENTS
This study was partially supported by a grant from the IRPA (Grant No. IRPA: 09-02-02-0081-EA209) and UKM-OUP-KPB-33-171/2008 awarded to Dr. Ismanizan Ismail and Dr. Zamri Zainal. Authors thank to Tee Choon Yang for critical Aertsing of this manuscript.
REFERENCES
- Abre de, B. and A.V. Melane, 2008. Vv-AMP1, a ripening induced peptide from Vitis vinifera shows strong antifungal activity. BMC Plant Biol., 8: 75-75.
CrossRef - Aerts, A.M., I.E. Francois, L. Bammens, B.P. Cammue, B. Smets, J. Winderickx, S. Accardo, D.E. De Vos and K. Thevissen, 2006. Level of M (IP) 2C sphingolipid affects plant defensin sensitivity, oxidative stress resistance and chronological life span in yeast. FEBS. Lett., 580: 1903-1907.
Direct Link - Aerts, A., K. Thevissen, S. Bresseleers, J. Sels, P. Wouters, B. Cammue and I. Francois, 2007. Arabidopsis thaliana plants expressing human beta-defensin-2 are more resistant to fungal attack: Functional homology between plant and human defensins. Plant Cell Rep., 26: 1391-1398.
Direct Link - Andresen, I., W. Becker, K. Schluter, J. Burges, B. Parthier and K. Apel, 1992. The identification of leaf thionin as one of the main jasmonate-induced proteins of barley (Hordeum vulgare). Plant Mol. Biol., 19: 193-204.
CrossRefDirect Link - Chareonpornwattana, S., K.V. Thara, L. Wang, S.K. Datta, W. Panbangred and S. Muthukrishnan, 1999. Inheritance, expression and silencing of a chitinase transgene in rice. Theor. Applied Genet., 98: 371-378.
CrossRefDirect Link - Chiang, C.C. and L.A. Hadwiger, 1991. The Fusarium solani-induced expression of a pea gene family encoding high cysteine content proteins. Mol. Plant Microbe Interact., 4: 324-331.
Direct Link - Chomczynski, P. and N. Sacchi, 1987. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Annu. Rev. Biochem., 162: 156-159.
CrossRefPubMedDirect Link - Epple, P., K. Apel and H. Bohlmann, 1997. Overexpression of an endogenous thionin enhances resistance of Arabidopsis against Fusarium oxysporum. Plant Cell, 9: 509-520.
CrossRefDirect Link - Florack, D.E.A., W.G. Dirkse, B. Visser, F. Heidekamp and W.J. Stiekema, 1994. Expression of biologically active hordothionins in tobacco. Effects of pre- and pro-sequences at the amino and carboxyl termini of the hordothionin precursor on mature protein expression and sorting. Plant Mol. Biol., 24: 83-96.
CrossRefDirect Link - Francois, I.E.J.A., W.V. Hemelrijck, A.M. Aerts, P.F.J. Wouters, P. Proost, W.F. Broekaert and B.P.A. Cammue, 2004. Processing in Arabidopsis thaliana of a heterologous polyprotein resulting in differential targeting of the individual plant defensins. Plant Sci., 166: 113-121.
CrossRef - De Gara, L., M.C. de Pinto and F. Tommasi, 2003. The antioxidant systems vis-a-vis reactive oxygen species during plant-pathogen interaction. Plant Physiol. Biochem., 41: 863-870.
CrossRefDirect Link - Hanks, J.N., A.K. Snyder, M.A. Graham, R.K. Shah, L.A. Blaylock, M.J. Harrison and D.M. Shah, 2005. Defensin gene family in Medicago truncatula: Structure, expression and induction by signal molecules. Plant Mol. Biol., 58: 385-399.
Direct Link - Jefferson, R.A., 1987. Assaying chimeric genes in plants: The GUS gene fusion system. Plant Mol. Biol. Rep., 5: 387-405.
CrossRefDirect Link - Lai, F.M., C. DeLong, K. Mei, T. Wignes and P.R. Fobert, 2002. Analysis of the DRR230 family of pea defensins: Gene expression pattern and evidence of broad host-range antifungal activity. Plant Sci., 163: 855-864.
CrossRef - Lay, F.T., H.J. Schirra, M.J. Scanlon, M.A. Anderson and D.J. Craik, 2003. The three-dimensional solution structure of NaD1, a new floral defensin from Nicotiana alata and its application to a homology model of the crop defense protein alfAFP. J. Mol. Biol., 325: 175-188.
CrossRef - Liu, Y.J., C.S. Cheng, S.M. Lai, M.P. Hsu, C.S. Chen and P.C. Lyu, 2006. Solution structure of the plant defensin VrD1 from mung bean and its possible role in insecticidal activity against bruchids. Proteins, 63: 777-786.
Direct Link - Mirouze, M., J. Sels, O. Richard, P. Czernic and S. Loubet et al., 2006. A putative novel role for plant defensins: A defensin from the zinc hyper-accumulating plant, Arabidopsis halleri, confers zinc tolerance. Plant J., 47: 329-342.
Direct Link - Park, H.C., Y.H. Kang, H.J. Chun, J.C. Koo and Y.H. Cheong et al., 2002. Characterization of a stamen-specific cDNA encoding a novel plant defensin in Chinese cabbage. Plant Mol. Biol., 50: 59-69.
PubMedDirect Link - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA., ISBN-13: 9780879695774, Pages: 397.
Direct Link - Song, X., J. Wang, F. Wu, X. Li, M. Teng and W. Gong, 2005. cDNA cloning, functional expression and antifungal activities of a dimeric plant defensin SPE10 from Pachyrrhizus erosus seeds. Plant. Mol. Biol., 57: 13-20.
Direct Link - Stec, B., 2006. Plant thionins-the structural perspective. Cell Mol. Life Sci., 63: 1370-1385.
Direct Link - Tabaeizadeh, Z., Z. Agharbaoui, H. Harrak and V. Poysa, 1999. Transgenic tomato plants expressing a Lycopersicon chilense chitinase gene demonstrate improved resistance to Verticillium dahliae race 2. Plant Cell Rep., 19: 197-202.
CrossRef - Thevissen, K., K.K. Ferket, I.E. Francois and B.P. Cammue, 2003. Interaction of antifungal plant defensins with fungal membrane components. Peptides, 24: 1705-1712.
CrossRef - Van Loon, L.C., M. Rep and C.M.J. Pieterse, 2006. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol., 44: 135-162.
CrossRefDirect Link - Wong, J.H. and T.B. Ng, 2005. Sesquin, a potent defensin-like antimicrobial peptide from ground beans with inhibitory activities toward tumor cells and HIV-1 reverse transcriptase. Peptides, 26: 1120-1126.
Direct Link - Schagger, H. and G. von Jagow, 1987. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal. Biochem., 166: 368-379.
CrossRefPubMedDirect Link