
Nishi Mathur
Microbial Biotechnology and Biofertilizer Laboratory, Department of Botany, J.N.V. University, Jodhpur-342003 Raj, India
Anil Vyas
Microbial Biotechnology and Biofertilizer Laboratory, Department of Botany, J.N.V. University, Jodhpur-342003 Raj, India
American Journal of Plant Physiology
Year: 2007 | Volume: 2 | Issue: 2 | Page No.: 122-138







ABSTRACT
The study of Arbuscular Mycorrhizal (AM) fungi and the AM symbiosis formed with host plant roots is complicated by the biotrophic and hypogeous nature of the mycobionts involved. To overcome this, several attempts have been made during the last three decades to obtain this symbiosis in vitro. The use of root-organ cultures has proved particularly successful. In this review, we describe the method by which root-organ cultures (transformed and nontransformed) have been obtained, together with the choice of host species, inoculation techniques and culture media. This is supported by a summary of some of the most important findings, regarding this symbiosis, that have been made at the physiological, biochemical and molecular levels.
PDF Abstract XML References Citation
How to cite this article
Nishi Mathur and Anil Vyas, 2007. Arbuscular Mycorrhiza on Root-Organ Cultures
. American Journal of Plant Physiology, 2: 122-138.
DOI: 10.3923/ajpp.2007.122.138
URL: https://scialert.net/abstract/?doi=ajpp.2007.122.138
DOI: 10.3923/ajpp.2007.122.138
URL: https://scialert.net/abstract/?doi=ajpp.2007.122.138
INTRODUCTION
The establishment of in vitro root-organ cultures has greatly influenced our understanding of the Arbuscular Mycorrhizal (AM) symbiosis. Because of its potential for research and inoculum production, we outline a full description of the culture methods and a summary of the important findings that have resulted from the use of this in vitro system.
Mycorrhizal Root-organ Culture Methods
Host Roots
Root-organ cultures were first developed by White (1943), Butcher and Street (1964) and Butcher (1980). These authors used excised roots on synthetic mineral media supplemented with vitamins and a carbohydrate source. However, profuse root growth, characterized by the formation of numerous lower order branches, has been obtained with relatively few plant species. The formation of lower order roots is essential for rapid increase in root biomass and the establishment of continuous cultures.
Pioneering work by Mosse and Hepper (1975) used root cultures obtained from Lycopersicum esculentum Mill. (tomato) and Trifolium pratense L. (red clover) to establish in vitro mycorrhiza with Glomus mosseae Nicolson and Gerd. The authors demonstrated for the first time that spores of an AM fungus could be successfully used to colonize excised roots growing on a mineral-based medium. Later, Strullu and Romand (1986, 1987) showed that it was also possible to reestablish mycorrhiza on excised roots of Fragaria xAnanassa Duchesne (strawberry), Allium cepa L. (onion) and tomato, using the intraradical phase (i.e., vesicles or entire mycorrhizal root pieces) of several species of Glomus as inoculum.
A natural genetic transformation of plants by the ubiquitous soil bacterium Agrobacterium rhizogenes Conn. (Riker et al., 1930) produces a condition known as hairy roots. This stable transfonnation (Tepfer, 1989) produces Ri T-DNA transformed plant tissues that are morphogenetically programmed to develop as roots. Their modified hormonal balance makes them particularly vigorous and allows profuse growth on artificial media (Tepfer, 1989).
Daucus carota L. (carrot) and Convolvulus sepium L. (bindweed) were among the earliest species to be transformed using Agrobacterium rhizogenes Conn (Tepfer and Tempe 1981). These Ri T-DNA transformed roots have since served in a wide range of fundamental and applied studies. One of the most important of these has been the study of the AM symbiosis. The first culture of hairy roots colonized by an AM fungus was achieved by Mugnier and Mosse (1987), successfully colonized Convolvulus sepium hairy roots using spores of G. mosseae but, as was the case with nontransformed clover root-organ cultures (Mosse and Hepper, 1975), no sporulation occurred. Spore production followed reductions in the concentration of certain nutrients in the culture medium (Becard and Piche, 1990) that allowed mycorrhizal inhibition to be avoided, but did not affect root growth and development. This led to the production of reproducible monoxenic cultures of G. intra radices that were characterized by large quantities of mycelium and spores (Diop et al., 1992).
