
Deepak Ganjewala
Central Institute of Medicinal and Aromatic Plants, P.O. CIMAP, Lucknow-226 015 (U.P.), India
LiveDNA: 91.1338
ORCID: 0000-0003-3911-1333
Rajesh Luthra
Human Resources and Development Group, CSIR Complex, Library Avenue, Pusa, New Delhi-110 012, India
American Journal of Plant Physiology
Year: 2007 | Volume: 2 | Issue: 4 | Page No.: 269-275







ABSTRACT
Cymbopogon flexuosus (Nees ex Steud) Wats mutant cv. GRL-1 leaves obtained from different developmental stages (10 to 50 d) were fed in vivo with 5 μCi μmole-1 [2-14C]- acetate (activity 0.1 mCi, specific activity 34.51 mCi/mmole). The essential oil, geranyl acetate and geraniol (free + bound geraniol as part geranyl acetate) biosynthesis was at peak during the initial (10-20 days) leaf developmental stages. The ratio of relative % distribution of label incorporated into geranyl acetate to that of geraniol decline substantially during leaf development (10 to 50), thereby suggesting the role of geranyl acetate esterase (GAE) in transformation of geranyl acetate to geraniol. Also, it could be said that acetylation capacity of the leaves was maximum during initial growth phase. The relative % distribution of label incorporated in geraniol (as part of geranyl acetate) and acetate moiety isolated after hydrolysis of geranyl acetate have shown that bound geraniol biogenesis was maximum at 10 days and in later stages there was no enhancement in the label incorporation. At 50 days most of the radioactivity was appeared in free geraniol as at this stage acetylation capacity of the leaves was virtually negligible. The study revealed that only young and rapidly expanding leaves have the capacity to synthesize essential oil through cytosolic aceate-mevalonate pathway.
PDF Abstract XML References Citation
How to cite this article
Deepak Ganjewala and Rajesh Luthra, 2007. Essential Oil Biosynthesis and Metabolism of Geranyl Aceate and Geraniol in Developing Cymbopogon flexuosus (Nees ex Steud) Wats Mutant cv. GRL 1 Leaf. American Journal of Plant Physiology, 2: 269-275.
DOI: 10.3923/ajpp.2007.269.275
URL: https://scialert.net/abstract/?doi=ajpp.2007.269.275
DOI: 10.3923/ajpp.2007.269.275
URL: https://scialert.net/abstract/?doi=ajpp.2007.269.275
INTRODUCTION
In vivio tracer studies of monoterpene biosynthesis have been extensively reviewed (Banthorpe et al., 1972, 1980; Charlwood and Banthorpe, 1978; Croteau, 1987; Singh et al., 1989b). Tracer studies indicated rapid metabolic turnover of monoterpenes in higher plants and the overall accumulation of monoterpenes in plants depends upon the balance between synthetic and catabolic processes (Croteau, 1988). Several labeled compounds such as CO2, glucose, sucrose, fructose, pyruvate, acetate and mevalonate have been used for such purpose in several aromatic plants (Francis and Allock, 1969; Francis and O’Connel, 1969; Luthra et al., 1993; Maffei et al., 2001). In lemongrass and citronella leaves [2-14C] acetate rather than [U-14C] sucrose was preferentially incorporated into monoterpenes (Singh and Luthra, 1988; Singh et al., 1990; Luthra et al., 1993).
