
E. Luvaha
Department of Botany and Horticulture, P.O. Box 333, Maseno, Kenya
G.W. Netondo
Department of Botany and Horticulture, P.O. Box 333, Maseno, Kenya
G. Ouma
Department of Botany and Horticulture, P.O. Box 333, Maseno, Kenya
American Journal of Plant Physiology
Year: 2008 | Volume: 3 | Issue: 1 | Page No.: 1-15







ABSTRACT
Six month old Mango (Mangifera indica L.) rootstock seedlings were grown in 20 L plastic pots in a greenhouse at Maseno University, Kenya. to investigate the effect of water deficit on its morphological and physiological characteristics such as plant height, number of leaves, stem diameter and gas exchange characteristics and chlorophyll content, respectively. A Completely Randomized Design (CRD) with four treatments and six replications was used. The treatments involved subjecting the rootstock seedlings to four different irrigation regimes namely watering daily, twice in a week, once in a week and once in two weeks. The measurements were taken after every two weeks for a period of three months. At the end of the experiment, destructive sampling to establish the root to shoot ratio were taken. The soil moisture content under the different irrigation regimes was also determined gravimetrically. Growth parameters increased under mild water stress except under extreme water deficit where there was wilting. Root to shoot ratio increased with increasing water deficit. Increase in water deficit reduced the gas exchange parameters but slightly increased chlorophyll content. It is concluded that water deficit significantly (p≤0.05) affects physiological and morphological characteristics of Mango.
PDF Abstract XML References Citation
How to cite this article
E. Luvaha, G.W. Netondo and G. Ouma, 2008. Effect of Water Deficit on the Physiological and Morphological Characteristics of Mango (Mangifera indica) Rootstock Seedlings. American Journal of Plant Physiology, 3: 1-15.
DOI: 10.3923/ajpp.2008.1.15
URL: https://scialert.net/abstract/?doi=ajpp.2008.1.15
DOI: 10.3923/ajpp.2008.1.15
URL: https://scialert.net/abstract/?doi=ajpp.2008.1.15
INTRODUCTION
Water deficit is a problem to plant growth and crop productivity in the vast dry land areas of Kenya. Irrigation is the main solution to this problem. Alternatively drought tolerant trees may be grown in these areas. It has been hypothesized that certain perennial plants are drought resistant but there is need to investigate the mechanisms by which these plants endure drought. Drought is a major factor limiting growth and development in higher plants and it is a common occurrence in many environmental conditions. As a result, perennial plants species have developed mechanisms to cope with an inadequate water supply. Plants avoid water deficit by developing deep roots or by minimizing water loss (e.g., stomatal closure, small leaves). Some species tolerate water deficit through osmotic or elastic adjustment or by the accumulation of osmoprotective substances such as cyclitols (Kozlowski et al., 1991). Species possessing both a drought avoidance mechanisms and an ability to acclimatize by active osmoregulation would be advantageous because of increased flexibility in response to changing environmental conditions. Osmotic adjustment may provide an ecological advantage for young seedlings by maintaining metabolic activity under suboptimal conditions during establishment when roots have not reached deep soil water (Kozlowski et al., 1991).
Water deficit affects growth, development, yield and quality of fruit trees in the greenhouse and field conditions (Clifford et al., 1998; Ma et al., 2006). Water deficit also affects gas exchange parameters (Barry et al., 2004; Pavel and de villiers, 2004), increases water use efficiency (Arndt et al., 2001) and alters carbon redistribution in favour of carbohydrates (Arndt et al., 2001). In mature mango trees water deficit reduces vegetative growth (Pavel and de villiers, 2004) .
The biggest portion of the land mass in Kenya is either arid or semi-arid. Mango is grown in this portion because of soil suitability but there is a problem of inadequate rainfall thus necessitating irrigation. The effect of water deficit on the growth and development of mango seedlings is not known by nursery producers and so is the frequency of irrigation which optimizes growth. The hypothesis of the present study is that increase in water deficit reduces growth and gas exchange parameters of mango seedlings. To prove this hypothesis and achieve the undermentioned objectives modern porometric methods will be applied to measure gas exchange parameters and standard methods used to determine the growth parameters.
Mango is a crop of major economic importance in Kenya produced mainly in the dry areas of Kenya. Irrigation is therefore necessary to ensure stable yields of high quality. It is therefore a necessary to investigate the effect of different level of water deficit on the morphological and physiological parameters of mango rootstock seedlings at Maseno, Kenya. The objectives of the present study were to investigate the effects of water deficit on physiological parameters of mango root stock seedlings and to investigate the effects of water deficit on morphological parameters of mango root stock seedlings.
