
A. Akhter
Laboratory of Plant Nutrition and Soil Science, Faculty of Agriculture, Yamagata University, 1-23 Wakaba Machi, Tsuruoka, Yamagata 997 8555, Japan
T. Wagatsuma
Laboratory of Plant Nutrition and Soil Science, Faculty of Agriculture, Yamagata University, 1-23 Wakaba Machi, Tsuruoka, Yamagata 997 8555, Japan
M.S.H. Khan
Laboratory of Plant Nutrition and Soil Science, Faculty of Agriculture, Yamagata University, 1-23 Wakaba Machi, Tsuruoka, Yamagata 997 8555, Japan
K. Tawaraya
Laboratory of Plant Nutrition and Soil Science, Faculty of Agriculture, Yamagata University, 1-23 Wakaba Machi, Tsuruoka, Yamagata 997 8555, Japan
American Journal of Plant Physiology
Year: 2009 | Volume: 4 | Issue: 1 | Page No.: 1-8







ABSTRACT
Toxic symptoms of crop plant species to Al are widely studied. The most dramatic symptom of Al toxicity is the inhibition of root elongation. As a matter, this symptom long been used to know the Al-tolerance of plant species. But, there was some scope to search proper concentration of Al during the study of Al-tolerance screening. Objective of the present study was to recommend proper concentration of Al for maize (Al-tolerant), soybean (intermediate Al-tolerant) and sorghum (Al-sensitive). Two concentrations of Al (2.5 and 2 0 μM AlCl3 in 0.2 mM CaCl2) were used. For maize former concentration (2.5 μM) was too light to inhibit root elongation (Al tolerance 60-77%) and could not discriminate Al-tolerance among the cultivar whereas later concentration 20 μM represents better illustration of tolerances (40-65%). Soybean was intermediate Al-tolerant crop and both concentration of Al could be suggested for Al-tolerance screening of this crop (40 to 79% for low Al and 17 to 65% for high Al). Sorghum was Al sensitive and high Al concentration made so severe inhibition of root elongation to all cultivars that Al-tolerance among the cultivars could not be differentiate well (Al tolerance 20-24%). On the other hand, at low Al concentration Al-tolerance was in the range of 38-51%. All crop species showed significant positive correlation (R2 = 0.945*, 0.936** or 0.921** for sorghum, soybean and maize, respectively) between the Al tolerance from 2.5 and from 20 μM Al. From these results it could be suggested that treatment concentration for Al tolerance screening should be conducted based on the Al tolerance level of each crop species.
PDF Abstract XML References Citation
How to cite this article
A. Akhter, T. Wagatsuma, M.S.H. Khan and K. Tawaraya, 2009. Comparative Studies on Aluminum Tolerance Screening Techniques for Sorghum, Soybean and Maize in Simple Solution Culture. American Journal of Plant Physiology, 4: 1-8.
DOI: 10.3923/ajpp.2009.1.8
URL: https://scialert.net/abstract/?doi=ajpp.2009.1.8
DOI: 10.3923/ajpp.2009.1.8
URL: https://scialert.net/abstract/?doi=ajpp.2009.1.8
INTRODUCTION
Roots injured by high aluminum are become stubby and thick, dark colored, brittle, poorly branched and rubberized with a reduced root length and volume (Nguyen et al., 2001). Shoot is also inhibited due to limiting supply of water and nutrients. Al toxicity caused Ca deficiency or reduced Ca transport within the plant by curling or rolling of young leaves, inhibited growth of lateral branches or a collapse of growing points or petioles. Young seedlings are affected more than older plants (Thaworuwong and van Diest, 1974). The retardation of plant growth in acid soil occurs not only by toxic elements but also by low pH. Moreover, low availability of nutrients such as Ca, Mg, K and Mo are reported by several researchers in the naturally occurring acid soils. Visible symptoms of Al toxicity include inhibition of root growth (Delhaize and Ryan, 1995), swelling of the root tip and/or sloughing off the epidermis, plasma membrane depolarization, alteration of Ca2+ fluxes at the root-tip, stimulation of callose deposition (Schreiner et al., 1994; Zhang et al., 1994) and induction of rigor in the actin cytoskeleton (Grabski and Schindler, 1995). During the last decade considerable advances have been made in both techniques to assess the potentially toxic Al species in environmental samples and knowledge about the mechanisms of Al toxicity and resistance in plants (Poschenrieder et al., 2008) but still remains unclear.