However, in Mentha spicata fructose is reported to be a most uptaken sugar for (-)-carvone biosynthesis (Maffei et al., 2001). Essential oils in plants are biosynthesized through mevalonate- isoprenoid pathway or newly discovered non-mevalonate (pyruvate triose-phosphate) pathway (Rhomer et al., 1993; Lichtenthaler et al., 1997a; McCaskill and Croteau, 1998; Luthra et al., 1999). The major route of precursors and cofactors generation appears to be sugar-phosphate metabolism via glycolysis and pentose phosphate pathway (McKaskill and Croteau, 1998). Earlier studies with developing lemongrass (Cymbopogon flexuosus) leaves have revealed that metabolism of sucrose (Singh and Luthra, 1988) and mobilization of starch was most rapid in immature leaves, ensuring an efficient supply of carbon to the growing leaf for various biosynthetic purposes including terpenoid synthesis (Singh et al., 1990). Further, the monoterpene biosynthesis is associated with specialized anatomical structures such as trichomes, secretory cavities, idioblasts resin canals, latex vessels and epidermal glands (Gershenzon and Croteau, 1993). In Cymbopogon species only young and rapidly expanding leaves have the capacity to biosynthesize and accumulate essential oil in specific oil cells that are present in parenchymal tissues (Singh et al., 1990; Sangwan et al., 1993; Lewinsohn et al., 1998; Luhthra et al., 2007).
Cymbopogon flexuosus, commonly known as lemongrass is economically important because of its production of an essential oil which is widely used in flavours, fragrance and cosmetics for powerful rose and or lemon like smell. In geraniol rich lemongrass mutant cv. GRL-1, geraniol (89.39%) is the main monoterpene (Patra et al., 1997), which is not subjected to secondary transformations (oxidation) into geranial and neral, alike other lemongrass species and cultivars. The enzyme NADP-dehydrogenase, which oxidizes geraniol into geranial and neral seem to be metabolically blocked in mutant cv. GRL-1. Therefore, the present study was undertaken to investigate the essential oil biosynthesis and the metabolism of geranyl acetate and geraniol using [2-14C] acetate in developing GRL-1 leaf in view of the metabolic blockage between geraniol and geranial.
MATERIALS AND METHODS
Plant Material
Geraniol rich lemongrass (Cymbopogon flexuosus) mutant cv. GRL-1 plants were raised from slips at the experimental farm of Central Institute of Medicinal and Aromatic Plants (CIMAP), Lucknow during January-February 2001 and followed by standard agronomic practices.
Incorporation of [2-14C] Acetate into Essential Oil and its Major Constituents
For in vivo incorporation studies, geraniol rich mutant cv. GRL-1 leaves were collected from different developmental stages from 10 to 50 days. Leaves (six numbers) were transferred to test tubes and fed with an aqueous solution containing [2-14C]-acetate (activity 0.1 mCi, specific activity 34.51 mCi/mmole) of strength 5 μCi μmole-1 and kept in bright sunlight. Half-strength Hoagland solution (Hoagland and Arnon, 1938) were added successively so that the cut ends of the leaves remained immersed in the solution. After complete uptake of the labeled solutions, the vials were kept filled with half-strength Hoagland solution and after 24 h of incubation the leaves were removed, cut into small pieces, weighed and subjected to micro-scale steam distillation using mini Clevenger apparatus. The essential oil was recovered by ether extraction and treated with anhydrous sodium sulfate to remove the traces of moisture. Aliquots were used for the determination of total radioactivity in the essential oil. Thin layer chromatography was also performed in the aliquots, using toluene-ethyl acetate (96:4 v/v) on silica gel 60 F254, 20x20 cm (Merk) as described by Singh et al. (1990). Geranyl acetate and geraniol spots were scrapped off and subjected to radioactivity determination. The purity of the geraniol and geranyl acetate, separated through TLC, was checked by GLC.
Determination of Radioactivity Incorporated into Geraniol and Acetate Moiety of Geranyl Acetate
The [2-14C] acetate labeled geranyl acetate scrapped off was hydrolyzed with 10% ethanolic KOH in the flask fitted with a guard tube containing fused calcium chloride by incubation at room temperature for 24 h. After the incubation, 1 mL of water was added to the mixture and geraniol was recovered by ether extraction. Aqueous phase contained the acetate, which was neutralized by addition of equal moles of HCL. An aliquot each of the ether and aqueous phase containing geraniol and acetate moiety respectively, was taken in a scintillation vial for radioactivity counting.
