
M. Naeem
Department of Botany, Aligarh Muslim University, Aligarh-202 002, India
M. Masroor A. Khan
Department of Botany, Aligarh Muslim University, Aligarh-202 002, India
J.B. Morris
United States Department of Agriculture, Agricultural Research Service, Plant Genetic Resources Conservation Unit, 1109 Experiment Street, Griffin, GA 30223-1797, United States of America
American Journal of Plant Physiology
Year: 2009 | Volume: 4 | Issue: 2 | Page No.: 58-69







ABSTRACT
Hyacinth bean (Lablab purpureus L.) accessions of different origins received from USDA, ARS, Plant Genetic Resources Conservation Unit, Griffin, GA, USA were evaluated for agrobotanical attributes, enzyme activities, nutraceuticals and quality in calcium deficient soil of Aligarh, Western Uttar Pradesh, India. Fresh and dry weights per plant, leaf-area, number and dry weight of nodules per plant, net photosynthetic rate, stomatal conductance and transpiration rate, total chlorophyll and carotenoid content, activities of nitrate reductase and carbonic anhydrase, leaf - N, P, K and Ca contents and nodule-nitrogen and leghaemoglobin contents, respectively were analyzed at 60, 90 and 120 day after sowing. Photosynthesis was measured only at 90 DAS. Yield attributes including pod number per plant, seed number per pod, 100-seed weight and seed-yield per plant were recorded at harvest (150 DAS). Protein and carbohydrate content as well as tyrosinase activity in hyacinth bean seeds were also determined. Among the five accessions, EC-497619 (A4) showed superior performance over the rest of the accessions. Accession A4 showed the highest values for growth, yield, physiological, biochemical and quality attributes in comparison to the other accessions. Net photosynthetic rate, stomatal conductance and transpiration rate were found maximum in the A4 accession. Chlorophyll and carotenoid content were also reported higher in accession A4. Accession A4 showed higher nitrate reductase and carbonic anhydrase activities than the other accessions. Nodule-nitrogen and leghaemoglobin content ranged from 5.267-5.314% and 0.110-0.130 mM, respectively. Mineral profiles, viz., nitrogen, phosphorus, potassium and calcium content varied from 3.610-3.643, 0.338-0.356, 3.020-3.124 and 1.764-1.804%, respectively. Seed protein of all accessions varied from 24.70-25.06%. Carbohydrate content ranged from 50.83-53.16% across all accessions tested. Accession A4 produced the highest tyrosinase activity in the seeds.
PDF Abstract XML References Citation
How to cite this article
M. Naeem, M. Masroor A. Khan and J.B. Morris, 2009. Agrobotanical Attributes, Nitrogen-Fixation, Enzyme Activities and Nutraceuticals of Hyacinth Bean (Lablab purpureus L.): A Bio-Functional Medicinal Legume. American Journal of Plant Physiology, 4: 58-69.
DOI: 10.3923/ajpp.2009.58.69
URL: https://scialert.net/abstract/?doi=ajpp.2009.58.69
DOI: 10.3923/ajpp.2009.58.69
URL: https://scialert.net/abstract/?doi=ajpp.2009.58.69
INTRODUCTION
The vegetarian populations in India consume large amounts of legumes, particularly, vegetable beans in their diet. The beans are naturally rich in carbohydrates, proteins, fat and fibers as well as minerals including calcium, phosphorus and iron. Furthermore, several legumes have tremendous potential as nutraceuticals because of their healing properties (Morris, 1999, 2003). Hyacinth bean belongs to the family Fabaceae and is grown in the tropical and sub-tropical India. Hyacinth bean (Lablab purpureus L.) has great potential as medicinal legume. Among the legumes, hyacinth bean constitutes an important source of therapeutic agents used in the modern as well as traditional systems of medicine (Morris, 1996, 1999, 2003). It carries tremendous healing potential. In fact, it is considered a multipurpose crop since it is used for food, forage, soil improvement, soil protection and weed control (Shivashankar and Kulkarni, 1989; Karachi, 1998; Morris, 1997, 2003; Pengelly and Maass, 2001; Maass, 2006). The young pods and tender beans are used as vegetables in India and tropical and warm temperate Asia. It is also been known for its use as a green manure and produces edible young pods, dried seeds, leaves and flowers (Morris, 1997, 2003). The seeds are used as a laxative, diuretic, anthelmintic, antispasmodic, aphrodisiac, anaphrodisiac, digestive, carminative, febrifuge and stomachic (Chopra et al., 1986; Kirtikar and Basu, 1995). Hyacinth beans contain fiber which is known to prevent cancer, diabetes, heart disease, obesity and is used as a laxative (Beckstrom-Sternberg and Duke, 1994). Hyacinth bean contains the potential breast cancer fighting a flavonoid known as kievitone (Hoffman, 1995). The flavonoid, genistein found in hyacinth bean may play a role in the prevention of cancer (Kobayashi et al., 2002) and as a chemotherapeutic and/or chemopreventive agent for head and neck cancer (Alhasan et al., 2001). Tryrosinase (polyphenol oxidase) is present in plant tissue and is important in fruit and vegetable processing as well as storage of processed foods. Prevention of browning of foods, enzymatic or nonenzymatic, has long been a concern of food scientists (Matheis, 1987; Sanchez-Ferrer et al., 1995; Paul and Gowda, 2000). Hyacinth bean contains tyrosinase, which has potential for the treatment of hypertension in humans (Beckstrom-Sternberg and Duke, 1994). However, it is not being used to its full potential.
