
Mohamed M. Azooz
Department of Biology, Faculty of Science, King Faisal University, Saudi Arabia
Mohamed A Al-Fredan
Department of Biology, Faculty of Science, King Faisal University, Saudi Arabia
American Journal of Plant Physiology
Year: 2009 | Volume: 4 | Issue: 1 | Page No.: 38-51







ABSTRACT
This research was carried out to study the inductive role of vit. C and its mode of application in mitigating the detrimental effects of irrigation with diluted (10, 20 and 30%) seawater on Vicia faba L. cv. Hassawi plants. The results showed that, seawater level at 10% exhibited insignificant changes, while the higher levels impaired growth by reducing seeds germination, dry weights of shoot and root, water status and chlorophyll contents. However, seawater irrigation enhanced carotenoids, antioxidant enzyme activities, MDA and ion leakage. The detrimental effects imposed by seawater were ameliorated by application of 100 ppm vit. C either by seed soaking or shoot spraying. Shoot spraying method gave the best effect. The inductive role of vit. C was associated with improvement of seed germination, growth, plant water status, carotenoids, endogenous ascorbic acid and antioxidant enzyme activities, which consequently led to reduction in ion leakage and MDA level. Moreover, vit. C alone or in combination with seawater increased the intensity of protein bands as well as synthesized additional new proteins with molecular weights of 382, 154, 132, 106 and 67 kDa. This may lead to an increase in the tolerance mechanisms of treated plants towards seawater salinity.
PDF Abstract XML References Citation
How to cite this article
Mohamed M. Azooz and Mohamed A Al-Fredan, 2009. The Inductive Role of Vitamin C and its Mode of Application on Growth, Water Status, Antioxidant Enzyme Activities and Protein Patterns of Vicia faba L. cv. Hassawi Grown under Seawater Irrigation. American Journal of Plant Physiology, 4: 38-51.
DOI: 10.3923/ajpp.2009.38.51
URL: https://scialert.net/abstract/?doi=ajpp.2009.38.51
DOI: 10.3923/ajpp.2009.38.51
URL: https://scialert.net/abstract/?doi=ajpp.2009.38.51
INTRODUCTION
Plants experience many problems living in or near seawater, which is physiologically dry because most plants and tissues are more dilute than seawater. Saudi Arabia is predominantly an arid country, and the shortage of fresh water resources for irrigation; is one of the problems of agriculture and limiting factors for obtaining higher crop yield (Hamdy et al., 2005; Al-Fredan, 2008). The water supplies from the existing surface irrigation systems are not enough to meet crops water requirements. So, there is increasing consciousness among agricultural scientists and planners for using/diluted seawater for irrigation of crops (Di-Baccio et al., 2004; Long et al., 2008). Development of soil salinity has increased significantly in the eastern region of the Kingdom of Saudi Arabia. The high concentrations of salt resulting from brackish groundwater, natural processes or disarrangement in irrigated agriculture result in inhibition of plant growth and yield (Demiral and Turkan, 2005).
Seeds germination, is one of the most critical phases of plant life, is greatly influenced by salinity (Abo-Kassem, 2007), which either induces a state of dormancy at low levels or completely inhibits germination at higher levels (Iqbal et al., 2006). Pahlavani et al. (2006) proved that genetic information regarding seed germination could help to improve seedling emergence in saline soils through breeding programs. Increasing of sodium concentration in plant tissue can result in an increase in oxidative stress, which causes deterioration in chloroplast structure and an associate lose in chlorophyll. This leads to a decrease in chlorophyll, while carotenoid content increased (Khosravinejad and Faboondia, 2008). Further, reactive oxygen species (ROS) like superoxide, hydrogen peroxide and hydroxyl radicals are generated (Wahid et al., 2007). ROS are highly reactive in the absence of any protective mechanism. They can seriously disrupt normal metabolism through oxidative damage to essential membrane lipids, proteins and pigments (Di-Baccio et al., 2004; Cakmak, 2005). To scavenge ROS; Mittler (2002) showed that, plants synthesis different types of defense system composed of non-enzymatic antioxidants, such as ascorbic acid and enzymatic antioxidants like catalase (CAT), peroxidase (POD), ascorbate peroxidase (APX) and glutathione reductase (GR). Scavenging system having potential to quench ROS in stress tolerance plants (Sairam et al., 2005; Koca et al., 2007).
The adaptation to salinity stress is accompanied by alterations in the level of protein patterns. Salinity induces the synthesis of salt stress-specific proteins; some of these proteins were suggested to protect the cell against the adverse effect of salt stress. Vitamins were generally found to affect on gene expression. They induced the synthesis and increased the amount of the original proteins which were already present in the control plants as well as the appearance of additional new bands (Azooz, 2004; Bassuony et al., 2008; Beltagi, 2008). The significant increase in the intensity of the original bands appeared in the control indicate that, vitamins have profound effect on the qualitative and quantitative changes in the protein component of these plants, which might linked with improvement of their growth and productivity.
Vicia faba L. cv. Hassawi is an important legume plant as a major source of protein. Legumes have been suggested as appropriate crops for the enhancement of productivity and reclamation of marginal lands, because these plants not only consider as a good fodder and protein rich seeds, but they also enrich soil nitrogen in symbiotic association with Rhizobium (Hungria and Vargas, 2000).