Low mineral media minimal (M) and modified Strullu-Romand (MSR) media (Table 1) were also successfully used to obtain mycorrhiza and fungal sporulation using nontransformed tomato root cultures (Chabot et al., 1992; Diop et al., 1994a, b). Nevertheless, transformed roots have a greater growth potential, which makes them more adaptable to different experimental conditions and they can be generated from most dicotyledonous plants (Tepfer, 1989). However, rigorous comparisons between transformed and nontransformed root cultures have never been made. Such studies should ideally be done using roots from the same plant material.
Whichever type of root system is chosen, success in establishing a mycorrhizal culture depends on the physiological state of the host root. Roots from the same clone, grown under the same conditions, can behave differently. Subculture frequency, explant selection and orientation of the Petri dishes during incubation (e.g., horizontal, upside down, or vertical) are important culture parameters that must be optimized for each clone.
Fungal Inocula
In most cases, two types of fungal inoculum can be used to initiate monoxenic cultures: either extraradical spores or propagules from the intraradical phase (i.e., mycorrhizal root fragments and isolated vesicles) of the fungus. However, cultures of AM fungal species that do not produce vesicles (e.g., Scutellospora and Gigaspora species) are systematically produced using spores, which are usually large and germinate vigorously. Sporocarps of G. mosseae have also been used in an attempt to establish in vitro cultures (Budi et al., 1999).
Spores
Spores are usually collected from the field, or from pot cultures, by wet sieving. With small spore samples (tens or hundreds), spores can be chosen individually under a dissecting microscope using a micropipette or fine tweezers. However, with larger spore samples gradient centrifugation must be used to separate out spores. Several centrifugation methods, based on the use of various highly concentrated substances (e.g., sucrose, glycerol, Percoll and Radiopoaque contrast media), have been successfully used (Mertz et al., 1979; Furlan et al., 1980; Hosny et al., 1996). It is important, however, that spores are not subjected to prolonged exposure to these substances.
Before being used as in vitro inoculum, spores must be surface sterilized (Becard and Piche, 1990). This step is critical because success depends on the elimination of all contaminants. It should be noted, however, that in some cases spores may carry bacteria between wall layers, making disinfection difficult or even impossible (Walley and Germida, 1996).
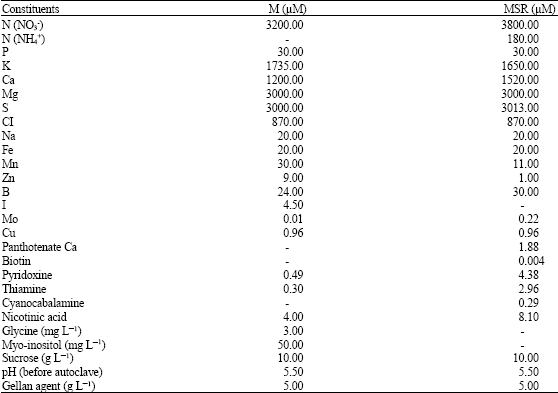
| Table 1: | Comparative composition of minimal (M) and modified Strullu-Romand (MSR) media |
 | |
A solution containing the strong oxidizing agent, chloramine T and a surfactant (e.g., Tween 20) is widely used to sterilize AM fungal spores. Although 20 min in a 2% solution usually gives satisfactory results, concentration and treatment duration can be adapted depending on contaminant levels and spore sensitivity. Ideally, spores should be gently agitated during sterilization, or a vacuum applied to degas the spore surface. Spores are subsequently rinsed in a streptomycin-gentamycin antibiotic solution (Becard and Piche, 1992). To maintain spore dormancy, all steps from spore isolation to rinsing should be done on ice. If spores are not to be used immediately, they should be stored at 4°C, either in distilled water, on water agar, or on 0.1% MgSO4.7H2O solidified with 0.4% gellan gum. To reduce the risk of contamination by bacteria or fungi that were not eliminated during the sterilization process, spore number should be limited in each Petri dish.