Keeping in mind the importance of this medicinal legume as a multipurpose crop, the aim of the present study is to investigate the performance of five hyacinth bean accessions of different origin, for various agrobotanical attributes, physiological, biochemical and quality attributes under the agro-climatic conditions of Aligarh, Western Uttar Pradesh in calcium deficient soil.
MATERIALS AND METHODS
Plant Material and Growth Conditions
Healthy seeds of hyacinth bean (Lablab purpureus L.) accessions, namely EC-497617 (Australia), EC-497616 (China), EC-497615 (Egypt), EC-497619 (Iran) and EC-497618 (Kenya) were received from the USDA, ARS, Plant Genetic Resources Conservation Unit, Griffin, GA, USA and denoted as A1, A2, A3, A4 and A5, respectively. Healthy seeds of uniform size were selected and their viability was tested using 1% tetrazolium salt. The seeds were surface sterilized with 95% ethyl alcohol for five minutes and then washed thoroughly with distilled water.
Healthy and viable Rhizobium culture (Rhizobium sp.), compatible for hyacinth bean, was obtained from Culture Laboratory, Government Agriculture Farm, Quarsi, Aligarh. The Rhizobium culture was prepared according to Subba Rao (1972). Two hundred grams of colorless Gum Arabic (coating material) and 50 g of sugar were dissolved in 500 mL warm water. After cooling, 100 g of Rhizobium culture was mixed with the solution. The required amount of seeds were mixed vigorously with the inoculum until they were evenly coated by the inoculum mixture. The inoculated seeds were placed in a clean tray and dried in shade prior to sowing. Then seeds were sown directly at a depth of 2 cm in soil into earthen pots containing a homogenous mixture of soil and farmyard manure (5:1). Initially five plants were maintained in each pot, but later were reduced to one healthy plant. The soil was maintained at proper moisture to ensure better seed germination. Physico-chemical characteristics of the soil were: texture-sandy loam, pH (1:2), 7.2, E.C. (1:2), 0.46 m mhos cm–1, available N, P and K 98.39, 6.78 and 142.9 mg kg–1 soil, respectively and calcium carbonate 0.12%. The soil samples were analyzed at the Government Soil Testing Laboratory, Quarsi Farm, Aligarh. Prior to seed sowing, a uniform basal dose of fertilizers (10 mg P and 120 mg Ca kg–1 soil) was applied. The source of phosphorus and calcium were potassium dihdrogen orthophosphate and calcium chloride. However, being a leguminous crop, hyacinth bean was not supplied with nitrogen. The requirement of nitrogen is fulfilled by the crop by virtue of biological nitrogen fixation since Rhizobium was applied to the seeds. An out door pot experiment was conducted during 2002-2003 using a simple randomized complete block design in the net-house at the Botany Department, AMU, Aligarh (27° 52' N latitude, 78° 51' E longitude and 187.45 m altitude). The crop was sown on 20th September, 2002 and harvested on 20th February, 2003. Each accession was replicated three times. The pots were watered thoroughly and plants were grown in natural condition.
Measurement of Growth and Yield Attributes
At 60 (vegetative stage), 90 (flowering stage) and 120 (pod-filling stage) days after sowing, three plants of each accession were uprooted carefully and washed with running tap water to remove foreign particles. Fresh plant weight was recorded. Leaf area was measured by outlining leaves of sampled plants on graph paper and dry weight of these leaves was recorded. Leaf-area per plant was determined using leaf dry weight per plant and dry weight of those leaves for which the area was estimated (Watson, 1958). The root nodules of each plant were washed under tap water and counted. The plants were then dried at 80°C for 24 h and the dry weights of nodules and plants were recorded.