MATERIALS AND METHODS
Six months old mango rootstock seedlings were transplanted in Polyvinyl (PVC) pots measuring 20 cm in diameter and 30 cm in height in a greenhouse at Maseno University Botanic Garden, Kenya from December 2003 to March 2004. The soils in Maseno are classified as very fine, kaolinitic, isohyperthermic kandiudalfic Eutrudox. Soils physical and chemical characteristics at a depth of 0.15 m were; pH 5.6 (in a 1:2.5 soil/water suspension)/organic C = 14.0 g kg-1, extractable P = 1.3 mg kg-1 exchangeable K = 0.3 cmolc kg-1, exchangeable Ca = 5.4 cmolc kg-1, Mg = 1.70 cmolc kg-1 and exchangeable acidity = 0.5 cmolc kg-1; sand = 27%; clay = 52%; silt = 21% and bulk density = 1.3 g cm-3 (Netondo, 1999). The Pots, containing soil mixture at a ratio of 1 sand: 2 loam: 3 compost were perforated at the bottom and placed on tables in the green house. All agronomic practices including weeding, pest control and fertilizer application were observed apart from watering (irrigation) which was controlled. The minimum and maximum temperatures were determined using a thermometer and relative humidity using a hygrometer. The mean values for temperature were 22 and 38°C, respectively, while that of relative humidity was 38+5%.
The experimental set up was a Completely Randomized Design (CRD) with four treatments and six replications. The four treatments were; watering daily (W), watering twice in week (X), watering once in one week (Y) and once in two weeks (Z). Soil moisture content varied with different treatments (Fig. 1).
Measurements were taken after every two weeks for a period of three months. The soil moisture content was determined gravimetrically. Gravimetric measurements were taken on the topsoil, at 10 cm from the top. The soil samples from each treatment were weighed (W1) then dried in an oven for 48 h at 100°C and the dry weight (W2) obtained. The percentage water content (W) was calculated as;
| Where: | ||
| W1 | = | Fresh weight, |
| W2 | = | Dry weight, |
W | = | Percentage soil moisture content. |
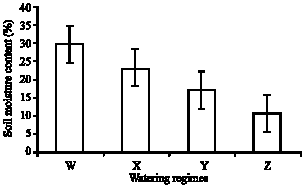 |
| Fig. 1: | The soil moisture content under different watering regimes (W, X, Y and Z) determined gravimetrically at Maseno, Kenya. W-Watering daily, X-Wateringtwice a week, Y-Watering once a week, Z-Watering once in two weeks |
The stem diameter was measured by a hand caliper, plant height with a meter rule and number of leaves were individually counted on a weekly basis. To get the root: shoot ratio, the whole plant was uprooted, rinsed, separated into shoot and root and oven dried for 48 h at 72°C. The root and shoot dry weights were measured with an electronic weighing balance. Root: shoot ratio was computed as.
The data was analyzed using the SAS Statistical computer package to obtain analysis of variance (ANOVA) tables and the correlation between various parameters.
Measurement of Gas Exchange Parameters and Chlorophyll Content
Measurement of parameters was taken for a period of 4 months during the duration of the experiment from December to March 2004. The parameters determined were CO2 assimilation, transpiration rate, stomatal conductance, leaf temperature, internal CO2 concentration. These measurements were taken on the most recently emerged, full expanded and well exposed leaves under bright light using an infrared gas analyzer (CIRAS 1-PPSystem, Stortfield, Hitchin, Herts, U. K.): Chlorophyll content was also determined using standard methods.
RESULTS
Soil Moisture Content
The soil moisture content results indicated a highly significant difference between treatment W (the control) and Z (extremely stressed plants.) No significant differences existed between treatment X and Y, which will in this context be referred to as plants exposed to mild water deficit. Figure 1 shows the percentage soil moisture content under each of the treatments.
Effect of Water Deficit on Plant Height
Plants exposed to mild water deficit (X and Y) had a higher growth rate compared to the control (W). There was an observed increase in height under all treatments except in Z when a decline was observed from D42. A significant difference (p≤0.05) between treatments was observed on all the days. The highest increase in height was in treatment Y then X, W and finally Z in that order (Fig. 2).