The major symptom of Al toxicity is a rapid inhibition of root growth (Zhang et al., 2007). Al inhibits root cell expansion and elongation and if over the long term, cell division as well. Al can inhibit cytoskeletal dynamics and interacts with both microtubules and actin filaments (Sivaguru et al., 1999, 2003). This growth inhibition of root further cause reduced plant vigor and yield (Rengel, 1992; Kochian et al., 2005). Toxicity symptoms of Al are similar to nutrient deficiencies (Taylor, 1988) though these general symptoms appear to be the consequence of inhibition of root development caused by targeted action of Al at root tips (Ryan et al., 1993). In many reports, Al tolerance screening of several crops species was done using same Al concentrations. For example, Wagatsuma et al. (2005) screened 18 crop plant species, cultivars and lines using same Al concentrations (20 μM AlCl3 in 0.2 mM CaCl2). On the other hand, several researchers used different concentration of Al for screening. For example, Ishikawa and Wagatsuma (1998) studied Al tolerance with 5 μM Al for barley, maize, pea and rice cultivars. Ma et al. (2005) screened Al tolerance of rice mutants 10 or 50 μM Al. Therefore, objective of the present study was to know whether unique concentration of Al enough for screening of several crop plant differing Al tolerance as species.
MATERIALS AND METHODS
Seeds of maize (Zea mays L.) cultivars were as follows: Royaldent-130, Golddent KD750, Golddent KD417, Royaldent TH470, Goldent KD640, Golddent KD772, Golddent KD720 and Royaldent TH058. Seeds of soybean (Glycine max L.) were as follows: Enrei, Kinshu, Bonodori, Fukura-1, Okuharawase, Ejonishiki, Shinsetsu, Etigohani, Tanba, Miyagishirome, Ryokkou, Miyoshiniki, Fukura-2, Yuagarimusume, Green-75, Sapporomidori, Hiden, Hokkaishirage, Ichiriki, Hisui, Bonnishiki and Ryokuheki. Seeds of sorghum (Sorghum bicolor Moench (L.)) were as follows: Furitsu, Big sugar, Genki, Sudax-316, Gorudo, Kousyouka, Haiguren, Sanzyaku, Syito, Sudax GM, Mairo, Ryokuniyousorgu, Brown Toumitsu and Toumitu A. All the seeds were collected from the Kanto Seed Co. Ltd., Japan and Takii Seed Co. Ltd., Japan. Seeds of maize and sorghum germinated on a nylon screen floating on tap water with aeration. Tap water contains (mg L-1) 8.0 Ca, 2.92 mg, 1.95 K and minor quantity of other minerals (P, Fe, Mn, Zn and Cu). Room temperature was maintained at 27 °C with 0.6 Cd m-2 light intensity.
Seeds of sorghum and maize were soaked in tap water for 24 h. After washing with tap water, seeds were spread on nylon screen floating on 9 L of tap water for germination. When the roots of the seedlings were about 4 cm, seedlings were pretreated with 0.2 mM CaCl2 solution for 6 h. pH of the pretreatment solution corresponds to the pH of the later stage of Al treatment solution. Before starting the actual treatment, root length of each seedling was measured by a ruler. Then the seedlings were subjected (Al treatment) or not-subjected (control) in 9 L of solution containing 2.5 μM AlCl3 (pH 5.0) or 20 μM AlCl3 (pH 4.9) for 24 h. As the availability of most toxic form of Al (Al3+) depends on the solution pH (Koyama et al., 1990), the pH of the culture medium was adjusted based on the added Al on the solution. Root lengths were measured once more just after finishing the treatments.
Seeds of soybean were surface sterilized with 0.5% sodium hypochlorite and spread on previously soaked (with 0.5 M CaSO4) filter paper under dark. Before germination, CaSO4 solution was sprayed once a day. Seedlings were transferred on a nylon screen on a container filled with 9 L of tap water. After growing to proper root length (approx. 4 cm), pretreatment and treatment was carried out in the same way as for sorghum and maize.
To determine the viability or normal growth of the soybean seedlings, the roots were treated with 0.2 mM CaCl2 for 24 h at pH 4.9. Then the roots were stained with fluorescein diacetate-propidium iodide (FDA-PI) (12.5 mg L-1 FDA, 5 mg L-1 PI) for 15 min following the procedure of Khan et al. (2008). The root tips were observed by fluorescent microscope (SMZ-10, Nikon, Japan) equipped with ultraviolet light (Nikon, Japan) (ex. 390 nm, ba 520 nm).