At harvest (150 DAS), six plants from each accession were uprooted randomly and were used for computing yield attributes including pod number per plant, seed number per pod, 100-seed weight and seed-yield per plant. Pods were threshed and cleaned. The seeds cleaned and counted. Seeds were sun dried for recording a more accurate 100-seed weight. Seed-yield was calculated accordingly.
The fresh leaves of each accession were used for analysis of various physiological and biochemical attributes except leaf-N, P, K and Ca contents.
Net Photosynthetic Rate, Stomatal Conductance and Transpiration Rate
Net photosynthetic rate, stomatal conductance and transpiration rate were measured on cloud-less clear days at 1100 h from fully expanded hyacinth bean leaves using a IRGA (Infra Red Gas Analyzer, LICOR 6200 Portable Photosynthesis System, Lincoln, Nebraska, USA). Before recording the measurement, the IRGA was calibrated and zero was adjusted approximately every 30 min during the measurement period. The atmospheric conditions during measurement were Photosynthetically Active Radiation (PAR) 1016±6 l μmol/m2/sec, relative humidity 60±3%, atmospheric temperature 22±1°C and atmospheric CO2 360 μmol mol–1. The ratio of atmospheric CO2 to intercellular CO2 concentration was constant. Leaves of each accession were enclosed in a 1 L gas exchange chamber for 60 sec. All attributes were measured three times for each accession. Photosynthesis was measured only at 90 days after sowing.
Total Chlorophyll and Carotenoid Content
Total chlorophyll and carotenoid content in fresh leaves were estimated using the method of Lichtenthaler and Buschmann (2001). The fresh tissues from interveinal leaf area were ground in a mortar and pestle containing 80% acetone. The Optical Density (OD) of the solution was read at 662 and 645 nm (chlorophyll a and b) and 470 nm (carotenoids) using a spectrophotometer (Spectronic 20D, Milton Roy, USA). Photosynthetic pigments were expressed as mg g–1 FW.
Nitrate Reductase Activity (NRA)
The enzyme activity was estimated by the intact tissue method developed by Jaworski (1971). Two hundred milligrams of fresh chopped hyacinth bean leaves were weighed and transferred to a plastic vial. Each vial contained 2.5 mL phosphate buffer (pH 7.5) and 0.5 mL potassium nitrate solution followed by the addition of 5% isopropanol. After incubation, 1% sulphanilamide and 0.02% N-(1-Naphthyl) ethylenediamine dihydrochloride (NED-HCL) was added. The OD of colour was read at 540 nm using a spectrophotometer. Nitrate reductase activity was expressed as nM NO2– g–1 FW h–1.
Carbonic Anhydrase Activity
The Carbonic Anhydrase (CA) activity in fresh leaves was analyzed using the method described by Dwivedi and Randhawa (1974). Two hundred milligram of fresh leaf pieces were weighed and transferred to petri plates. The leaf pieces were dipped in 10 mL of 0.2 M cystein hydrochloride for 20 min at 4°C. Four mL of 0.2 M sodium bicarbonate solution and 0.2 mL of 0.022% bromothymol blue was added to the homogenate. The reaction mixture was titrated against 0.05 N HCl using methyl red as indicator. Carbonic anhydrase activity was expressed as μM CO2 kg–1 leaf FW sec–1.
Nutrient Analysis
Leaf samples of each accession were digested according to the method used by Lindner (1944) for the estimation of leaf -N, P, K and Ca contents.
Leaf-Nitrogen Content
Leaf-nitrogen content was estimated using the method of Lindner (1944) as well. A 10 mL aliquot (peroxide-digested material) was taken in a 50 mL volumetric flask. To this, 2 mL of 2.5 N sodium hydroxide and 1 mL of 10% sodium silicate solutions were added to neutralize the excess of acid and to prevent turbidity, respectively. In a 10 mL graduated test tube, 5 mL aliquot of this solution was taken and 0.5 mL Nessler’s reagent was added. The contents of the test tubes were allowed to stand for 5 min for maximum colour development. The OD of the solution was read at 525 nm, using a spectrophotometer. The reading of each sample was compared with the standard calibration curve of ammonium sulphate to estimate the percent nitrogen content.
Leaf-Phosphorus Content
The method of Fiske and Subba Row (1925) was used to estimate the leaf-phosphorus content in the digested material. The same aliquot was used to determine the leaf-P content. A 5 mL aliquot was taken in a 10 mL graduated test tube where 1 mL molybdic acid (2.5%) was added carefully, followed by addition of 0.4 mL 1-amino-2-naphthol-4-sulphonic acid. When the colour of tube turned blue, the volume was made up to 10 mL with the addition of double distilled water. The solution was shaken for 5 min. The OD of the solution was read at 620 nm using a spectrophotometer.