Vitamin C is a small, water-soluble antioxidant molecule that acts as a primary substrate in the cyclical pathway for detoxification and neutralization of superoxide radicals and singlet oxygen (Noctor and Foyer, 1998). Many studies have been reported that vitamin C when used with optimal concentration exhibited beneficial effect on growth and yield of some crop plants grown under saline conditions (Azooz, 2004; Khan et al., 2006; Bassuony et al., 2008). They reported that, vit. C can play an inductive role in alleviating the adverse effect of salinity on plant growth and metabolism in many plants. So, the main objective of this study was to investigate the inductive role of 100 ppm vit. C solution either before (seed soaking) or after (shoot spraying) cultivation on seed germination, growth, water status, antioxidant enzymes and protein patterns of Vicia faba L. cv. Hassawi under irrigation with diluted seawater.
MATERIALS AND METHODS
This study was conducted within the period of May-October 2008 in King Faisal University, Saudi Arabia.
The experiment was carried out with different levels (0, 10, 20 and 30%) of seawater and 100 ppm vitamin C solution, either by seed soaking, or shoot spraying. This work includes 2 parts; the first part was at seed germination stage, while the second part was at the vegetative growth stage.
The First Part
It was conducted to follow any possible alterations in gene expression in germinated seeds as a result of seeds treatment with seawater alone, vitamin C alone or both in comparing with untreated control seeds (seeds germinated in tap water).
Seed Germination
Homogenous seeds of Vicia faba L. cv. Hassawi were surface-sterilized for 4 min in 2% sodium hypochlorite solution; then rinsed with sterilized water 3 times and air-dried. These seeds were divided into two groups. The first group was soaked in distilled water (control), while the second group was soaked in 100 ppm aqueous solution of vit. C for 8 h and air-dried. Then, the seeds were germinated in sterile petri dishes lined with filter paper and moistured with appropriate amounts of different levels of seawater [0, 10 (10 volumes seawater/90 volumes tap water), 20 and 30%] and incubated at 22°C in dark conditions. Three replicates petri dishes were prepared from each treatment (12 seeds per each petri dish). Seeds were considered to be germinated after the radical emerged from the testa. After germination, seedlings were harvested, frozen in liquid nitrogen and stored under -80°C. The protein was then extracted to analyzed using electrophoretic technique.
Extraction of Protein
The frozen tissue (0.5 g) was homogenized in 2 mL of extraction buffer [50 mM Tris-HCl (pH 6.8), 20 mM dithiothreitol (DTT), 0.5 mM MgCl2, 0.004% EDTA and 10% sucrose]. The homogenate was incubated in water bath at 40°C for 30 min and boiled for 4 min. Thereafter, it was centrifuged at 13.000 g for 1 h at 4°C. The samples were mixed with 0.025% Bromophenol Blue before their loading in the gel electrophoresis.
Electrophoresis
One dimensional Sodium Dodecyl Sulphate Polyacrylamide Gel Electrophoresis (SDS-PAGE) of proteins was applied according to the procedure of Laemmli (1970) with some modifications.
Gel Staining
After electrophoresis, protein bands in the gel were visualized by a Coomassie Brilliant Blue R-250 (CB) with Bismarck Brown R (BBR) according to Choi et al. (1996).
The Second Part
Another experiment was conducted to investigate the effect of 100 ppm vit. C and its mode of application (either by seed soaking or shoot spraying) on growth, water status, photosynthetic pigments, antioxidant enzymes of 30 day-old Vicia faba L. cv. Hassawi plants grown under seawater salinity.
The experimental seeds were grown in plastic pots lined with polyethylene bags and filled with soil composed of clay and sand (1:2 v/v). Seeds were sown in the plastic pots and the seedlings selected on the basis of vigor and uniformity; were subjected to the desired diluted seawater levels (0, 10, 20 and 30%). Five plants were allowed to grow in each pot and three replicates were prepared for each level of seawater.
Two different methods of vit. C application have been used in the present study. Thus, three sets were prepared; the first set (seedlings untreated with vit. C, control), the second set (seedlings having seed treatment with vit. C) and the third set (seedlings receiving foliar application of vit. C).
Plant Growth and Treatments
The all pots of the three sets were kept in growth chamber maintained at 22/20 ±2°C day/night (12 h) temperature cycles and relative humidity 60% until having two foliage leaves. Then, the shoots of the first set (reference control), were sprayed with distilled water, while the plants of the third set were sprayed with aqueous solution of 100 ppm vit. C. Spraying was conducted three times at intervals 7 days (the soil was covered during spraying). Thereafter, the test plants were regularly irrigated every two days with tap water to reach in each treatment to the above desired seawater levels, by weighing the whole system (pot, soil and polyethylene bag) and left to grow for the period of the experiment (30 days).
Harvesting
The plants of all groups were harvested and split up into the root and shoot system. The freshly harvested roots and shoots were rinsed with deionized water and blotted on paper towels before being weighed (fresh weight). To determine the dry weight, roots and shoots were dried in an aerated oven at 70°C until constant weight. Transpiration rate was estimated as described by Bozcuk (1975). Relative Water Content (RWC) of leaves was determined according to Smart (1974). Ion leakage (EC%) of the excised leaves was measured as described by Yan et al. (1996).
Photosynthetic Pigments
Chlorophyll (chl. a and b) and carotenoids contents in leaves were estimated in 80% acetone extracts according to Lichtenthaler and Wellburn (1983).