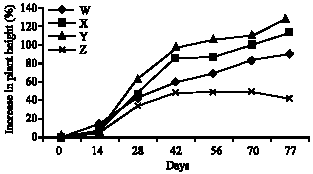 |
| Fig. 2: | The effect of water deficit on plant height of mango root stock seedlings measured as the length of the plant above the soil surface grown at Maseno, Kenya. W-Wateringdaily, X-Watering twice in a week, Y-Watering once in a week, Z-Watering once in two weeks |
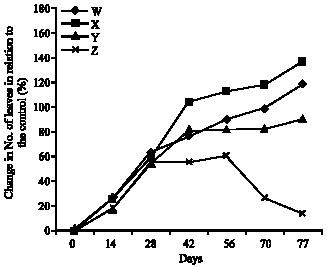 |
| Fig. 3: | The effect of water deficit on the number of leaves of mango rootstock seedlings grown at Maseno, Kenya. W-Watering daily, X-Watering twice in a week, Y-Watering once in a week, Z-Watering once in two weeks |
Effect of Water Deficit on the Number of Leaves
There was a general increase in the number of leaves except in treatment Z where a decline was observed after D28. Plants under mild stress (X and Y) showed a higher increased compared to the control (W). A significant difference (p≤0.05) was observed between W and X as compared to Y and Z on D0 and D14 after which there was no observed significant differences between treatments. There was also a decline in number of leaves in treatment Z due to drying/senescence of lower mature leaves hence a highly significant difference (p≤0.001) was observed between treatment Z and the rest (W, X and Y) on D70 (Fig. 3).
Effect of Water Deficit on Stem Diameter
A steady increase in the diameter was observed in the control (W) and treatment X. There was however no marked significant differences (p≤0.05) between the treatments apart from at D14. At D70 and D72 there was a significant difference (p≤0.05) between treatments W and Z as well as X and Z (Fig. 4).
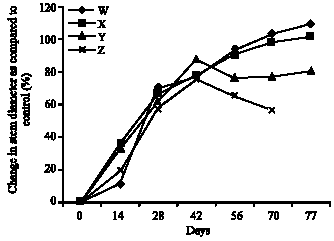 |
| Fig. 4: | The effect of water deficit on stem diameter of mango rootstock seedlings, measured 10 cm above the soil surface, grown at Maseno, Kenya W-Watering daily, X-watering twice in a week, Y-Watering once in a week, Z-Watering once in two weeks |
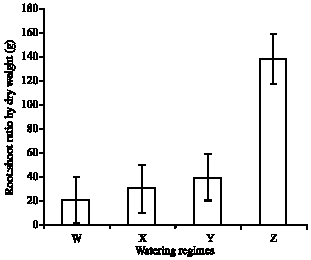 |
| Fig. 5: | The root: shoot ratio of M. indica rootstock seedlings grown under four different watering regimes (W, X, Y and Z). W-Watering daily, X-Watering twice in a week, Y-Watering once in a week, Z-Watering once in two weeks |
Effect of Water Deficit on Plant Biomass
The root: shoot ratio by weight increased with increasing water deficit. The values for the highly stressed seedlings were significantly higher than those of well-watered plants. There was more extensive growth of both adventitious and taproots in seedlings exposed to water deficit (Z) as compared to the control (W) (Fig. 5).
Effects of Water Deficit on Gas Exchange Parameters and Chlorophyll Content
Transpiration
The rate of transpiration was higher in the well-watered plants compared to the extremely stressed plants on all the days, except on D0 and D14. The lowest rates of transpiration in all treatments were observed on D14 when the temperatures in the green house were lowest (24°C) and the humidity was higher (40%) than observed on all other days of data collection. On D0, there was no significant differences between the treatments. However on D14 there was a significant difference (p≤0.05) between the treatments.
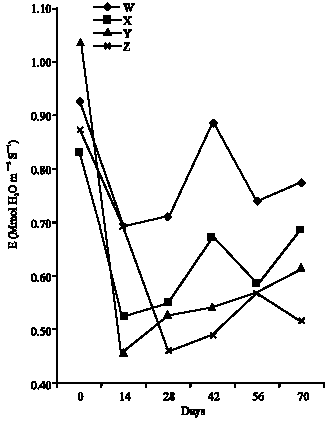 |
| Fig. 6: | The effect of water stress on the rate of transpiration (E) in mango rootstock seedlings W-Watering daily, X-Watering twice in a week, Y-Watering once in a week, Z-Watering once in two weeks |
At D56 a decline in the rate of transpiration was observed under extreme water stress (treatment Z) as compared to W, X and Y where an increase in transpiration was observed. Highly significant differences (p≤0.001) were observed between the treatments at D28, D42, D56 and D70 (Fig. 6).