Leaf-Potassium and Calcium Content
Potassium and calcium contents were analyzed using flame-photometrics. In the flame-photometer, the solution (peroxide-digested material) is discharged through an atomizer in the form of a fine mist into a chamber, where it is drawn into a flame. Combustion of the elements produces light of a particular wavelength (λmax for K = 767 nm (violet)). The light produced was conducted through the appropriate filters to impinge upon a photoelectric cell that activates a galvanometer. Both leaf potassium and calcium content in the same aliquot were estimated and recorded with the help of emission spectra using specific filters in a flame-photometer. Leaf-N, P, K and Ca content were expressed in percent on the dry weight basis.
Nodule-Nitrogen and Leghaemoglobin Content
Nodule-nitrogen content was also estimated by the method of Lindner (1944). Leghaemoglobin (Lb) content in fresh nodules was determined as described by Sadasivam and Manickam (2008). The solution’s OD was recorded at 556 and 539 nm. The Lb content was calculated using the following formula:
where, OD 556 and 539 represent absorbance (OD). Values recorded at 556, 539 nm, respectively and D is the initial dilution.
Seed-Protein Content
The seed protein content was estimated using the method of Lowry et al. (1951). Hyacinth bean seed was ground to a powder using a mortar and pestle. The seed powder was transferred to a mortar where 5% cold trichloroacetic acid (TCA) was present. Extracted protein was measured at 660 nm using a spectrophotometer. The reading was compared with a calibration curve obtained by using known dilution of standard egg albumin solution and the percent seed protein content was calculated on a dry weight basis.
Total Carbohydrate Content
Total carbohydrate content in seeds was analyzed as described by Sadasivam and Manickam (2008). One hundred milligrams of hyacinth bean powder was poured into a tube containing boiling sulphuric acid and centrifuged at 4000 rpm. Four milliliter of anthrone reagent was added and the resulted dark green colour was recorded at 630 nm. The reading was compared with the calibration curve obtained using a known dilution of a standard of glucose and the percent carbohydrate content was calculated on a dry weight basis.
Determination of Tyrosinase (Polyphenol Oxidase) Activity
The tyrosinase enzyme was extracted according to Paul and Gowda (2000) and assayed spectrophotometrically, using the procedure of Cosetang and Lee (1987). The enzyme assay mixture contained 0.9 mL of 0.05 M sodium acetate buffer (pH 4.5), 0.1 mL of substrate (L-3, 4-dihydroxy phenylalanine) (L-DOPA) and 10-100 μg of the enzyme. The optical density of coloured solution developed due to formation of the compound dopachrome was read at 480 nm. One unit of the enzyme activity corresponded to an amount of enzyme that caused an increase in the absorbance of 0.001 min–1 at 25°C. The reference cuvette contained all the ingredients except the enzyme in a final volume of 1 mL. The activity of tyrosinase was expressed as U mg–1 protein.
Statistical Analysis
The data were analyzed by one-way ANOVA. Mean values were analyzed at the 0.05 level of probability according to Gomez and Gomez (1984).
RESULTS
Growth and Yield Attributes
The mean data and Least Significant Differences (LSD) (p≤0.05) of agrobotanical attributes from five hyacinth bean accessions are presented in Table 1 and 2. Accession A4 followed by A1 had the greatest fresh and dry weights per plant at 60, 90 and 120 DAS. Accession A5 had the lowest fresh and dry weights at 60, 90 and 120 DAS. Both accession A4 and A1 produced the greatest leaf-area among all accessions at 60, 90 and 120 DAS.
| Table 1: | Growth attributes of five accessions of hyacinth bean (Lablab purpureus L.) studied at 60, 90 and 120 DAS |
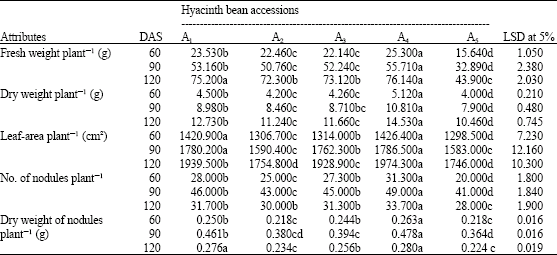 | |
| Mean values of 3 replicates. Mean values within a column followed by the same letter(s) are not significantly different (p≤0.05) | |
| Table 2: | Yield attributes of five accessions of hyacinth bean (Lablab purpureus L.) studied at 150 DAS |
 | |
| NS: Not Significant: Mean values of 3 replicates. Mean values within a column followed by the same letter(s) are not significantly different (p≤0.05) | |
Accession A5 produced the least amount of leaf-area at all growth stages (Table 1). Nodule number and dry weight were highest in accession A4 at 60, 90 and 120 DAS, while accession A5 produced the lowest number of nodules and nodule dry weight at similar stages (Table 1).