Assays of Some Antioxidant Enzyme Activities
Enzyme Extraction
The samples were prepared as described by Mukherjee and Choudhuri (1983). A leaf sample (0.5 g) was frozen in liquid nitrogen and finely ground by pestle in a chilled motor, the frozen powder was added to 10 mL of 100 mM phosphate buffer (KH2PO4/K2HPO4) pH 7.0, containing 0.1 mM Na2EDTA and 0.1 g of Polyvinylpyrrolidone, the homogenate was filtered through cheese cloth, then centrifuged at 15.000 x g for 10 min at 4°C. The supernatant was recentrifuged at 18.000 x g for 10 min and the resulted supernatant was collected and stored at 4°C for CAT, POD, APX and GR assays.
Assay of Catalase Activity
Catalase (EC 1. 11. 1. 6) activity was assayed in a reaction solution (3 mL) contained 50 mM phosphate buffer (pH 7.0), 30% (w/v) H2O2 and 0.5 mL of enzyme extract (Aebi, 1984). The reaction was started by the addition of enzyme extract. The activity of catalase was estimated by the decrease of absorbency at 240 nm for 1 min as a consequence of H2O2 consumption (Havir and McHale, 1987).
Assay of Peroxidase Activity
Peroxidase (EC 1. 11. 1. 7) activity was determined according to Maehly and Chance (1954) by the oxidation of guaiacol in the presence of H2O2. The increase in absorbance due to formation of tetraguaiacol was recorded at 470 nm (Klapheck et al., 1990).
Assay of Ascorbate Peroxidase Activity
The activity of ascorbate peroxidase (EC 1. 11. 1 11) was assayed according to Chen and Asada (1992). The reaction was started by addition of H2O2. The activity of enzyme was assayed by measuring the decrease in absorbance at 290 nm for 1 min of ascorbate as ascorbate oxidized.
Determination of Lipid Peroxidation
The level of lipid peroxidation was determined by measuring the amount of Malon Di Aldehyde (MDA) produced by the thiobarbituric acid (TBA) reaction as described by Heath and Packer (1968). The absorbance of supernatant was recorded at 532 and 600 nm. After subtracting the non-specific absorbance at 600 nm, the MDA content was calculated using its molar extinction coefficient (155 mM-1 cm-1) and the results expressed as nmol (MDA) g-1 fresh weight.
Ascorbic Acid Determination
It was determined according to the method of Mukherjee and Choudhuri (1983). The absorbance was read at 525 nm. Its concentration was calculated from a standard curve plotted with known concentrations of ascorbic acid.
Statistical Analysis
All data were analyzed statistically by one-way ANOVA. Values in the figures indicate mean values±SD based on three independent determinations (n = 3) and the Least Significant Difference (LSD) was used to test the difference between treatments; p≤0.05 was considered statistically significant. Statistical analysis were performed using SPSS program.
RESULTS AND DISCUSSION
The germination percentage of Vicia faba L. cv. Hassawi seeds under different levels of seawater irrigation (Fig. 1) was unaffected at 10%. However, a significant decrease at the higher levels was recorded. The maximal germination percentage was 31.7% as compared with control. Seeds soaking in 100 ppm vit. C increased their percentage of germination. It is noticeable that the inhibitory effect imposed by seawater irrigation was completely alleviated at the mild (20%), while at the highest (30%) seawater level; the maximal germination percentage was 83.3%. The inhibitory effect of seawater on seed germination may be partially osmotic due to declining solute potential or ion toxicity due to accumulation of some ions in the seeds, which can alter some physiological processes such as enzyme activation (Croser et al., 2001; Hajer et al., 2006; Jaleel et al., 2007). Abo-Kassem (2007) reported that high salinity delayed radical emergence and decreased germination percentage. The improvement effect of vit.C on germination proved the success of using vit. C as pretreatment of Vicia faba L. seeds to reduce the inhibitory effect of seawater stress on their germination. These positive results of vit. C on seeds germination were reported by Shaddad et al. (1990) and Arab and Ehsanpour (2006). Arrigoni and De Tullio (2000) reported that, exogenous ascorbic acid increased the level of ascorbic acid uptake by different tissues. The additional ascorbic acid (vit. C) is associated with the partial inhibition of ROS production (Shalata and Neumann, 2001). So, it can be concluded that, the inductive role of vit. C in seed germination is attributed to its antioxidant activity.
Changes of protein patterns by one-dimensional SDS-PAGE have analyzed in germinated seeds of Vicia faba L. cv. Hassawi (Fig. 2), to follow any possible alterations in gene expression in these seeds as a result of seeds treatment with 30% seawater alone, vitamin C alone or both in comparing with control (seeds germinated in tap water).