Generally, AM fungal spores do not need specific conditions or the presence of a host root to germinate. However, root exudates and 2% CO2 can stimulate germination and (or) postgermination hyphal growth (Becard and Piche, 1989; Buee, 2000). Recalcitrant spores can be placed alongside a growing root. If spores fail to germinate within 20 days, either the sterilization treatment was too strong or the spores were immature, dormant, or dead. It is well known that spores of some AM fungal species require cold stratification (4°C) prior to germination (Smith and Read, 1997). This requirement can vary within a genus: Gigaspora gigantea (Nicolson and Gerd.) Gerd. And Trappe (Koske, 1981) and Gigaspora margarita Becker and Hall require a cold treatment, whereas Gigaspora rosea Nicolson and Schenck (formally misidentified as Gigaspora margarita (Bago et al., 1998a). The cold treatment (14-21 days) is best applied prior to spore isolation, when the spores are still attached to the extraradical mycelium.
Mycorrhizal Root Fragments
In general, mycorrhizal roots used to initiate monoxenic cultures come from trap plants grown in pot cultures, with field-collected soil or AM fungal propagules. Leek (Allium porrum L.) plants are widely used because of their high susceptibility to colonization. Young, healthy, translucent leek roots should be chosen for in vitro culture establishment, as sections of roots with vesicles can be easily located. The roots are then disinfected in an ultrasonic processor under a laminar-flow hood. Treatment duration and reagent concentrations can be adapted to specific situations (e.g., host plant, root age and contamination level).
Disinfected roots are cut into 510 mm lengths and incubated on a synthetic medium. Water-agar medium is also effective (Diop et al., 1994a). Petri dishes should be incubated in the dark at 27°C. Hyphal regrowth from root pieces is usually observed within 2-15 days.
As shown for spores (mentioned earlier), there is no evidence that mycorrhizal root pieces need specific exogenous conditions or a host plant for hyphal regrowth (Diop et al., 1994a). Following incubation, mycorrhizal root pieces showing hyphal regrowth are transferred, using a cork borer, to a fresh Petri dish with an actively growing root or an actively growing root is transferred to the Petri dish containing the mycorrhizal root.
Although field-collected roots have never been directly used as starter inoculum for in vitro cultures, their use should not be excluded. Vesicles within roots may be less contaminated than the root surface, offering a better source of inoculum. The vesicle extraction method (Strullu and Romand, 1987; Strullu and Plenchette, 1991) could be useful for such inocula. Vesicles, enzymatically extracted from roots, have been used to establish cultures with G. intraradices, Glomus versiforme (Karsten) Berch and Glomus macrocarpum Tulasne and Tulasne (Strullu and Romand, 1986, 1987), but vesicles are rarely used for routine inoculation. Comparisons have been made between the use of in vitro produced spores and vesicles isolated from leek plants grown in pot culture (Nantais, 1997). Briefly, for a given number of propagules, root colonization was more efficient when using spores than when using isolated vesicles.
Culture Media
The ingredients of the two most widely used and equally successful media for in vitro mycorrhizal root cultures are listed in Table 1. The M medium is a modified White’s medium initially developed for tomato root-organ cultures (Butcher, 1980). The macroelement composition of White’s medium is considerably lower than that of MS and B5 media, commonly used for in vitro plant cultures (Becard and Piche, 1990). However, this dilute medium is adequate for root growth. The composition of M medium is even poorer and was developed following a bioassay that compared the effects of different element concentrations on mycorrhiza formation.