Table 1 indicates that accession A4 produced the highest number of pods per plant (45.2), 100-seed weight (4.5 g) and seed-yield (45.60 g) per plant, while accession A5 produced lowest yield (34.64 g). However, all accessions produced similar values for seed number per pod and did not significantly differ from each other (Table 2).
Physiological and Biochemical Attributes
The net photosynthetic rate was maximized in accession A4 followed by accession A1. Accession A5 had the lowest rate at 90 DAS (Fig. 1). It was observed that stomatal conductance and transpiration rate maximized in accession A4, while accession A1 was similar with that of A4. Accession A5 had the lowest transpiration rate and stomatal conductance (Fig. 1). Both accessions, A4 and A1 produced the highest chlorophyll content and were superior to the other accessions at 60, 90 and 120 DAS (Fig. 1). Both accessions A5 and A2, had similar chlorophyll content at 60, 90 and 120 DAS (p≤0.05). Maximum carotenoids were generated in the leaves of accessions A4 and A1 at 60 DAS followed by A3, A2 and A5 at all three DAS (Fig. 1). Maximum total chlorophyll (1.889 mg g–1) and carotenoid (0.568 mg g–1) content were found in all accessions at 90 DAS. Accession A4 had the maximum nitrate reductase activity and was similar to accessions A1 and A3. Accession A5 produced the lowest NR activity (Fig. 1). It was observed that A4 produced the highest carbonic anhydrase activity and accession A5 produced the lowest at all three growth stages of the hyacinth bean plant (Fig. 1). Nodule-nitrogen content was maximum in accession A4 (5.314, 4.439 and 3.435% at 60, 90 and 120 DAS).
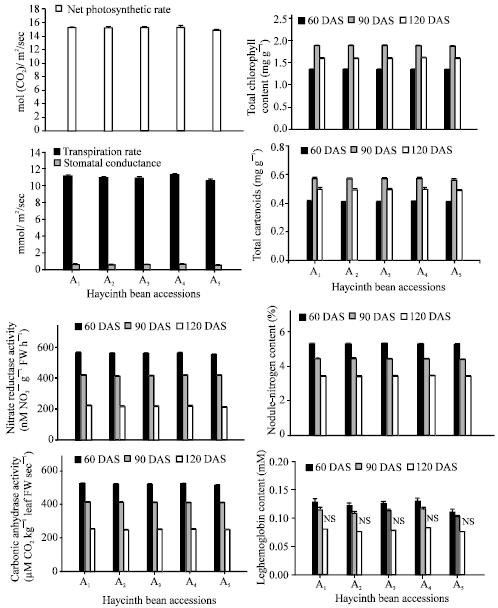 | |
| Fig. 1: | Changes in net photosynthetic rate, stomatal conductance and transpiration rate (90 DAS), total chlorophyll and carotenoid content, nitrate reductase activity, carbonic anhydrase activity, nodule-nitrogen content and leghaemoglobin content of hyacinth bean (Lablab purpureus L.) accessions studied at 60, 90 and 120 DAS. Error bars (τ) show LSD at 5% level |
However, accessions A1 (5.310, 4.434 and 3.430%) and A4 (5.314, 4.439 and 3.435%) were not significantly different and were almost equal in nodule-nitrogen content at 60, 90 and 120 DAS (Fig. 1). Accession A5 produced the lowest nodule nitrogen content at all three stages. In the present study, maximum nodule-nitrogen content was found at 60 DAS in all the accessions (Fig. 1).
Accessions A4 and A1 produced the highest concentration of leghaemoglobin content at 60 and 90 DAS while accession A5 produced the least (Fig. 1). Nitrogen is the most abundant macro element and was highest in accession A4 followed by accession A1 at 60, 90 and 120 DAS. Accession A5 produced the lowest amount of nitrogen at all growth stages (Fig. 2). Potassium content among hyacinth bean accessions occupied the second position followed by calcium and phosphorus concentrations at 60, 90 and 120 DAS.