Determination of Radioactivity
The radioactivity in the ether aliquots of essential oil and TLC isolated and hydrolyzed product of the geranyl acetate was analyzed by β-liquid scintillation counter (LKB Wallace 1409) using PPO-POPOP-toluene cocktail and cocktail W-dioxane. The counting efficiency of the instrument for [2-14C] acetate was 95%.
RESULTS AND DISCUSSION
Essential oil biogenesis at different developmental stages of leaf was assessed as the capacity to incorporate [2-14C] acetate into essential oil in vivo. Maximum [2-14C] acetate incorporation (pmol/10 leaves) into essential oil and its major constituents geranyl acetate and geraniol was observed at 24 h (Fig. 1). The incorporation of [2-14C] acetate into essential oil (pmol/10 leaves) was substantial during the initial growth stages (10 to 15 days) than declined significantly up to 68.0% by the end of leaf growth cycle (Table 1). Geranyl acetate has shown the similar [2-14C] acetate incorporation pattern to that of essential oil.
| Table 1: | Changes in biosynthetic rate of essential oil and its major constituents during leaf development. To determine the rate of essential oil biosynthesis, 15 days aysold leaves (six numbers) were incubated with 5 μCi μmole-1 aqueous solution of sodium [2-14C] acetate (activity 0.1 mCi, specific activity 34.51 mCi/mmole) for 24 h. Essential oil was extracted with diethyl ether and analyzed by liquid scintillation counter |
 | |
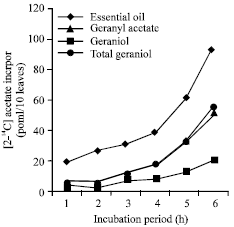 | |
| Fig. 1: | Time course of [2-14C] acetate incorporation. Fifteen days old (immature) leaves were fed with an aqueous solution of sodium [2-14C] acetate |
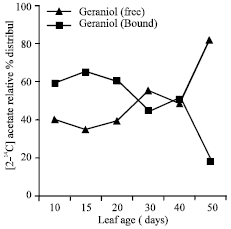 | |
| Fig. 2: | Radioactivity detected (relative %) in free and bound geraniol (as part of geranyl acetate) |
At 20 days, label detected into geranylc acetate was 89.99 (pmol/10 leaves), remained only 13.67 (pmol/10 leaves) by the end of leaf growth (50 days). In general, the label (pmol/10 leaves) detected into geraniol was lower as compared to that of geranyl acetate throughout the leaf development except at 50 days (Table 1). Biogenetic capacity of the leaves to synthesize total geraniol (Bound+free), expressed as sum of the label incorporated into free and bound geraniol (as part of geranyl acetate) was also substantial during early stages of leaf development (Table 1). The label (relative %) detected into bound geraniol decreased while increased correspondingly into free geraniol with leaf growth (Fig. 2). These results have shown that only immature mutant cv. GRL-1 leaves are biogenetically most active to synthesize essential oil through classical acetate-mevalonate pathway. The result presented are consistent to those reported earlier in C. flexuosus (Singh et al., 1989), C. martinii (palmarosa) (Dubey, 1999) and other species including Anethum graveolens (Porter et al., 1981), Salvia officinalis (Croteau et al., 1981), Carum caravi (Bouwmeester et al., 1998) and Mentha piperita (Gershenzon et al., 2000). In mutant cv. GRL-1 the relatively little incorporation into essential oil in the later stages of leaf growth may be attributed to low turnover of endogenous monoterpene pool. The saturation level of the biosynthetic sites for the substrate varies with age, system, precursor demand and quality of the end products exerting strain on the physiological status of the cell (Croteau, 1987; Singh et al., 1989, 1991). In Mentha, 14C-acetate and 14-C-palmitate was not incorporated into carvone rather fructose was found to be a most uptaken sugar suggesting the presence of an alternative non-mevalonate pathway for carvone biosynthesis (Maffei et al., 2001). Recently, Luan et al. (2005) studied the metabolism of deuterium labeled geraniol in Vitis vinifera and demonstrated the stereoselective reduction to (S)-citronellol, E/Z isomerization to nerol, oxidation to neral/geranial and glycosylation of the corresponding monoterpene alcohols which were dependent on the ripening stage.