RESULTS AND DISCUSSION
Roots of maize were inhibited by the treatment of 2.5 and 20 μM AlCl3 (Fig. 1). Al tolerance was highest in Royaldent TH085 in both treatment conditions. On the other hand, Al tolerance was least in Royaldent 130 (40%) and Golddent KD750 (42%) in 20 μM Al. Except for these two cultivars, all others (Golddent KD417, Royaldent TH470, Golddent KD640, Golddent KD772, Golddent KD720 and Royaldent TH058) were in the same tolerant group based on treatment with 20 μM Al (data ranged from 54-65%). Highest Al tolerance was observed in Royaldent TH085 in both treatment conditions. Though there were some variations in the Al tolerances at 2.5 μM but these did not differ significantly among the cultivars (Fig. 1).
In low Al concentration (2.5 μM Al), proper discrimination of the maize cultivars can not be done (Fig. 1). The reason might be that this low Al concentration can not make enough stress condition as average Al tolerance of maize is high. In this study it can be found that high Al (20 μM Al) make greater inhibition than low Al in all maize cultivars. However, better discrimination could be observed in high Al treatment conditions. Wagatsuma et al. (2005) used this high Al concentration (20 μM AlCl3) deliberately for screening crops, cultivars or lines having wide variation.
Response of soybean cultivars to both Al treatment (low and high Al) conditions were almost similar though greater variation were observed with 20 μM Al (Fig. 2). However, tendency of Al tolerance were in the order of Ryokoheki, Bonnishiki>Hisui, Ichiriki>Hokkaishirage, Hiden, Sapporomidori, Green-75, Yuagarimusume, Fukura-2>Miyagishirome, Tanba, Etigohani, Shinsetsu, Ejonishiki, Okuharawase, Fukura-1>Bonodori, Kinshu, Enrei.
However, wider range of Al tolerance was found in soybean for both high and low Al conditions which led us to suggest that both kind of Al tolerance screening can be recommended for these crop species (Fig. 2).
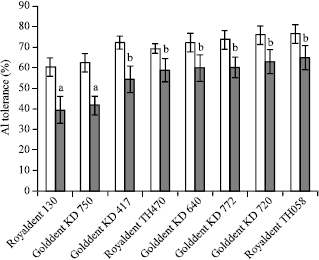 | |
| Fig. 1: | Al tolerance of maize cultivars for 24 h in 2.5 μM ( |
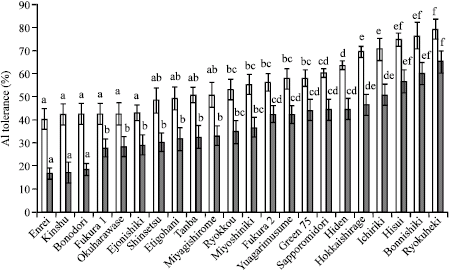 | |
| Fig. 2: | Al tolerance of soybean cultivars for 24 h in 2.5 μM ( |
Presence of Ca in the external solution is essential to maintain intact PM which permits selective ion uptake and prevents the solute leakage from the cytoplasm. The calcium requirement for optimum growth in dicotyledons is higher than monocotyledons (Loneragan et al., 1968, 1969). For this reason, earlier checking of soybean roots was needed whether these roots are getting permeabilized with the same Ca concentration with maize or sorghum. Among the three crops, sorghum and maize were monocotyledonous, however, soybean is dicotyledonous crop and therefore, the Ca requirement of soybean is higher than sorghum and maize. In the present experiment, considering only the Ca source in medium, 72 mg of Ca was equally supplied, i.e., 7.2 mg of Ca was supplied to each seedling in 10-seedling treatments, this is considered as the lowest Ca amount which can grow almost intact seedling without Ca deficiency. In 20-seedlings treatment, Ca allocation from medium is calculated as 3.6 mg which is far lower than the limitation to induce Ca deficiency and therefore the red fluorescence observed under UV light in root-tip portion is ascribed to Ca deficiency. After treating 2, 6 or 8 seedlings in each 9 L of 0.2 mM CaCl2, root-tips emitted green fluorescence under UV light (Fig. 3). Although root-tips of the seedlings treated as 10-seedling treatment emitted whitish green fluorescence, it can be considered as almost intact PM. On the other hand, root-tips emitted yellowish fluorescence when 20-seedlings were treated in the same 9 L of 0.2 mM CaCl2 indicating that the root-tip cells became partially permeabilized. Approximately 0.2 mm-apical portions of roots emit reddish fluorescence in 6, 8 or 10-seedling treatments; however, this may be ascribed to early drying while taking picture. The red fluorescence of these three treatments (6, 8, 10 seedling) is clearly different from that of the 20-seedling treatment condition. In 20-seedlings treatment, not only the root-tip but also the proximal parts emit yellowish fluorescence.