The MSR medium is a modified A medium, which was developed to optimize the growth of the intraradical phase of the fungus in vitro. The macroelement composition of MSR is similar to that of the M medium. Differences between the two media occur in oligoelement and vitamin concentrations: MSR medium lacks iodide, myo-inositol and glycine and M medium lacks panthotenate, biotin and cyanocobalamine. These various components are perhaps not essential, since their absence in either medium has no apparent negative effect on the AM symbiosis.
Both media are adjusted to pH 5.5 before autoclaving and are solidified with gellan gum. Almost 30 AM fungal isolates from the Acaulosporaceae, Gigasporaceae and Glomaceae are now successfully grown on these media (Table 2). However, as the compositions of the M and MSR media were established empirically, they could probably be further optimized. It is conceivable, for example, that AM fungi isolated from acidic or alkaline soils require either lower or higher pH, respectively, in vitro.
Glomales in vitro Collection
Since 1975, when Mosse and Hepper (1975) first grew the mycelium of G. mosseae using an in vitro system, at least 27 AM fungal species have been successfully cultivated on root-organ cultures. The majority of these have been obtained during the last decade (Table 2).
| Table 2: | Species of Glomales cultivated on root-organ cultures |
 | |
| aSpecies differentiating only vesicle-like spores | |
The increasing number of species of AM fungi cultivated in vitro and the possibility of continuous cultivation and cryopreservation, has led to the development of an international collection of in vitro AM fungi: the Glomales in vitro collection (GINCO). This collection has resulted from a collaboration between the Mycotheque de l’Universite Catholique de Louvain (MUCL, Belgium) and the Eastern Cereal and Oilseed Research Centre (ECORC, Agriculture and Agri-Food Canada), which is responsible for the Canadian Collection of Fungal Cultures (CCFC/DAOM, Canada). GINCO aims to conserve biodiversity and provide high-quality, contaminant-free AM fungal inocula for scientific research. GINCO, in collaboration with a team of scientists working on AM fungal physiology, biochemistry; taxonomy and ecology, intends to increase the number of taxa available, offer specialized training and develop an international network of collaborative research.
Fungal Morphological Features Before and after Root Colonization
Previously, most structural studies concentrated on the intraradical plant-fungus interfaces and relatively few studies investigated the structural aspects of the precolonization and extraradical phases. However, in vitro cultivation of AM fungi using root-organ cultures opens new avenues for hyphal structural studies during spore germination, precolonization and development of the extraradical mycelium.
In vitro Germination of Spores
The use of AM root-organ cultures allows the aseptic production of spores of various AM fungal species. Although it is well known that cold stratification is important to break the inherent dormancy-like stage found in certain AM fungal species, recent observations showed that this treatment not only affects spore germinability but also has a dramatic effect on germ tube morphology. Cold treatment applied for more than 14 days led to full germination with strong apical dominance and sparse branching as previously described by Mosse (1988). In the absence of a cold treatment, a unique germination pattern was observed: germ tubes were short with profuse branching, spiraling around and close to the spores.
Although AM fungi have the capacity for initial germination, germ tube elongation is fatally blocked in the absence of a host plant (Bonfante and Perotto, 1995). Recently, in vitro studies using two-photon microscopy revealed autolytic zones in live but senescent germ tubes of Gi. rosea (Bago et al., 1998b). These areas coexisted with zones exhibiting complete cytoplasmic integrity. Cytological analyses suggested that portions of these coenocytic hyphae were undergoing cell death or apoptotic processes (programmed death). This precolonization senescence phenomenon is theoretically reversible.