Study of relative percent distribution of oil incorporated radioactivity into major oil constituents have shown that at 20 days of leaf growth label detected into geranyl acetate was 66.51% while in geraniol only 12.63% and trend was reversed with 49% in geraniol and 32% in geranyl acetate by the end of leaf growth phase (Fig. 3). Marked fluctuation in radioactivity incorporation into unknown constituent was observed during leaf growth from 10 to 50 days with significant (38.3%) value at 40 days (Fig. 3). The incorporation pattern of label into unknown constituent, which are mainly composed of monoterpene hydrocarbons (Thapa et al., 1981) exhibited the similar trend to those of earlier reported in C. flexuous (Sigh et al., 1990). Similar observations were also reported in C. khasianus, where the increase in the feeding time of the labeled precursor [2-14C] acetate resulted in the decrease in the label in citral with a corresponding increase in hydrocarbons and or/the unidentified products (Verma et al., 1985).
The ratio of label detected into geranyl acetate to geraniol (GA:G) at 20 days was highest (3.19) and significantly go down up to 0.65 at 50 days. Since geranyl acetate is the progenitor of geraniol, the ratio of GA to G could be influenced by the catalytic activity of enzymes mediating this transformation.
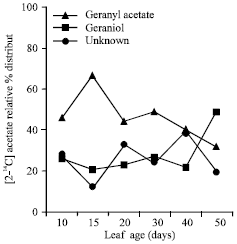 | |
| Fig. 3: | Relative percentage distribution pattern of radioactivity in major essential oil (geranyl acetate and geraniol) and unidentified constituents |
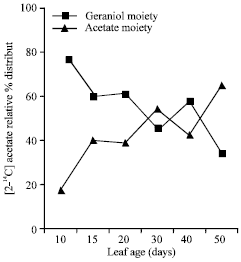 | |
| Fig. 4: | Radioactivity detected in geraniol and acetate moiety after hydrolyzing geranyl acetate with 10% ethanolic KOH |
Geranyl acetate esterase (GAE) of palmarosa (C. martinii) (Dubey and Luthra, 2001) and lemongrass (C. flexuosus) mutant cv. GRL-1 (unpublished) have been reported to influence the quality of essential oil by hydrolyzing geranyrl acetate into geraniol. The activity profile of GAE studied during lemongrass mutant cv. GRL-1 leaf development was found to be paralleled to that of the ratio of geranyl acetate to geraniol (unpublished). Therefore the decrease in ratio of GA:G is expected in mutant cv. GRL-1 leaf development. Earlier, Singh et al. (1990) in lemongrass reported the role of NADP specific geranyl dehydrogenase which influenced the geraniol to citral ratio by oxidizing geraniol into trans citral. In lemongrass mutant cv. GRL-1, alcohol dehydrogenase is metabolically blocked, hence it could not be measured.
Geranyl acetate was hydrolyzed using 10% ethanolic KOH to determine the radioactivity incorporated into geraniol and acetate moiety. The results have shown that during early stages of leaf growth label detected into geraniol was relatively very high (60-80%) which declined with corresponding increase in label detected into acetate moiety (Fig. 4). These results suggested that acetylation of geraniol was maximum during the initial stages of leaf development similar to those observed in palmarosa (Dubey, 1999).