Among sorghum cultivars, highest Al tolerance was observed for Toumitu A in both high Al (24.0%) and low Al (50.7%) conditions (Fig. 4). On the other hand, Furitsu was most Al sensitive sorghum cultivar (19.5% for high Al and 37.5% for low Al). Al tolerance distribution of sorghum was poorly distributed in high Al conditions (Fig. 4). In fact, almost no distribution could be recognized in this stage. Sorghum could be suggested as highly Al-sensitive crop species, this high Al concentration might be very lethal to the root and elongation of all the roots stops (almost) soon after the treatment. On the other hand, Al tolerance was within a range to recognize Al-tolerant and -sensitive cultivars in low Al conditions.
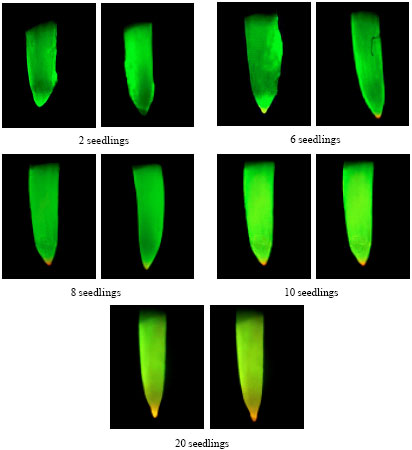 | |
| Fig. 3: | Plasma membrane permeability in control treatment. Roots were washed with deionized water and stained with fluorescein diacetate-propidium iodide (FDA-PI) for 15 min and observed under UV light. Roots emitting green fluorescene are intact and that of red fluorescence are permeabilized cells |
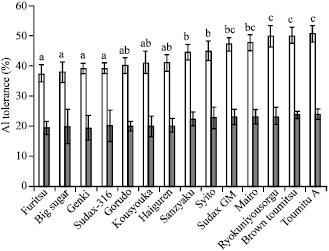 | |
| Fig. 4: | Al tolerance of sorghum cultivars for 24 h in 2.5 μM ( |
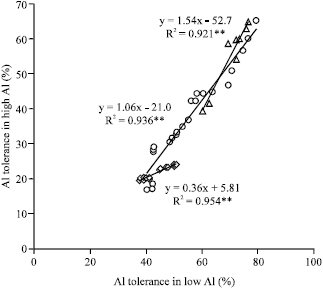 | |
| Fig. 5: | Correlation between the screening conditions (Al concentrations) for maize ( |
Correlation between the Al tolerances among the low and high Al conditions for these three crop species was significant i.e., R2 = 0.954**, 0.936** and 0.921** for sorghum, soybean and maize, respectively (Fig. 5). This result suggest that there are little variation of Al tolerance (for high Al conditions) within a crop cultivars in some cases, that can be extrapolate to other Al tolerance (e.g., low Al conditions) and vice versa.