Stomatal Conductance
The trend in stomatal conductance is almost similar to that of transpiration. The stomatal conductance was highest in the well-watered plants (W) and lowest in the extremely water stressed plants (Z) on all days except on D0 and D28. At D0 there was no significant difference between treatments. However, a highly significant difference (p<0.001) occurred on D14 and D42. The same case was observed between D56 and D70 (Fig. 7).
There was a steady increase in CO2 assimilation with time except on D42 where a decline occurred. In the highly stressed plants (Z) had a decline was observed from D56 to D70. Significant differences (p≤0.05) occurred between particular treatments on certain days. Plants that were highly stressed (treatment Z) apparently had a higher photosynthetic rate than W and Y, which received more water implying a possible resistance of the photosynthetic apparatus to water stress (Fig. 8).
Intercellular CO2 Concentration (Ci)
There was an initial increase in Ci among the treatments from D0 to D28 except in treatment 2 when a decline was first observed at D14 then the Ci concentration rose up. A significant difference (p≤0.05) was observed between the treatments at D28. Generally Ci concentration was higher in the highly stressed plants (Z) as compared to W, X and Y.
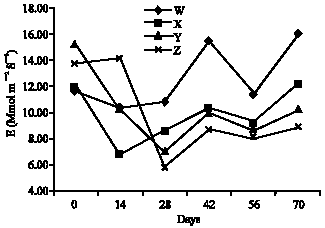 |
| Fig. 7: | The effect of water stress on leaf stomatal conductance (g) of mango rootstock seedlings grown at Maseno University, Kenya. W-watering daily, X-watering twice in a week, Y-watering once in a week, Z-watering once in two weeks |
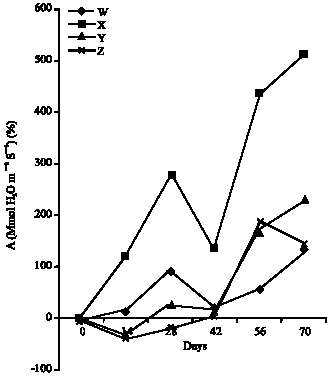 |
| Fig. 8: | The effect of water stress on the rate of CO2 assimilation (A) on the leaves of mango rootstock seedlings W-watering daily, X-watering twice in a week, Y-watering once in a week, Z-watering once in two weeks |
A highly significant difference (p≤0.001) was also observed among treatments at D56 where the more stressed plants (Y and Z) had a higher Ci than W and X (Fig. 9).
Leaf Temperature
The general trend of the graph shows a higher leaf temperature in treatment Y and Z which were the most stressed as compared to W and X. A highly significant difference (p≤0.001) was observed between treatments on all the days except at D0 (Fig. 10).
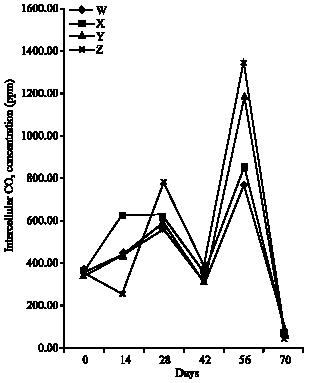 |
| Fig. 9: | The effect of water stress on intercellular CO2 concentration (Ci) on mango rootstock seedlings. W-watering daily, X-watering twice a week, Y-watering once a week, Z-watering once in two weeks |
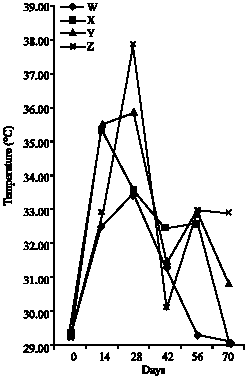 |
| Fig. 10: | Effect of water stress on leaf temperature of mango roostock seedlings grown at Maseno University, Kenya. W-watering daily, X-watering twice in a week, Y-watering once in a week, Z-watering once in two weeks |
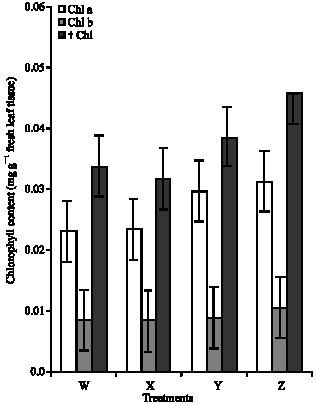 |
| Fig. 11: | The effect of water stress on chlorophyll extracted from fresh leaves of mango rootstock seedlings. W-watering daily, X-watering twice in a week, Y-watering once in a week, Z-watering once in two weeks |
Chlorophyll Content
There was a steady rise in chlorophyll a and total chlorophyll content with increase in water stress as shown in Fig. 11. Values for chlorophyll b however remained almost constant in all treatments except in Z when there was a slight increase. Total chlorophyll showed a general increase with stress, especially between treatment Y and Z.