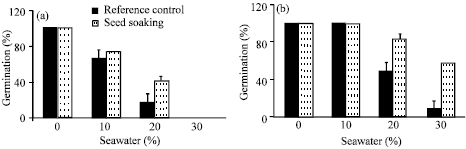 | |
| Fig. 1: | Effect of different seawater levels (%) on percentage germination of Vicia faba L. cv. Hassawi seeds after being soaked for 8 h in 100 ppm vit. C and air dried. (a) 1st, (b) 2nd and (c) 3rd day. Vertical bars represent ±SD |
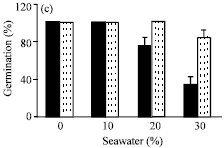
 | |
| Fig. 2: | Analysis of protein patterns by one-dimensional SDS-PAGE extracted from germinated seeds of Vicia faba L. cv. Hassawi in 30% seawater and/or 100 ppm vit. C solution. M: Marker protein (14.2-272 kDa); Lane C : Control (Seeds germinated in tap water only); Lane 1: Seeds soaked in 100 ppm vit. C and germinated in tap water (0.0% seawater); Lane 2: Seeds germinated in 30% seawater only and Lane 3: Seeds soaked in 100 ppm vit. C and germinated in 30% seawater |
Protein bands indicate the presence of about 14 polypeptides with apparent molecular weights ranging from 14-382 kDa. Seeds germinated in 30% seawater (lane 2) showed that seawater salinity enhanced the synthesis of most original proteins which were already present in control seeds (lane C), especially 272, 182, 82 and 26 kDa polypeptides as well as synthesized additional five new proteins with molecular weight of 382, 154, 132, 106 and 67 kDa. Soaking of seeds in vit. C elevated the levels of proteins in most bands of both seeds germinated either in 0 (control) or 30% seawater. Further, in seeds germinated in 0% seawater; vit. C (lane 1) induced synthesis of one new band with molecular weights of 67 kDa. This was accompanied with disappearance of one band (17 kDa). While, in seeds germinated in 30% seawater, vit. C (lane 3) also, resulted in appearance of five new proteins with molecular weight of 382, 154, 132, 106 and 67 kDa. In addition, the protein band (17 kDa) which had disappeared in seeds germinated in 0% seawater and vit. C; reappeared. Similar results were reported by Azooz (2004), Kassim and Dowidar (2006) and Beltagi (2008).
The changes in protein profile may be due to adaptation of Vicia faba L. seeds to seawater stress. The new bands of proteins in seedlings germinated in seawater or in combination with vit. C may be due to de novo synthesis of new protein (Gopala Roa et al., 1987; Azooz, 2004). Bassuony et al. (2008) have shown, vitamin treatments induced a significant alterations in the enzymes related to protein metabolism; which indicate that vitamins might act as activators of protein synthesis. The new bands and the significant increase in the intensity of the original bands appeared in the control indicate that, vit. C has stimulatory effect on the protein component, which might linked with the improvement of seed germination and growth. Therefore, it can be suggested that the new proteins which appeared in seedlings germinated in 30% seawater alone or with vit. C and did not appear in untreated seedlings (control), may play an inductive role in triggering a special system helping Vicia faba L. cv. Hassawi seeds to tolerate seawater stress and increase their capacity to germinate.
Fresh and dry weights of root and shoot (Fig. 3a-d) and water status in terms of Water Content (WC), Relative Water Content (RWC) of leaves and transpiration (Fig. 4a-d) of Vicia faba L. cv., Hassawi plants, exhibited variations as a result of seawater irrigation.
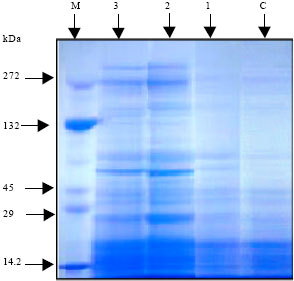
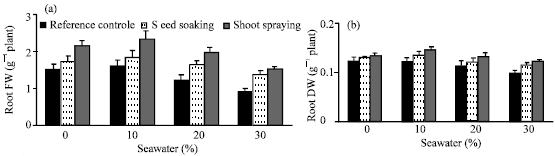 | |
| Fig. 3: | Effect of vit. C (100 ppm) treatments either by seed soaking or shoot spraying on (a) fresh weight and (b) dry weight of root, (c) fresh weight and (d) dry weight of shoot and (e) root/shoot ratio of Vicia faba L. cv. Hassawi plants grown under different levels of seawater. Vertical bars represent ±SD |
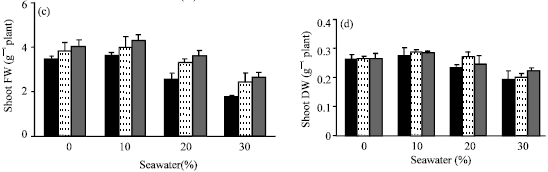
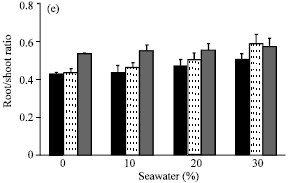
In comparing with control, no significant differences were found in growth parameters (fresh and dry weights of root and shoot) and water status of plants irrigated with 10% seawater. Moreover, stimulation effects on dry weight of shoot (Fig. 3d) and relative water content of leaves (Fig. 4c) were recorded. However, a significant decrease was observed at the higher seawater levels. The growth parameters yields of tested plants appeared to be positively correlated with their WC and RWC. Seawater salinity caused more inhibition in roots growth than in shoots. So, root/shoot ratios (on the basis of fresh weight) were increased (Fig. 3e) with increased of seawater level. Kaya et al. (2003) reported that, root growth was more sensitive and adversely affected as compared to shoot growth under salinity conditions. Reduction in plant growth as a result of seawater stress has been reported earlier in several plants (Hajer et al., 2006; Alqurainy, 2007; Long et al., 2008). Increasing of seawater level reduced the absorption of water leading to a drop in water content of tested plants. Thus, the inhibitory effect of seawater on growth parameters could be attributed to the osmotic effect of seawater salinity (Salter et al., 2007). In addition, the changes in water status under seawater stress may cause a reduction in meristem activity as well as cell elongation (Shah, 2007).