In vitro Development of the Extraradical Phase
The use of root-organ cultures in compartmentalized Petri dishes (St-Amaud et al., 1995) also allows time-lapse studies of extraradical mycelial development in root-free compartments. When comparing ammonium and nitrate as nitrogen sources, it was found that the presence of ammonium in the distal compartment drastically reduced spore production. This also suggests that in the presence of ammonium, the mycelium of the extraradical phase developed coiled hyphae and hyphal aggregations that were never observed in the presence of nitrate. This in vitro system allowed to observe the structural development of the extraradical phase of G. intraradices, which comprises an organized radial network of runner hyphae from which lower order branches (at a 45° angle) develop at regular intervals (between 25 and 300 μm). Some of these ramifications developed into new runner hyphae and others bore arbuscule-like structures (ALS) and spores. Ultrastructural investigations revealed that ALS (renamed branched absorbing structures or BAS) are very similar to intraradical arbuscules and that, like arbuscules, they are sites of intense metabolic activity. Arbuscules and BAS are also similar in terms of their gross morphology (thinner diameter with in-creased dichotomous branching). The extent to which these structures are functionally comparable remains to be elucidated. However, prolific branching of the fungus to form BAS results in an important increase in surface area and so produces a structure better adapted for nutrient uptake. It has also been shown that increased acidification of the medium coincides with a higher production of spore-associated BAS. This change in pH could be a direct consequence of a greater phosphate uptake, to provide storage products for the spores. It also appears that inorganic nitrogen and phosphate absorption by extraradical mycelium is closely correlated with BAS development.
BAS and arbuscules also have similar life-spans (approx. 7 days). However, the reason for such short life-spans and the evidence for possible host involvement are generally lacking (Smith and Read, 1997). Apoptotic processes within these structures may explain the prompt degradation observed. This hypothesis is supported by the early events leading to nuclear degradation observed within lysed compartments in extraradical hyphae (Bago et al., 1999).
MYCORRHIZATION OF THE MICRO PROPAGATED PLANTLETS-A CASE STUDY
Survival and Establishment of in vitro Raised Ziziphus Nummularia and Ziziphus Mauritiana by Application of Am Fungi
Glomus deserticola Trappe, Bloss and Menge and Glomus constrictum Trappe, are one of the most commonly occurring VAM fungi of arid and semiarid regions, were cultured and multiplied in root organ cultures of Z. nummularia and Z. mauritiana under in vitro conditions. The in vitro produced VAM fungi established efficient symbiosis with in vitro raised plantlets of Z. nummularia and Ziziphus mauritiana. This VAM strain improved the biomass production, nutrient uptake and acclimatization of the in vitro produced plantlets of Z. nummularia and Ziziphus mauritiana in pots under green house conditions.
Introduction
Z. nummularia and Z. mauritiana are an important multipurpose fruit plants of arid and semiarid regions. These plants are used as a source of fuel, fodder and food. The cultivation is done mostly in nutrient deficient sandy soils of drought prone areas. Rapid in vitro multiplication of this plant has successfully been done using tissue culture techniques (Mathur et al., 1993).
However the major handicap of tissue culturists in arid and semi-arid regions is survival and establishment of in vitro raised plantlets into fields (Mathur and Vyas, 1995a). Arid and semi-arid regions are characterised by high temperature, water scarcity and nutrient deficient sandy soils. All of these factors combined contribute towards harsh environmental conditions, due to the fact that the primary stresses imposed on vegetation by arid environments are lack of water and mineral nutrients (Fisher and Turner, 1978). Therefore, any factor which enhances nutrient uptake and water transport is likely to contribute to the success of desert vegetation. VAM symbiosis may be of particular significance in coping with P-deficiency stress in natural ecosystems (Mc Arthur and Knowles, 1993).
Due to its beneficial effect VA mycorrhizae are receiving considerable attention in agriculture and forestry (Peterson et al., 1984). In vitro production of different species of Glomus, viz., G. fasciculatum (Allen et al., 1982). G. intraradices (Chabot et al., 1992), G. mosseae (Nopamornbodi et al., 1988), either axenically or in root organ culture using various host plants has been reported. However, there are no reports about the culturing of G. deserticola and G. constrictum (most commonly occurring VAM species of arid and semiarid regions) under in vitro conditions. Hence, the present investigation was undertaken to produce G. deserticola and G. constrictum under in vitro conditions in association with Z. nummularia and Z. mauritiana.