Since, uptake of Al into root as been reported to be very rapid usually within tens of minutes and the search for the primary sites for Al toxicity and tolerance of Al has so far been elusive, the use of short-term screening techniques for differential Al tolerance, root elongation measurement has been the most popular and has even been suggested to be used as a common method to know Al tolerance (Horst, 1995) in spite of it`s inherent complexity (Rengel, 1996). The primary response to Al stress in plants occurs in roots, as reduced elongation at the tip, followed by swelling and distortion of differentiated cells (Wang et al., 2006). Within meristematic and root cap cells, Al toxicity is associated with an increased vacuolation and turnover of starch grains (De Lima and Copeland, 1994), as well as disruption of dictyosomes and their secretory function (Bennet et al., 1985; Puthota et al., 1991). Al tolerance screening are conducted based on two major criteria i.e., root length measurement and staining technique. The short-term Al tolerance screening techniques include the use of stains like Eriochrome Cyanine R (Ma et al., 1997), hematoxylin staining (Polle et al., 1978; Anas and Yoshida, 2000, 2004), FDA-PI staining (Koyama et al., 1995) and callose formation (Li et al., 2000; Anas and Yoshida, 2002) has used for sorghum screening as rapid expression of Al tolerance. In this experiment root length measurement was conducted as root length measurement is the most suitable approach for genetic and molecular studies in which a precise quantitative response for Al stress is needed (Wang et al., 2006). It is also suitable for identifying genotypes with superior alleles for Al tolerance (Hede et al., 2002). Ma et al. (2000, 2005) screened Al tolerance of rice using 50 μM AlCl3. On the other hand, Wagatsuma et al. (2005) screened Al tolerance using 20 μM AlCl3. There are references having Al tolerance study with different Al concentrations. But no report treated suitability of Al tolerance screening based on differential Al concentrations. Here we clarified that better Al tolerance screening study could be conducted selecting Al concentration based on the Al-tolerance level of specific crops. Here, two Al concetnrations and 3 crop species (several cultivars from each crop) were used to clarify the role of Al concentration on subitable Al tolerance screening. In conclusion, it could be suggest that though Al tolerance are different in low and high Al concentrations, for sensitive crop species, Al tolerance study should be done with low Al conditions. On the other hand, tolerance study for Al-tolerant crop species should be done with higher Al concentrations.
ACKNOWLEDGMENT
This study was supported by a Grant-in-Aid for Scientific Research (A) to T.W. (No. 18208008) from the Japan Society for the Promotion of Science.
REFERENCES
- Delhaize, E. and P.R. Ryan, 1995. Aluminum toxicity and tolerance in plants. Plant Physiol., 107: 315-321.
PubMedDirect Link - Grabski, S. and M. Schindler, 1995. Aluminum induces rigor within the actin network of soybean cells. Plant Physiol., 108: 897-901.
PubMedDirect Link - Ishikawa, S. and T. Wagatsuma, 1998. Plasma membrane permeability of root-tip cells following temporary exposure to Al ions is a rapid measure of Al tolerance among plant speices. Plant Cell Physiol., 39: 516-525.
Direct Link - Kochian, L.V., M.A. Pineros and O.A. Hoekenga, 2005. The physiology, genetics and molecular biology of plant aluminum resistance and toxicity. Plant Soil, 274: 175-195.
CrossRefDirect Link - Loneragan, J.F. and K. Snowball, 1969. Calcium requirements of plants. Aust. J. Agric. Res., 20: 465-478.
CrossRef - Loneragan, J.F., K. Snowball and W.J. Simmons, 1968. Response of plants to calcium concentration in solution culture. Aust. J. Agric. Res., 19: 845-857.
CrossRef - Nguyen, V.T., M.D. Burow, H.T. Nguyen, B.T. Le, T.D. Le and A.H. Paterson, 2001. Molecular mapping of genes conferring aluminum tolerance in rice (Oryza sativa L.). Theor. Applied Genet., 102: 1002-1010.
CrossRefDirect Link - Poschenrieder, C., B. Gunsé, I. Corrales and J. Barceló, 2008. A glance into aluminum toxicity and resistance in plants. Sci. Total Environ., 400: 356-365.
CrossRef - Rengel, Z., 1992. Disturbance of cell Ca2+ homeostasis as primary trigger of Al toxicity syndrome. Plant Cell Environ., 15: 931-938.
CrossRefDirect Link - Ryan, P.R., J.M. Ditomaso and L.V. Kochian, 1993. Aluminium toxicity in roots: An investigation of spatial sensitivity and the role of the root cap. J. Exp. Bot., 44: 437-446.
CrossRefDirect Link - Schreiner, K.A., J. Hoddinott and G.J. Taylor, 1994. Aluminum-induced deposition of (1,3)-betaglucans (callose) in Triticum aestivum L. Plant Soil, 162: 273-280.
Direct Link - Sivaguru, M., B. Ezaki, Z.H. He, H.Y. Tong and H. Osawa et al., 2003. Alumium-induced gene expression and protein localization of a cell wall-associated receptor kinase in Arabidopsis. Plant Physiol., 132: 2256-2266.
CrossRefDirect Link - Sivaguru, M., F. Baluska, D. Volkmann, H.H. Felle and W.J. Horst, 1999. Impacts of aluminum on the cytoskeleton of the maize root apex. Short-term effects on the distal part of the transition zone. Plant Physiol., 119: 1073-1082.
PubMedDirect Link - Thaworuwong, N. and A. Van Diest, 1974. Influence of high acidity and aluminum on the growth of lowland rice. Plant Soil, 41: 141-159.