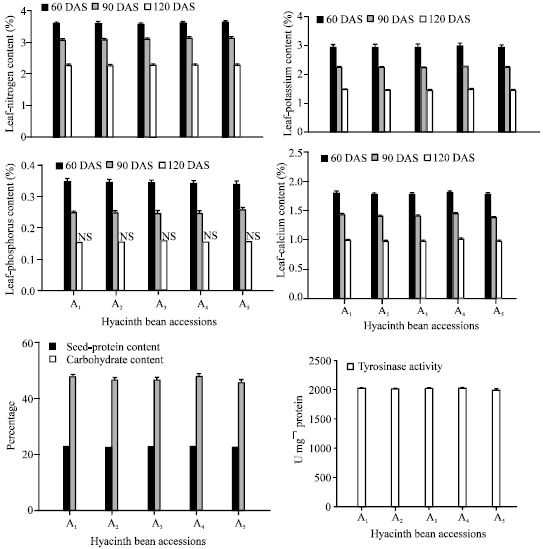 | |
| Fig. 2: | Changes in leaf-nitrogen, phosphorus, potassium and calcium contents (60, 90 and 120 DAS), seed-protein content, carbohydrate content and tyrosinase activity of hyacinth bean (Lablab purpureus L.) accessions analyzed at 150 DAS. Error bars (τ) show LSD at 5% level |
Accession A4 proved superior followed by A1 for P, K and Ca contents at all the growth stages (Fig. 2). All the accessions were not significantly differ from each other for phosphorus content at 120 DAS (Fig. 2). A4 and A1 had the maximum content of protein (25.06 and 25.03%, respectively) and A5 and A2 (24.70 and 24.80%, respectively), followed by A3 (24.86%) had less protein content, which had almost equal value (Fig. 2). On the other hand, A4 surpassed the others for carbohydrate content and had the maximum value (53.16%), whereas, A5 reported the minimum (50.62%). As far as tyrosinase activity is concerned, accession A4 produced the maximum activity for tyrosinase and A5 gave the minimum value (Fig. 2).
DISCUSSION
The present study indicates that differences among hyacinth bean accessions in these agro-botanical attributes exist. Accession A4 proved superior over all other accessions and significant differences (p≤0.05) for most attributes were observed. Variation in biomass production among hyacinth bean accessions due to accessions phenology, environment and season has been reported in various accessions (Holland and Mullen, 1995; Bernas, 1996; Murphy and Colucci, 1999; Pengelly and Maass, 2001; Ewansiha et al., 2007). According to agro-climatic conditions of India, intraspecific variations have been found on various physiological and morphological traits in other medicinal plants (Kulkarni et al., 1984; Singh et al., 1992). Furthermore, it has been reported that varietal differences among the accessions are greater than differences between related species or genera (Millikan, 1961). Significant differences in pod number, 100-seed weight and seed-yield among accessions was probably due to differences in their genetic makeup and the environmental conditions under which the hyacinth bean plants were grown. The higher values for nitrate reductase and carbonic anhydrase activities, net photosynthetic rate, stomatal conductance, nodule-nitrogen content together with appropriate amounts of nitrogen, phosphorus, potassium and calcium were also responsible for better hyacinth bean growth, number and dry weight of nodules. Hence, higher values of fresh and dry weights, number of pods and 100-seed weight of all accessions were recorded. Seed-yield is a cumulative performance of pod number, seed number per pod and 100-seed weight, respectively. Enhancement in yield attributes would ultimately culminate into seed-yield production of each accession. Similar studies regarding accession variation for different attributes in other plants were reported by Virk et al. (1989), Singh et al. (1992), Mishra et al. (2001), Choudhary and Gupta (2002), Khan et al. (2003), Naeem et al. (2006), Idrees et al. (2007) and Morris (2008).
In this study, the number and dry weight of nodules increased up to 90 DAS in all accessions (Table 1), however, the values decreased sharply thereafter. This is due to the fact that the initial competition for photosynthates was confined to roots, nodules and aerial vegetative organs. However, at 90 DAS, flowering and fruit setting provided strong sinks for the utilization of photosynthates. This created a shortage of photosynthates supply to the nodules leading to nodule degeneration as suggested by Samiullah and Khan (2003). The contents of chlorophyll and carotenoid were the optimum at 90 DAS in accession A4. The chlorophyll content declined at later growth stages (Fig. 1). Reduction in total chlorophyll and carotenoid content might be due to the accelerated leaf-senescence as a result of ageing. Nitrate reductase and carbonic anhydrase were highest in accession A4 at 60 DAS; however these activities decreased with increasing hyacinth bean age. Interestingly these activities were slower from the vegetative to flowering stage and more rapid from the flowering to fruiting stage in all accessions studied (Fig. 1).