In conclusion, lemongrass mutant cv. GRL-1 leaves biosynthesize and accumulates essential oil only during the initial growth phase while rapidly expanding. Substantial [2-14C] acetate incorporation suggested the role of cytosolic acetate-mevalonate pathway in formation of essential oil and its major constituents geranyl acetate and geraniol. Geranyl acetate rapidly metabolizes into geraniol during leaf development. Geraniol, however, could not undergo oxidation to geranial due to metabolic block in NADP specific alcohol dehydrogenase. However, it is now known that, under most circumstances, the monoterpenes are not formed by the classical acetate-mevalonate pathway, but rather by the glyceraldehydes-3-phosphate/pyruvate pathway. Therefore, studies on the specifically labeled glucose into terpenoid compounds is being undertaken in mutant cv. GRL-1, in distinguishing the operation of the traditional acetate-mevalonate pathway and the glyceraldehydes-3-phosphate pathway in the formation of the monoterpenes.
ACKNOWLEDGMENT
The authors are grateful to Dr. Sushil Kumar, former director, CIMAP, Lucknow, India, for providing facilities.
REFERENCES
- Banthorpe, D.V., B.V. Charlwood and M.J.O. Francis, 1972. The biosynthesis of monoterpenes. Chem. Rev., 72: 115-153.
CrossRef - Bouwmeester, H.J., J. Gershenzon, M.C.J.M. Konings and R. Croteau, 1998. Biosynthesis of the monoterpenes limonene and carvone in the fruit of caraway: I. Demonstration of enzyme activities and changes with development. Plant Physiol., 117: 901-912.
Direct Link - Croteau, R., M. Flton, F. Karp and R. Kjonas, 1981. Relationship of camphor biosynthesis o leaf development in sage (Salvia officinalis). Plant Physiol., 67: 820-824.
Direct Link - Dubey, V.S. and R. Luthra, 2001. Biotranformation of geranyl acetate to geraniol during palmarosa (Cymbopogon martinii Roxb. Wats var. motia) inflorescence development. Phytochemistry, 57: 675-680.
PubMedDirect Link - Francis, M.J.O. and M. O'Connel, 1969. The incorporation of mevalonic acid into rose petal monoterpenes. Phytochemistry, 8: 1705-1708.
CrossRef - Gershenzon, J., M.E. McConkey and R. Croteau, 2000. Regulation of monoterpene accumulation in leaves of peppermint. Plant Physiol., 122: 205-213.
Direct Link - Hoagland, D.R. and D.I. Arnon, 1938. The water culture method for growing plants without soil. Calif. Agric. Exp. Station, C347: 1-39.
Direct Link - Lewinsohn, E., N. Dudai, Y. Tadmor, I. Katzir, U. Ravid, E. Putievsky and D.M. Joel, 1998. Histochemical localization of citral accumulation in lemongrass leaves (Cymbopogon citratus (DC.) Stapf., Poaceae). Ann. Bot., 81: 35-39.
Direct Link - Lichtenthaler, H.K., M. Rohmer and J. Schwender, 1997. Two independent biochemical pathways for isopentenyl diphosphate and isoprenoid biosynthesis in higher plants. Physiol. Plant., 101: 643-652.
Direct Link - Luan, F., A. Mosandl A. Munch and M. Wust, 2005. Metabolism of geraniol in grape berry mesocarp of Vitis vinifera L. cv. Scheurebe: Demonstration of stereoselective reduction, E/Z-isomerization, oxidation and glycosylation. Phytochemistry, 66: 295-303.
Direct Link - Luthra, R., P.M. Luthra and S. Kumar, 1999. Redefined role of mevalonate-isoprenoid pathway in terpenoid biosynthesis in higher plants. Curr. Sci., 76: 133-135.
Direct Link - Maffei, M., W. Camusso and D. Caramellino, 2001. Fructose is the most uptaken sugar for (-)-carvone biosynthesis in Mentha spicata (L). J. Plant Physiol., 158: 811-813.
CrossRefDirect Link - McKaskill, D. and R. Croteau, 1998. Some caevets for bioengineering terpenoid metabolism in plants. Trends Biotechnol., 16: 349-355.
Direct Link - Rhomer, M., M. Knani, P. Simonin, Sutter and H. Sahm, 1993. Isoprenoid biosynthesis in bacteria: A novel pathway for early steps leading to isopentenyl diphosphate. Biochem. J., 295: 517-524.
Direct Link