DISCUSSION
Effect of Water Deficit on Soil Moisture Content
There was an observed decrease in soil moisture content with increasing water deficit. The soil moisture content mainly controls the status of the plant water potential (Levitt, 1980). It is important to maintain proper soil moisture since there is a very close relationship between the soil moisture and crop yield. Moisture requirements for plants, however differ with species, stage of development and plant age. (Kramer, 1983). In present study losses of moisture from the soil may be attributed to surface evaporation, transpiration and drainage. Under field conditions, surface evaporation may be checked through cultural practices such as mulching (Singh, 1980).
Effect of Water Deficit on the Morphological Characteristics
Growth involves both cell growth and development. Cell growth and development is a process comprising three stages namely cell division, cell enlargement and cell differentiation (Hsiao, 1973). These processes are very sensitive to water deficit- because of their dependence on cell turgor. The low increase in plant height under extreme water deficit may have been due to reduced cell turgor which affected cell division and expansion.
However, cell division has been reported to be less sensitive to water deficit than cell expansion or enlargement (Hsiao, 1973). Turgor pressure in growing cells provides the driving force for cell expansion (Jones, 1992). Therefore, reduced growth rate under water deficit can be qualitatively related to reduced cell turgor or a reduction in cell wall extensibility. Cell turgor decreases with any dehydration-induced decrease in cell water potential. Severe water deficit reduces stem elongation in Penium maximum (Corlette et al., 1994) as in this study-with mango seedlings. Mild water deficit (X and Y) increased plant height. This may be difficult to explain due to the fact that the control received excess water which caused oxygen deficiency that inhibited plant growth (Levitt, 1980). Excess water may also have leached away some soil nutrients causing nutrient deficiency which reduced plant growth under such irrigation treatments.
Water stress or deficit also increases the senescence of leaves in wheat (Teare et al., 1982). Observed wilting in mature leaves due to water deficit may be associated with carbohydrate depletion due to mobilization and export followed by senescence. Growth inhibition after wilting enhances nucleic acid destruction (Hsiao, 1973). Water deficit reduces protein synthesis and causes destruction of polysomal mRNAS in the zone of elongation of the hypocotyls (Hsiao, 1973).
Generally, plants show increased root to shoot ratio under water deficit conditions (Westgate and Boyer, 1985). Similar results were obtained in our study with M. indica rootstock seedlings. This is an indication of adaptation for survival in drought areas since increased root surface area allows more water to be absorbed from the soil. However, since water deficit causes a decline in growing zones of roots and hypocotyls, other factors, which may be genetic or metabolically controlled may also be involved (Creelman et al., 1990). The dissimilar response of roots and shoots to water deficit could be resulting from roots being closer than shoots to an external supply of water. A reduction in shoot growth coupled with continued root growth would result in an improved plant water status under extreme water deficit conditions. Similar results have also been observed on other crops growing under water deficit conditions such as cotton (Ackerson and Kreig, 1977) and soybean (Creelman et al., 1990). Further, plant growth regulators may also affect plant growth pattern under low water potential. Absiscic Acid (ABA) accumulation in the hypocotyls region in water stressed plants inhibits growth but this does not affect root growth as in maize plant (Raymond et al., 1987) Other than low water potential and ABA accumulation, continued root growth under water deficit may also have been due to the accumulation of sugars in the root tips. This implies that there is a continued supply of import of assimilates into the roots. In wheat, Jones (1987) proved that the accumulation of ABA in the hypocotyls causes a redistribution of assimilates from the hypocotyls in the roots. This may explain the behaviour of Mango seedlings in this study.
In this study, root growth may have been reduced by the use of pots. This restricted root growth has negative consequences on shoot growth (Radin et al., 1987). In maize seedlings, for example, root growth continues at very low water potential which are completely inhibitory to shoot growth (Sharp and Davies, 1979). However, under normal field conditions both the tap and adventitious roots grow very extensively in order to trap the scarce water from deeper soil horizons. In case of extreme water deficit root development is unlikely to occur causing further stress. There was no significant difference between the control and the plants under mild water deficit in the root to shoot ratio implying that these plants can survive some degree of water deficit.