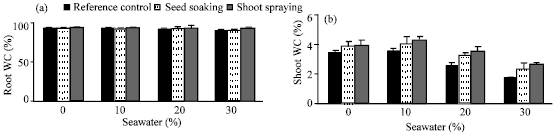 | |
| Fig. 4: | Effect of vit. C (100 ppm) treatments either by seed soaking or shoot spraying on water content % of (a) root and (b) shoot, (c) leaf relative water content % and (d) transpiration rate of Vicia faba L. cv. Hassawi plants grown under different levels of seawater. Vertical bars represent ±SD |
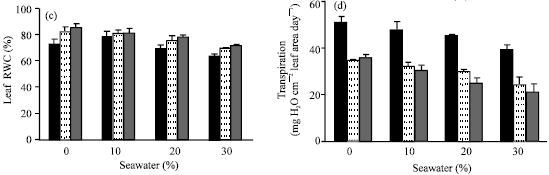
The adverse effects of seawater salinity on the growth parameters, WC and RWC were mitigated by seed soaking or shoot spraying with 100 ppm vit. C. These results are in coincidence with that cited by Azooz (2004), Alqurainy (2007) and Athar et al. (2008). They suggested that, vit. C could be accelerated cell division and cell enlargement of treated plants. Shoot spraying with vit. C was more effective in improving of growth parameters of treated plants which was associated with increasing of their WC, RWC of leaves and reduction in transpiration rate. This indicates that shoot spraying probably reflects the efficiency of water uptake and utilization or reduces water loss which consequently causes a concomitant increase in leaf water potential. Hence, it can be concluded that the beneficial effect of vit. C on growth parameters of Vicia faba L. cv. Hassawi has been related to the efficiency of their water uptake and utilization. These suggestions are in a good agreement with present results, which cleared that the increase of WC and RWC was associated with a decrease in transpiration rate. Further, it could be suggested that, the effectiveness of vit. C depends on its mode of application, which may enhance the endogenous level of vit.C and water status of treated plants.
Photosynthetic pigments of Vicia faba L. cv. Hassawi leaves (Fig. 5a-d) were substantially affected under seawater irrigation. The content of chl.a and chl.b was more or less unchanged under 10% seawater level. While, at higher levels of seawater; a significant decrease was observed. On the other side, the content of carotenoid was increased at low and moderate seawater levels as compared with control. The reduction in chl.b was higher (about 44%) than chl.a, (about 30%) below the control at the highest seawater level, resulting in a higher chl. a/chl. b ratio (Fig. 5d). The inhibitory effect of seawater stress on photosynthetic pigments was completely alleviated as result of vit. C treatments. Moreover, the values of pigments were higher than those of control plants at most seawater levels used. These results reinforce the results obtained by Shah (2007) and Beltagi (2008). The reduction observed in chlorophyll content under seawater irrigation could be as a result of inhibition of chlorophyll biosynthesis or increased of its degradation (Khan et al., 2006). Further, under seawater stress, an oxidative stress could result, which causes deterioration in chloroplast structure. This leads to a decrease in chlorophyll content, while carotenoid content increased (Khosravinejad and Farboondia, 2008). Carotenoids are known to act as efficient quenchers of free radical caused by ROS.
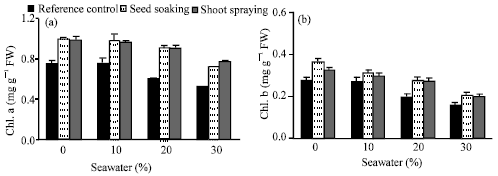 | |
| Fig. 5: | Effect of vit. C (100 ppm) treatments either by seed soaking or shoot spraying on (a) chl.a, (b) chl.b, (c) carotenoids and (d) chl.a/chl.b ratio of Vicia faba L. cv. Hassawi plants grown under different levels of seawater. Vertical bars represent ±SD |
Thus, increasing of carotenoids in plants treated with seawater and/or vit. C could be enhanced the capacity of these plants to minimize the damage caused by ROS. Therefore, chlorophyll content of plants treated with vit. C was increased, that could result from the protection effect of vit. C and carotenoids to the photosynthetic apparatus from seawater induced oxidative stress (Khan et al., 2006).
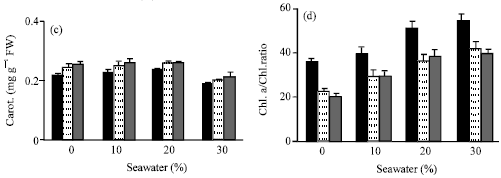
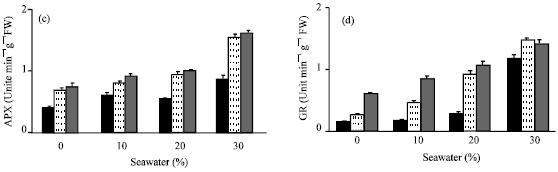
Seawater irrigation induced variations in the activities of antioxidant enzymes (Fig. 6a-d). The level of antioxidant enzyme activities (CAT, POD, APX and GR) was increased. This increase was proportional to seawater concentration. The activity of APX and GR (Fig. 6c and d) was higher than CAT and POD ((Fig. 6a and b). Similar results were recorded by Sairam et al. (2005), Mandhania et al. (2006) and Athar et al. (2008). The increases in the activities of antioxidant enzymes may be attributed to the adaptive defense of tested plants against the toxic effect of salinity imposed by seawater irrigation (Bor et al., 2003; Long et al., 2008). They suggested that an accumulation of ROS might have occurred in response to seawater salinity. The oxidative stress generated by ROS was inhibited by increasing in the activities of CAT, POD, APX and GR. Application of vit. C either by seed soaking or shoot spraying caused a further increase in the activities of these antioxidant enzymes. Shoot spraying was more effective than seeds soaking. The higher activity of APX and GR as a result of shoot spraying than in seed soaking was confirmed to explain the higher growth parameters recorded in case of shoot spraying than seed soaking. The role of APX and GR in the H2O2 scavenging in plant cells has been well established in ascorbate-glutathione cycle. This cycle has been implicated in mitigating the effect of ROS (Molina et al., 2002). Vit. C acts as a substrate for APX to scavenge ROS produced in the thaylakoid membrane (Davey et al., 2000). Present results indicated that, such an increase in the protective system of the antioxidant enzyme activities was insufficient to protect plants against damage caused by seawater salinity. Vit. C treatments increased the antioxidant enzyme activities that may be sufficient to protect plants against this damage, which consequently reflect the positive effects of vitamin C on growth, chlorophyll content and water status of treated plants.