As far as nitrogen-fixation in hyacinth bean accessions is concerned, accession A4 generated the maximum number and weight of nodules, which requires phosphorus nutrition during the nodulation period. Actually, legumes require high amount of phosphorus for their growth, nodule formation and N2-fixation. Leguminous crops have a high phosphorus utilization rate because of their greater requirement during nodulation (Carling et al., 1978) and N2-fixation (Israel, 1987; Sonoboir and Sarawgi, 2000). The observed enhancement in number and dry weight of nodules due to phosphorus availability in the soil could be due to an additional fixation of molecular nitrogen leading to an improvement in nitrogen metabolism for hyacinth bean. The present study showed that nodule-nitrogen and leghaemoglobin contents declined with the age of the crop. Bacteria in the nodules depend upon the plants for their energy source. Therefore, prior to flowering, nodules can compete as a carbohydrate sink. However, once the plant enters the reproductive phase, seeds act as a stronger sink for carbohydrates than nodules, consequently the latter show a decreased dry nodule weight and lower nitrogen-fixing capacity.
Differences in mineral elements among other leguminous crops have been reported by Deka and Sarkar (1990), El Siddig et al. (2002) and Vadivel and Janardhanan (2002). Considering leaf-N, P, K and Ca contents at 90 and 120 DAS in hyacinth bean accessions, a decrease in the concentration of all these nutrients was noted, with the increase in the age of the hyacinth bean (Fig. 2). Such a decrease in N, P, K and Ca contents of leaf may be due to a continuous utilization of these nutrients by the developing pods (sink) and their translocation from vegetative parts (source). Hyacinth bean seeds have higher values for carbohydrates when compared to groundnut (26.1%) and soybean (20.9%) (Narsinga Rao et al., 1989). This study shows that hyacinth bean has healthy mineral nutrients, seed-protein and carbohydrates for use as a high value grain, nutrient and green manure crop.
CONCLUSION
Hyacinth beans contain numerous and important therapeutic compounds for potential use in modern as well as traditional systems of medicine. This study proves that hyacinth bean seed is an important source of proteins, carbohydrates and minerals such as phosphorus, potassium and calcium. On the basis of present study conducted under the agro-climatic conditions of Aligarh, Western Uttar Pradesh, it is to be concluded that hyacinth bean (accession A4) grew well in calcium deficient soil and can be cultivated in our country. Whereas, accession A5 showed poorest performance under such conditions. Hyacinth bean has the potential to provide a quality and nutritious vegetable to the people of India.
ACKNOWLEDGMENT
The authors are grateful to Dr. Brad Morris, USDA, ARS, Plant Genetic Resources Conservation Unit, Griffin, GA 30223-1797, USA for generous supply of seeds of hyacinth bean accessions.
REFERENCES
- Alhasan, S.A., O. Aranha and F.H. Sarkar, 2001. Genistein elicits pleiotropic molecular effects on head and neck cancer cells. Clin. Cancer Res., 7: 4174-4181.
Direct Link - Bernas, P., 1996. Research note: Dry matter production and chemical composition of introduced forages at two moist savanna sites in Ghana. Trop. Grassland, 30: 418-421.
Direct Link - Coseteng, M.Y. and C.Y. Lee, 1987. Changes in apple polyphenoloxidase and polyphenol concentrations in relation to degree of browning. J. Food Sci., 52: 985-989.
CrossRefDirect Link - Dwivedi, R.S. and N.S. Randhawa, 1974. Evaluation of a rapid test for the hidden hunger of zinc in plants. Plant Soil, 40: 445-451.
CrossRefDirect Link - El Siddig, O.O.A., A.H. El Tinay, A.W.H, Abd Alla and A.E.O. Elkhalifa, 2002. Proximate composition, minerals, tannins, in vitro protein digestibility and effect of cooking on protein fractions of hyacinth bean (Dolichos lablab). J. Food Sci. Technol., 39: 111-115.
Direct Link - Ewansiha, S.U., U.F. Chiezey, S.A. Tarawali and E.N.O. Iwuafor, 2007. Potential of Lablab purpureus accessions for crop-livestock production in the West African savanna. J. Agric. Sci., 145: 229-238.
CrossRef - Fiske, C.H. and Y. Subbarow, 1925. The colorimetric determination of phosphorus. J. Biol. Chem., 66: 375-400.
CrossRefDirect Link - Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedure for Agricultural Research. 2nd Edn., John Wiley & Sons, Hoboken, New Jersey, ISBN: 9780471870920, Pages: 704.