Effects of Water Deficit on Gas Exchange Parameters
Transpiration decreased significantly in the plants under extreme water deficit as compared to the control. Similar results have been reported in beans (Ouma, 1988), soybeans (Wahbi and Sinclair, 2007), tomato (Xu et al., 1995), wheat (El Hafid et al., 1998), in pears (Ma et al., 2006) and in peach (DeJong et al., 2005). The transpirational pathway of water flow in woody plants is the xylem in which the conducting elements are non-living and heavily thickened and lignified tracheids and xylem vessels (Jones, 1992). CO2 assimilation rates were also reduced and this agrees with previous studies (DeJong et al., 2005; Ma et al., 2006) in peach and pear trees, respectively. Apparently, there is a correlation between transpiration and photosynthesis. Similar results were reported in sunflower by Robertson et al. (1985). Under water deficit cells lose their turgidity causing stomatal closure. This limits the rate of CO2 diffusion through the stomata causing a decline in the photosynthesis. The rate of transpiration is also affected by either environmental factors such as temperature, radiation and relative humidity (Jones, 1992). The fluctuations observed in transpiration on specific days such as D42 (Fig. 2) are due to daily changes in temperature and relative humidity. Higher air temperatures increase the rate of transpiration. The effect of this is probably to enhance the cooling of leaves by evaporation (Burke et al., 1990).
Osmotic effects may also have played a role in the decrease of cell division and expansion. Although this parameter was not measured in our study, it is known that water deficit causes low water potential which increases the osmotic potential in plants.
The water potential at which plants get stressed however varies with species, plant age and stage of development (Levitt, 1980). In oak seedlings, for example, growth of the seedlings decreased as the osmotic potential of the soil solution decreased from 0.03 to -0. to -0.8 Mpa (Larson and Pashely, 1973) and sunflower showed a sharp reduction in shoot growth as the height dropped below 0. 032 Mpa (Sionit et al., 1973). Growth in height and diameter of cotton stems reduced even when 35% of the root was in soil at a water potential of -0.1 Mpa (Klepper et al., 1973. These results show that increase in height in mango seedlings is reduced only under severe water deficit implying that the mango can withstand certain levels of dehydration by osmotic adjustment through active solute accumulation in the leaves (Clifford et al., 1998). The number of leaves of the mango seedlings was also reduced by water deficit. A significant increase was observed in the number of leaves in plants under mild water deficit compared to those under severe deficit, as was observed in plant height. Severe water deficit has reduced leaf development in many crops (Ouma, 1988). The reduction in leaf number under extreme water deficit may have been due to reduction in leaf formation and abscission of lower leaves eventually leading to the wilting of the whole plant. Water deficit reduces leaf growth by reducing rates of cell division and expansion due to turgor loss and increased synthesis of abscission acid (Tezera et al., 2002). Water deficit also causes reduced leaf initiation (Boyer, 1976). It is apparent from our study and those of Tezera et al. (2002) that as in sunflower mild water deficit does not inhibit the growth of mango seedlings. The reduction of leaf number under severe water deficit was partially due to leaf senescence caused by increased carbon remobilization from the leaves and their preferential redistribution to stems and roots resulting in leaf shedding (Arndt et al., 2001).
Reducing the number of leaves could be a phenomenon by the plants to reduce transpiration surface hence water loss. Similar behaviour has been reported in sorghum exposed to severe water deficit It shows an initial decrease in the daily increment due to accelerated senescence (McCree, 1985). In cotton, water deficit causes a reduction in leaf development and an increase in leaf senescence (Fernandez-Conde, 1998).
Leaf temperature however increased with decline in transpiration. Transpiration cools the plant by loss of latent heat of vaporization. Wilting of some seedlings under extreme deficit was observed afterD56. Plants were unable to absorb capillary water which was tightly held by soil. The reduction in transpiration rate in plants under water deficit may also be attributed to morphological changes such as increased cell wall thickness and cell wall lignifications (Netondo, 1999).
Stomatal conductance took a similar trend like that of transpiration. Stomatal conductance in the water stressed plants was generally lower as compared to the well-watered plants. Reduction of leaf water potential led to the development of a water deficit in the leaves causing guard cells to loose turgor and hence stomatal pores to reduce. In addition the increased resistance may have led to reduced water transport in the leaves further causing a decrease in the stomatal conductance. Reduction in stomatal conductance decreases transpiration and limits photosynthesis (Tezera et al., 2002) as demonstrated in present study. In some plants, stomatal conductance declines even before severe water deficit sets in thereby avoiding desiccation during drought. This has been observed in Quercus ilex (Fortelli et al., 1986). Leaves of plants exposed to higher water deficits have higher ABA concentration. This may have also contributed to regulation of closing and opening of the stomata in the leaves of the mango rootstock seedlings. Several workers have reported reduction of stomatal conductance from increased water deficit (Dejong et al., 2005).