MATERIALS AND METHODS
The plantlets of Z. nummularia and Z. mauritiana were raised from different explant source under in vitro conditions on modified MS medium containing various concentrations and combinations of auxins and cytokinins (Mathur et al., 1993). VA mycorrhizal fungi G. deserticola Trappe, Bloss and Menge and G. constrictum Trappe, collected from rhizosphere soils of Z. nummularia and Z. mauritiana, were cultured on Cenchrus ciliaris for establishing pot cultures following Mathur and Vyas (1996). Roots of C. ciliaris from these pot cultures were regularly examined to test purity of culture. The pure pot culture of C. ciliaris containing spores of only G. deserticola were used for inoculating Z. nummularia and only G. constrictum were used for inoculating Z. mauritiana. The spores of G. deserticola and G. constrictum along with the infected root segments of C. ciliaris were isolated from soils of the pot cultures using the wet sieving and decanting technique (Gerdemann and Nicolson, 1963). The spores were then purified from root pieces and debris by sucrose gradient centrifugation (Jenkins, 1964). The collected spores were washed repeatedly with sterilized distilled water to remove sucrose. These purified spores along with the infected root segments were surface sterilized with sodium hypochlorite (2.5% v/v) for 20 min and washed repeatedly with sterilized distilled water to remove traces of sodium hypochlorite. Because the infected root segments were fully colonized by active spores of G. deserticola and G. constrictum, respectively, these root segments were also used for inoculation. Hence, the spores of G. deserticola and G. constrictum along with C. ciliaris root segments were used to inoculate in vitro raised plantlets of Z. nummularia and Z. mauritiana, respectively.
At the time of rooting of In vitro raised plantlets, 10-15 spores of G. deserticola and G. constrictum along with infected root segments of C. ciliaris were inoculated onto roots of Z. nummularia and Z. mauritiana respectively in 250 mL flask. At the same time a control was maintained by inoculating roots of Z. nummularia and Z. mauritiana with sterilized root segments of C. ciliaris without any VAM spores. These flasks were kept under controlled conditions having 60% humidity and 20-25°C temperature. After 40 days the plantlets were transferred into pots of 18 cm diameter containing sterilized sand:vermiculite in 3:1 ratio (Mathur et al., 1993) and were kept under green house conditions. The samples were harvested 90, 120, 150 and 180 days after inoculation. Plant dry weight were recorded after drying them in a hot air oven at 60°C for 48 h. Root samples collected from the pots were gently washed under tap water and suitably processed (Phillips and Hayman, 1970) to calculate the percentage of root colonization (Giovannetti and Mosse, 1980). Phosphorus in plant material was estimated by Vanadomolybdate method (Jackson, 1973) and nitrogen in plant material was estimated by micro-Kjeldhal method. There were twenty-five replicates for each treatment. The control plants were not supplied with VAM inoculum.
RESULTS AND DISCUSSION
The inoculation of Z. nummularia plantlets with spores of G. deserticola and infected root segments of C. ciliaris resulted in establishment of a symbiotic relationship. The VAM fungi proliferated vigorously under in vitro conditions and produced all the structures of VAM infection, viz. external and internal hyphae, vesicles and arbuscules. The influence of in vitro produced VAM fungi was evaluated for nutrient uptake and biomass production in the in vitro produced plantlets of Z. nummularia (Table 3). Overall growth of mycorrhizal in vitro raised plantlets was higher than non-mycorrhizal plantlets after all harvest. However, the samples of last harvest showed more than 2-fold increase in plant height and almost 3-fold increase in plant dry weight when inoculated with VAM fungi. Similar trend was also noticed in effect of VAM fungi on nutrient uptake in in vitro raised plantlets of Z. nummularia. Addition of VA mycorrhiza resulted in more than 140% increase in uptake of both the nutrients P and N, compared with the control (Table 4). This result likely is due to the vigorous proliferation and efficient colonization of the roots in vitro raised plantlets of Z. nummularia by G. deserticola (Table 3).