Direct Link - Holland, J.F. and C.L. Mullen, 1995. Register of Australian grain legume cultivars. Lablab purpureus (L.) Sweet (lablab) cv. Koala. Aust. J. Exp. Agric., 35: 559-559.
CrossRefDirect Link - Hoffman, R., 1995. Potent inhibition of breast cancer cell lines by the isoflavonoid Kievitone: Comparison with genistein. Biochem. Biophy. Res. Commun., 211: 600-606.
CrossRefDirect Link - Israel, D.W., 1987. Investigation of the role of phosphorus in symbiotic dinitrogen fixation. Plant Physiol., 84: 835-840.
Direct Link - Jaworski, E.G., 1971. Nitrate reductase assay in intact plant tissues. Biochem. Biophys. Res. Commun., 43: 1274-1279.
CrossRefDirect Link - Karachi, M., 1998. Growth and nutritive value of Lablab purpureus accessions in semi-arid Kenya. Trop. Grassland, 31: 214-218.
Direct Link - Khan, M.M.A., J. Puthukudi, F. Mohammad, M.H. Siddiqui and M. Naeem, 2003. Screening tomato varieties for phytonutrients productivity and yield performance. Muarik Bull., 6: 59-64.
Direct Link - Kobayashi, T., T. Nakata and T. Kuzumaki, 2002. Effect of flavonoids on cell cycle progression in prostate cancer cells. Cancer Lett., 176: 17-23.
Direct Link - Lichtenthaler, H.K. and C. Buschmann, 2001. Extraction of photosynthetic tissues: Chlorophylls and carotenoids. Curr. Protocol. Food Anal. Chem., 1: F4.2.1-F4.2.6.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Maass, B.L., 2006. Changes in seed morphology, dormancy and germination from wild to cultivated hyacinth bean germplasm (Lablab purpureus: Papilionoideae). Genet. Res. Crop Evol., 53: 1127-1135.
CrossRef - Mishra, P., G.C. Uniyal, S. Sharma and S. Kumar, 2001. Pattern of diversity for morphological and alkaloid yield related trades among the periwinkle Catharathus roseus accessions collected from in and around Indian Subcontinent. Genet. Resour. Crop Evol., 48: 273-286.
Direct Link - Morris, J.B., 1997. Special-purpose legume genetic resources conserved for agricultural, industrial and pharmaceutical use. Econ. Bot., 51: 251-263.
CrossRef - Morris, J.B., 2003. Bio-functional legumes with nutraceutical, pharmaceutical and industrial uses. Econ. Bot., 57: 254-261.
Direct Link - Morris, J.B., 2008. Macrotyloma axillare and M. uniflorum: Descriptor analysis, anthocyanin indexes and potential uses. Genetic Resour. Crop Evolu., 55: 5-8.
Direct Link - Murphy, A.M. and P.E. Colucci, 1999. A tropical forage solution to poor quality diets: A review of Lablab purpureus. Livestock Res. Rural Dev. 11(2).
Direct Link - Pengelly, B.C. and B.L. Maass, 2001. Lablab purpureus (L.) sweet-diversity, potential use and determination of a core collection of this multi-purpose tropical legume. Genetic Res. Crop Evolu., 48: 261-272.
CrossRefDirect Link - Paul, B. and L.R. Gowda, 2000. Purification and characterization of a polyphenol oxidase from the seeds of field Bean (Dolichos lablab). J. Agric. Food Chem., 48: 3839-3846.
Direct Link - Samiullah and N.A. Khan, 2003. Physiological investigations on interactive effect of P and K on growth and yield of chickpea. Ind. J. Plant Physiol., 8: 165-170.
Direct Link - Singh, B.M., S.K. Pareek, S. Mandal, M.L. Maheshwari and R. Gupta, 1992. Variability in periwinkle (Catharanthus roseus). Indian J. Agric. Sci., 62: 47-50.
Direct Link - Vadivel, V. and K. Janardhanan, 2002. Agrobotanical traits and chemical composition of Cassia obtusifolia L.: A lesser-known legume of the Western Ghats of South India. Plant Foods Hum. Nutr., 57: 151-164.
CrossRefDirect Link - Watson, D.J., 1958. The dependence of net assimilation rate on leaf-area index. Ann. Bot., 22: 37-54.
Direct Link - Sanchez-Ferrer, A., J.N. Rodriguez-Lopez, F. Garcia-Canovas and F. Garcia-Carmona, 1995. Tyrosinase: A comprehensive review of its mechanism. Biochim. Biophys. Acta, 1247: 1-11.
PubMed