Net photosynthesis (CO2 assimilation) seems to increase in all treatments except the rate was higher in the well-watered plants as compared to the stressed ones. However no significant difference was observed (Fig. 2). CO2 assimilation is affected by both stomatal and non-stomatal factors. In our study, it appears that the photosynthetic apparatus may have been resistant to dehydration since there was no decline in net CO2 under extreme water deficit. The lower rate of increase in the CO2 assimilation under water deficit may be attributed to reduced stomatal conductance. However, in order for stomatal closure to have an effect on both transpiration must at least be limited by rate of CO2 diffusion through the stomata (Boyer, 1976).
There was a higher Water Use Efficiency (WUE) in the stressed plants as compared to the well watered. Water use efficiency is the ratio of leaf photosynthesis to transpiration (A/E) measured simultaneously (El Hafid et al., 1998) or the carbon gained during photosynthesis in relation to the water lost during transpiration (Hsiao, 1973). Similar results were obtained in pawpaw plants (Carica papaya) (Clemente and Marler, 1996). This is a major adaptive behaviour for plants growing in dry areas. However, there is a consequence of maximizing WUE by minimizing water-loss, carbon acquisition is probably an adaptation for mangoes to water limited areas.
Another important factor, which may have a profound effect on CO2 assimilation in the plants, is the internal CO2 concentration (Ci). In this study CO2 assimilation seemed not to be affected by water stress. Therefore the low increase in CO2 assimilation under water stress. without a corresponding decline in Ci, could be due to non-stomatal effects on the photosynthetic processes, possibly an increase in the mesophyll resistance as suggested by Cornic et al. (1989). Similar results were reported in wheat (Kecheva et al., 1994). A reduction in Ci can be very detrimental to the photosynthetic process especially in the presence of enzyme Rubisco which has a high affinity for oxygen (O2) when the intercellular CO2 concentration is low. Therefore under low Ci, photosynthesis is limited by enzyme Rubisco. For many species, Ci tends to remain constant over a range of environmental conditions, including water deficit (Pearcy, 1981).
The chlorophyll content is another factor that affects the photosynthetic process in green plants. In our study, however, chlorophyll a was a more resistance to hydration, it increased slightly with water deficit as compared to chlorophyll b, which was constant. The slight increase in total chlorophyll under water deficit suggests that the chlorophyll pigments in these leaves were somewhat resistant to dehydration.
CONCLUSIONS
From the present study it can be concluded that water deficit affects the growth and physiological characteristics of Mango rootstock seedlings. The rate of transpiration reduced with increasing water deficit. Transpiration is controlled by other environmental factors such as temperature, humidity and air movements. These factors are however controlled in the greenhouse where we conducted this studies. As transpiration is reduced leaf temperature is increased. These confirm that the transpiration process cools the leaf surface by reducing latent heat of evaporation. Drying and shedding of lower leaves observed under extreme water deficit in our studies is a mechanism for water conservation. The rate of CO2 assimilation and intercellular CO2 concentration and chlorophyll content were affected by water deficit in our study. A reduction in transpiration coupled with an increase in CO2 assimilates implies that photosynthesis was largely controlled by non-stomatal factors. Continued CO2 assimilation under water deficit is only possible in the drought tolerant plants. This implies that the photosynthetic apparatus is resistant to increase in water deficit. Increase in chlorophyll a may be due to continued synthesis of this pigment. Even under stress conditions reduction in growth rate of the M. indica seedlings was only observed under extreme stress. However growth rate under mild stress was higher than under well-watered conditions. Under extreme water deficit, growth rate continued at a lower rate until wilting set in. This is a clear indication that the seedlings can withstand stress up to a certain limit. From the present studies watering once a week would be most recommended for potted seedlings in a green house.
Shoot growth was reduced under extreme water deficit as compared to the plants watered daily. Shoot to root ratio is also an adaptation to water deficit in most plants growing under arid conditions to increase the surface area for water absorption while reducing transpiration. In our study, the proliferation of the roots was inhibited by the polythene pots and there was exhaustion of plant nutrients in the soil due to the small volumes of soil used in the pots and continuous leaching of these nutrients during watering.