MDA is one of the end products which are produced as a result of lipid peroxidation damage caused by free radicals produced from the oxidative stress. Ion leakage is a reflection of seawater stress-induced damage to cellular membranes (Jain et al., 2001).
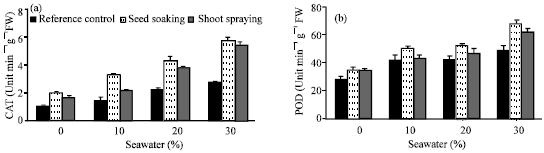 | |
| Fig. 6: | Effect of vit. C (100 ppm) treatments either by seed soaking or shoot spraying on (a) catalase (CAT), (b) peroxidase (POD), (c) ascorbic peroxidase (APX) and (d) glutathione reductase (GR) of Vicia faba L. cv. Hassawi plants grown under different levels of seawater. Vertical bars represent ±SD |
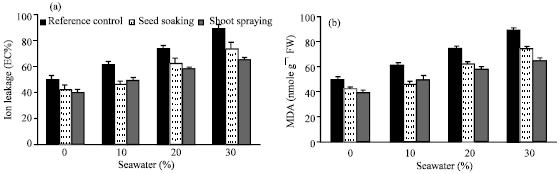 | |
| Fig. 7: | Effect of vit. C (100 ppm) treatments either by seed soaking or shoot spraying on (a) ion leakage (EC%) and (b) lipid peroxidation (MDA) of Vicia faba L. cv. Hassawi plants grown under different levels of seawater. Each value is the mean of three replicates (n = 3). Vertical bars represent ±SD |
Therefore, ion leakage and MDA concentration have been widely used as selection criterion to assess salt injury in various plants. The results obtained (Fig. 7a and b) cleared that, there were no substantial changes in ion leakage (EC%) or MDA content of plants irrigated with 10% seawater. However, they were significantly increased at the higher levels of seawater, in comparing with control. The increase in ion leakage was parallel with the increase of MDA produced. This indicates that, the increase in the ion leakage and the level of MDA in the present study could be due to plasmalemma injury caused by ROS (Yin et al., 2008). These results confirmed the results obtained by Mandhania et al. (2006) and Athar et al. (2008). Seed soaking or shoot spraying with vit. C reduced the ion leakage and MDA contents of treated plants. Vit.C induced improvement of membrane integrity which may have contributed to limit the ion leakage.
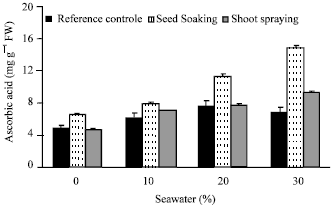 | |
| Fig. 8: | Effect of vit. C (100 ppm) treatments either by seed soaking or shoot spraying on the endogenous ascorbic acid of Vicia faba L. cv. Hassawi plants grown under different levels of seawater. Vertical bars represent ±SD |
The content of endogenous ascorbic acid was increased as a result of seawater and/or exogenous ascorbic acid (vit. C) treatment (Fig. 8). This agrees with Arrigoni and De Tullio (2000) and Athar et al. (2008); they found that, the exogenous application of ascorbic acid increased the endogenous ascorbic acid. They concluded that, ascorbic acid plays an important role as an antioxidant and protects the plant during oxidative damage by scavenging free radicals and active oxygen that are generated during salt stress conditions.
Finally, it could be concluded that, our results could be explained the inductive role played by vit. C in overcoming the detrimental effects of seawater and enhancing the capacity of treated plants to scavenge the free radicals produced as a result of seawater stress. This was associated by improvement of plant growth, water status, carotenoids, endogenous vit. C and antioxidant enzymes activities especially APX and GR. Further, vit. C increases protein synthesis in germinated seeds including de novo synthesis of new proteins and accumulation of certain existing proteins. These indicate that, plants treatment with vit. C trigger some unknown physiological processes which subsequently lead to improvement of seed germination, growth and development of treated plants.
ACKNOWLEDGMENT
The researchers are grateful to the Deanship of the Scientific Research, King Faisal University, Saudi Arabia for funding this study.
REFERENCES
- Abo-Kassem, E.E.M., 2007. Effects of salinity: Calcium interaction on growth and nucleic acid metabolism in five species of Chenopodiaceae. Turk. J. Bot., 31: 125-134.
Direct Link - Aebi, H., 1984. Catalase in vitro. In: Methods in Enzymology, Packer, L., Academic Press, Cambridge, Massachusetts, United States, ISBN: 9780121820053, pp: 121-126.
CrossRefDirect Link - Al-Fredan, M.A., 2008. Effect of treated municipal waste water and Rhizobia strains on growth and nodulation of faba bean (Vicia faba L. cv. Hassawi). Pak. J. Biol. Sci., 9: 1960-1964.