CrossRefDirect Link - Wagatsuma, T., M.S.H. Khan, I.M. Rao, P. Wenzl, K. Tawaraya and T. Yamamoto et al., 2005. Methylene blue stainability of root-tip protoplasts as an indicator of aluminum tolerance in a wide range of plant species, cultivars and lines. Soil Sci. Plant Nutr., 51: 991-998.
CrossRef - Zhang, G.C., J. Hoddinott and G.J. Taylor, 1994. Characterization of 1,3-β-D-glucan (callose) synthesis in roots of Triticum aestivum in response to aluminum toxicity. J. Plant Physiol., 144: 229-234.
CrossRefDirect Link - Zhang, J., Z. He, H. Tian, G. Zhu and X. Peng, 2007. Identification of aluminum-responsive genes in rice cultivars with different aluminium sensitivities. J. Exp. Bot., 58: 2269-2278.
CrossRefPubMedDirect Link - Ma, J.F., S. Taketa and Z.M. Yang, 2000. Aluminum tolerance genes on the short arm of chromosome 3R are linked to organic acid release in triticale. Am. Soc. Plant Physiol., 122: 687-694.
Direct Link - Koyama, H., K. Ojima and T. Yamaya, 1990. Utilization of anhydrous aluminum phosphate as sole source of phosphorus by a selected carrot cell line. Plant Cell Physiol., 31: 173-177.
Direct Link - Wang, J.P., H. Raman, G.P. Zhang, N.J. Mendham and M.X. Zhou, 2006. Aluminium tolerance in barley (Hordeum vulgare L.): Physiological mechanisms, genetics and screening methods. J. Zhejing Univ. Sci. B, 7: 769-787.
CrossRefDirect Link - De Lima, M.L. and L. Copeland, 1994. Changes in the ultra-structure of the root tip of wheat following exposure to aluminium. Aust. J. Plant Physiol., 21: 85-94.
Direct Link - Bennet, R.J. C.M. Breen and M.V. Fey, 1985. The primary site of aluminium injury in the root of Zea mays L. S. Afr. J. Plant Soil, 2: 8-17.
CrossRefDirect Link - Puthota, V., R. Cruz-Ortega, J. Johnson and J. Ownby, 1991. Ultrastructural Study of the Inhibition of Mucilage Secretion in Wheat Root Cap by Aluminium. In: Plant-Soil Interaction at Low Soil pH. Wright, R.J., V.C. Baligar and R.P. Murrmann (Eds.). Kluwer Academic Publishers, Dordrecht pp: 779-787.
- Ma, J.F., S.J. Zheng, X.F. Li, K. Takeda and H. Matsumoto, 1997. A rapid hydroponic screening for Al-tolerance in barley. Plant Soil, 191: 133-137.
CrossRefDirect Link - Polle, E., C.F. Konzak and J.A. Kittrick, 1978. Visual detection of Al tolerance levels in wheat by hematoxylin staining of seedling roots. Crop Sci., 18: 823-827.
Direct Link - Anas and T. Yoshida, 2000. Screening of Al-tolerant sorghum by hematoxylin staining and growth response. Plant Prod. Sci., 3: 246-253.
Direct Link - Anas, A. and T. Yoshida, 2004. Heritability and genetic correlation of Al-tolerance with several agronomic characters in sorghum assessed by hematoxylin staining. Plant Prod. Sci., 7: 280-282.
Direct Link - Koyama, H., T. Toda, S. Yokota, Z. Deqair and T. Hara, 1995. Effects of aluminum and pH on root growth and cell viability in Arabidopsis thaliana strain Landsberg in hydroponic culture. Plant Cell Physiol., 36: 201-205.
CrossRefDirect Link - Li, X.F., J.F. Ma, S. Hiradate and H. Matsumoto 2000. Mucilage strongly binds aluminum but does not prevent roots from aluminum injury in Zea mays. Physiol. Plant., 108: 152-160.
Direct Link - Anas, A. and T. Yoshida, 2002. Genotypic difference of Sorghum bicolor in the callus formation and callus growth on aluminum-containing medium. Plant Prod. Sci., 5: 242-247.
Google - Hede, A.R., B. Skovmand, J.M. Ribaut, D. Gonzalez-de-Leon and O. Stolen, 2002. Evaluation of aluminum tolerance in a spring rye collection by hydroponic screening. Plant Breed., 121: 241-248.
CrossRef