ACKNOWLEDGMENTS
In conducting our research project we wish to thank most sincerely the Botanic Gardens of Maseno University for offering the greenhouse facilities and polythene pots Further we wish to thank Mr. Peter Olewe for helping with data collection. The study leave provided by the Lake Basin Development Authority Kenya to the first author is gratefully acknowledged. Last but not least, we wish to thank most sincerely the Secretarial staff namely Caroline Ogenga, Agatha Nyayieka and Irene Ogonda for their typing service and to the Vice Chancellor, Prof. F. Onyango Maseno University for creating an enabling environment for research and scholarly activities.
REFERENCES
- DeJong, T.M., T. Cao, M. Ahmed and K.S. Shackel, 2005. Water stress and crop load effects on vegetative and fruit growth of elegent, lady. Peach (Prunus persica L.) (Batch trees). Fruits, 60: 55-68.
CrossRef - El Hafid, D.H. Smith, M. Karrou and K. Samir, 1998. Physiological responses of spring Durum wheat cultivars to early season drought in a Mediterranean environment. Ann. Bot., 81: 363-370.
Direct Link - Fortelli, M.N., K.M. Radoglon and H.A. Constantnidon, 1986. Water stress responses of seedlings of four Mediterranean oak species. Tree Physiol., 20: 1065-1075.
CrossRefPubMedDirect Link - Hsiao, T.C., 1973. Plant responses to water stress. Annu. Rev. Plant Physiol., 24: 519-570.
CrossRefDirect Link - Jones, H.G., 1987. Repeated flowering in apple caused by water stress or defoliation. Trees: Struct. Function, 1: 135-138.
CrossRef - Kecheva, M.L., T.D. Tsoner and L.P. Popova, 1994. Stomatal and non-stomatal limitation in photosynthesis in two wheat cultivars subjected to water stress. Photosynthetica, 30: 107-116.
Direct Link - Klepper, B., H.M. Taylor, M.G. Hulck and E.L. Fiscus, 1973. Water relations and growth of cotton in drying soil. Agron. J., 65: 307-310.
Direct Link - Larson, M.M. and I. Pashely, 1973. Effects of water stress and gibberellic acid on the initial growth of oak seedlings. Can. J. For. Res., 3: 75-82.
Direct Link - McCree, K.J., 1986. Whole plant carbon balance during osmotic adjustment to drought and salinity stress. Aust. J. Plant Physiol., 13: 33-43.
CrossRefDirect Link - Pavel, W. and A.J. de villiers, 2004. Responses of mango trees to reduced irrigation regimes. Acta Hortic., 645: 63-68.
Direct Link - Pearcy, R.W., 1981. Some relationships between the biochemistry of photosynthesis and gas exchange of leaves. Planta, 153: 376-387.
Direct Link - Radin, J.W., B.A. Kimball, D.L. Hendricks and L. Manney, 1987. Photosynthesis of cotton plants exposed to elevated levels of carbon dioxide in the field. Photosynth. Res., 12: 191-203.
Direct Link - Raymond, P., M. Sangy and P.E. Pilet, 1987. Quantification of abscissic acid in a single maize root. Plant Physiol., 85: 8-9.
Direct Link - Robertson, J.M., R.P. Pharis, Y.Y. Huang, D.M. Reid and E.C. Yeung, 1985. Drought induced increases in abscissic acid levels in the root apex of sunflower. Plant Physiol., 79: 1086-1089.
Direct Link - Sharp and W.J. Davies, 1979. Solute regulation and growth by roots and shoot of water stressed maize plants. Planta, 147: 43-49.
Direct Link - Teare, I.D., S. Nasser and P.J. Kramer, 1982. Changes in water status during water stress at different stages of development in wheat. Physiol. Plant., 55: 296-300.
Direct Link - Tezara, W., V. Mitchell, S.P. Driscoll and D.W. Lawlor, 2002. Effects of water deficit and its interaction with CO2 supply on the biochemistry and physiology of photosynthesis in sunflower. J. Exp. Bot., 53: 1781-1791.
CrossRef - Wahbi, A. and T.R. Sinclair, 2007. Transpiration response of arabidopsis, maize and soybean to the drying of artificial and mineral soil. Environ. Exp. Bot., 59: 188-192.
CrossRef - Westgate, M.E. and J.S. Boyer, 1985. Osmotic adjustment and the inhibition of leaf, root, stem and silk growth at low water potentials in maize. Planta, 104: 540-549.
CrossRef