CrossRefDirect Link - Arab, L. and A.A. Ehsanpour, 2006. The effects of ascorbic acid on salt induced alfalfa (Medicago sativa L.) in vitro culture. Biokemistri, 18: 63-69.
Direct Link - Arrigoni, O. and M.C. De Tullio, 2000. The role of ascorbic acid in cell metabolism: Between gene-directed functions and unpredictable chemical reactions. J. Plant Physiol., 157: 481-488.
CrossRefDirect Link - Athar, H.U.R., A. Khan and M. Ashraf, 2008. Exogenously applied ascorbic acid alleviates Salt-induced oxidative stress in wheat. Environ. Exp. Bot., 63: 224-231.
CrossRefDirect Link - Azooz, M.M., 2004. Proteins, sugars and ion leakage as a selection criterion for the salt tolerance of three sorghum cultivars at seedling stage grown under NaCl and nicotinamide. Int. J. Agric. Biol., 6: 27-35.
Direct Link - Bassuony, F.M., R.A. Hassanein, D.M. Baraka and R.R. Khalil, 2008. Physiological effects of nicotinamide and ascorbic acid on Zea mays plant grown under salinity stress. II. Changes in nitrogen constituents, protein profiles, protease enzyme and certain inorganic cations. Aust. J. Applied Sci., 2: 350-359.
- Beltagi, S.B., 2008. Exogenous ascorbic acid (vitamin C) induced anabolic changes for salt tolerance in chick pea (Cicer arietinum L.) plants. Afr. J. Plant Sci., 2: 118-123.
Direct Link - Bor, M., F. Ozdemir and I. Turkan, 2003. The effect of salt stress on lipid peroxidation and antioxidants in leaves of sugar beet Beta vulgaris L. and wild beet Beta maritima L. Plant Sci., 164: 77-84.
CrossRefDirect Link - Cakmak, I., 2005. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. J. Plant Nutr. Soil Sci., 168: 521-530.
CrossRef - Chen, G.X. and K. Asada, 1992. Inactivation of ascorbate peroxidase by thiols requires hydrogen peroxide. Plant Cell Physiol., 33: 117-123.
CrossRefDirect Link - Choi, J., S. Yoon, H. Hong, D. Choi and G. Yoo, 1996. A modified Coomassie Blue staining of proteins in polyacrylamide gels with Bismarck Brown R. Anal. Biochem., 236: 82-84.
CrossRef - Croser, C., S. Renault, J. Franklin and J. Zwiazk, 2001. The effect of salinity on the emergence and seedling growth of Picea mariana, Picea glauca and Pinus banksiana. Environ. Pollut., 115: 9-16.
CrossRefDirect Link - Davey, M.W., M. Van Montagu, D. Inze, M. Sanmartin and A. Kanellis et al., 2000. Plant L-ascorbic acid: Chemistry, function, metabolism, bioavailability and effects of processing. J. Sci. Food Agric., 80: 825-860.
CrossRefDirect Link - Demiral, T. and I. Turkan, 2005. Comparative lipid peroxidation, antioxidant defense systems and proline content in roots of two rice cultivars differing in salt tolerance. Environ. Exp. Bot., 53: 247-257.
CrossRefDirect Link - Di Baccio, D., F. Navari-Izzo and R. Izzo, 2004. Seawater irrigation: Antioxidant defence responses in leaves and roots of a sunflower (Helianthus annuus L.) ecotype. J. Plant Physiol., 161: 1359-1366.
CrossRefPubMedDirect Link - Gopala Rao, P., C.D. Reddy and J.K. Ramaiah, 1987. Effect of B-vitamins on the protein component of clusterbeans Cyamopsis tetragonoloba L. Taub. Ann. Bot., 59: 281-284.
Direct Link - Hajer, A.S., A.A. Malibari, H.S. Al-Zahrani and O.A. Almaghrabi, 2006. Responses of three tomato cultivars to sea water salinity 1. Effect of salinity on the seedling growth. Afr. J. Biotechnol., 5: 855-861.
Direct Link - Hamdy, A., V. Sardob and K.A. Farrag Ghnnem, 2005. Saline water in supplemental irrigation of wheat and barley under rainfed agriculture. Agric. Water Manage., 78: 122-127.
CrossRef - Havir, E.A. and N.A. McHale, 1987. Biochemical and developmental characterization of multiple form of catalase in tobacco leaves. Plant Physiol., 84: 450-455.
PubMed - Heath, R.L. and L. Packer, 1968. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys., 125: 189-198.
CrossRefPubMedDirect Link - Hungria, M. and M.A.T. Vargas, 2000. Environmental factors affecting N2 fixation in grain legumes in the tropics, with an emphasis on Brazil. Field Crop Res., 65: 151-164.
CrossRefDirect Link - Iqbal, M., M. Ashraf, A. Jamil and S. Ur-Rehman, 2006. Does seed priming induce changes in the levels of some endogenous plant hormones in hexaploid wheat plants under salt stress? J. Integr. Plant Biol., 48: 181-189.
CrossRefDirect Link - Jain, M., G. Mathur, S. Koul and N.B. Sarin, 2001. Ameliorating effects of proline on salt stress lipid peroxidation in cell lines of groundnut (Arachis hypogea L.). Plant Cell Rep., 20: 463-468.
CrossRef - Jaleel, C.A., R. Gopi, B. Sankar, P. Manivannan, A. Kishorekumar, R. Sridharan and R. Panneerselvam, 2007. Studies on germination, seedling vigour, lipid peroxidation and proline metabolism in Catharanthus roseus seedlings under salt stress. South Afr. J. Bot., 73: 190-195.
CrossRefDirect Link - Kassim, W.A. and S. Dowidar, 2006. Amino acids and soluble protein profile of radish seedlings under salt stress as affected by GA3 priming. Indian J. Plant Physiol., 11: 75-82.
Direct Link - Khan, M.A., M.Z. Ahmed and A. Hameed, 2006. Effect of sea salt and L-ascorbic acid on the seed germination of halophytes. J. Aird Environ., 67: 535-540.
CrossRef - Kaya, M.D., A. Ipek and A. Ozturk, 2003. Effects of different soil salinity levels on germination and seedling growth of safflower (Carthamus tinctorius L.) Turk. J. Agric. For., 27: 221-227.
Direct Link - Khosravinejad, F., R. Heydari and T. Farboodnia, 2008. Effects of salinity on photosynthetic pigments, respiration and water content in two barley varieties. Pak. J. Biol. Sci., 11: 2438-2442.
PubMedDirect Link - Klapheck, S., I. Zimmer and H. Cosse, 1990. Scavenging of hydrogen peroxide in the endosperm of Ricinus communis by ascorbate peroxidase. Plant Cell Physiol., 31: 1005-1013.
CrossRefDirect Link - Koca, H., M. Bor, F. Ozdemir and I. Turkan, 2007. The effect of salt stress on lipid peroxidation, antioxidative enzymes and proline content of sesame cultivars. Environ. Exp. Bot., 60: 344-351.
CrossRefDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Lichtenthaler, H.K. and A.R. Wellburn, 1983. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans., 11: 591-592.
CrossRefDirect Link - Long, X.H., S.K. Mehta and Z.P. Liu, 2008. Effect of NO-3-N enrichment on seawater stress tolerance of Jerusalem artichoke (Helianthus tuberosus). Pedosphere, 19: 113-123.
Direct Link - Maehly, A.C. and B. Chance, 1954. The assay of catalases and peroxidases. Meth. Anal. Biochem., 1: 357-424.
PubMedDirect Link - Mandhania, S., S. Madan and V. Sawhney, 2006. Antioxidant defence mechanism under salt stress in wheat seedling. Biol. Plant., 50: 227-231.
CrossRefDirect Link - Mittler, R., 2002. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci., 7: 405-410.
CrossRefPubMedDirect Link - Molina, A., P., Bueno, M.C. Marin, M.P. Rodriguez-Rosales, A. Belver, K. Venema and J.P. Donaire, 2002. Involvement of endogenous salicylic acid content, lipoxygenase and antioxidant enzyme activities in the response of tomato cell suspension cultures to NaCl. New Phytol., 156: 409-415.
CrossRefDirect Link - Mukherjee, S.P. and M.A. Choudhuri, 1983. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Plant Physiol., 58: 166-170.
CrossRefDirect Link - Noctor, G. and C.H. Foyer, 1998. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Mol. Biol., 49: 249-279.
CrossRefPubMedDirect Link - Pahlavani Saeidi, M.H.G. and A.F. Mirlohi, 2006. Estimates of genetic parameters for seed germination of safflower in different salinity levels. Asian J. Plant Sci., 5: 133-138.
CrossRefDirect Link - Sairam, R.K., G.C. Srivastava, S. Agarwal and R.C. Meena, 2005. Differences in antioxidant activity in response to salinity stress in tolerant and susceptible wheat genotypes. Biol. Plant, 49: 85-91.
CrossRefDirect Link - Salter, J., K. Morris, P.C.E. Bailey and P.I. Boon, 2007. Interactive effects of salinity and water depth on the growth of Melaleuca ericifolia Sm. (Swamp paperbark) seedlings. Aquatic Bot., 86: 213-222.
CrossRef - Shaddad, M.A., A.F. Radi, A.M. Abdel-Rahman and M.M. Azooz, 1990. Response of seeds of Lupinus termis and Vicia faba to the interactive effect of salinity and ascorbic acid or pyridoxine. Plant Soil, 122: 177-183.
CrossRef - Shah, S.H., 2007. Effects of salt stress on mustard as affected by gibberellic acid application. Genet. Applied Plant Physiol., 33: 97-106.
Direct Link - Shalata, A. and P.M. Neumann, 2001. Exogenous ascorbic acid (vitamin C) increases resistance to salt stress and reduces lipid peroxidation. J. Exp. Bot., 52: 2207-2211.
CrossRefDirect Link - Wahid, A., M. Perveen, S. Gelani and S.M.A. Basra, 2007. Pretreatment of seed with H2O2 improves salt tolerance of wheat seedlings by alleviation of oxidative damage and expression of stress proteins. J. Plant Physiol., 164: 283-294.
CrossRef - Yan, B., Q. Dai, X. Liu, S. Huang and Z. Wang, 1996. Flooding-induced membrane damage, lipid oxidation and activated oxygen generation in corn leaves. Plant Soil, 179: 261-268.
CrossRef - Smart, R.E. and G.E. Bingham, 1974. Rapid estimation of relative water content. Plant Physiol., 53: 258-260.
PubMedDirect Link - Yin, H., Q. Chen and M. Yi, 2008. Effect of short-term heat stress on oxidative damage and responses of antioxidant system in Lilium longiflorum. Plant Growth Regul., 54: 45-54.
CrossRef