The present investigation reveals successful symbiosis of VA mycorrhizal fungus Glomus deserticola in association with Z. nummularia under in vitro conditions. Mosse and Hepper (1975) were first to produce a simplified in vitro system for the study of VAM. Strullu and Romand (1986) reported regeneration of vigorous hyphae of three Glomus species from sterilized root fragments. During the present investigation, culturing of VAM fungi was achieved on modified MS medium, which is also the best medium for micropropagation of the host plant Z. nummularia (Mathur et al., 1993). This is of importance because both the plantlets as well as the VAM fungi can be propagated simultaneously on the same medium under in vitro conditions.
| Table 3: | VAM spore population and percentage of root colonization in rhizosphere of in vitro raised Z. nummularia plantlets at different harvests |
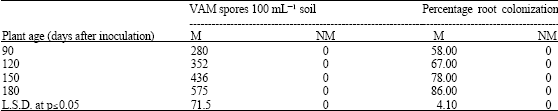 | |
| M = Mycorrhizal inoculated, NM = Non-mycorrhizal inoculated | |
| Table 4: | Biomass production and nutrient uptake in vitro raised plantlets of Z. nummularia by in vitro produced G. deserticola at different harvests |
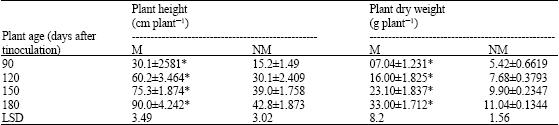 | |
| M = Mycorrhizal inoculated, NM = Non-mycorrhizal inoculated, ANOVA test * Significant at 0.1% level (at each harvest) LSD at px0.5, comparing M vs NM | |
| Table 4: | Continued |
 | |
| M = Mycorrhizal inoculated, NM = Non-mycorrhizal inoculated, ANOVA test * Significant at 0.1% level (at each harvest) LSD at px0.5, comparing M vs NM | |
The in vitro produced VAM fungi also were found to be significantly beneficial for biomass production and nutrient uptake of in vitro raised Z. nummularia plantlets in pots. The increased nutrient uptake by VAM endophytes has been well recognised (Koide end Schriener, 1992; McArthur and Knowles, 1993). Thus, the increased plant biomass which can be attained by increased nutrient levels was observed during the present investigation.
The inoculation of in vitro produced Z. mauritiana plant- lets with spores and infected root segments of G. constrictum resulted in establishment of symbiotic relationship. The VAM fungi proliferated vigrously under in vitro conditions by germination of spores. These germinating spores then penetrated roots of Z. mauritiana and proliferated vigrously in cortical region and formed vesicles and arbuscules. The potentiality of in vitro produced VAM fungi was evaluated towards nutrient uptake, biomass production and establishment of in vitro produced plantlets of Z. mauritiana. Table 5 represents observations regarding plant biomass production and nutrient uptake in mycorrhizal and non-mycorrhizal Z. mauritiana plantlets. Overall growth of mycorrhizal in vitro raised plantlets was higher than non-mycorrhizal ones at all of the harvests. However, the samples of the last harvest showed a more than two-fold increase in plant height and plant dry weight when inoculated with VAM fungi. A similar trend was also noticed as an effect of VAM fungi on nutrient uptake in vitro raised plantlets of Z. mauritiana. Addition of VAM increased more than two-fold the uptake of both of the nutrients, i.e. N and P (Table 5), by vigrous proliferation and efficiently colonizing the roots of in vitro raised plantlets at a greater rate (Table 6), ultimately leading to better survival of plantlets into the field (Table 7). The present investigation reveals successful sporulation and proliferation of G. constrictum under in vitro conditions in association with Z. mauritiana on modified MS medium. First contact between the root and fungus occurred within 2 days, possibly due to the use of VAM root segments in addition to spores as source of inoculum. After 15 days of growth, the medium surface was covered with a network of hyphae of VA mycorrhiza. Many stages of the vegetative life cycle of G. constrictum, including numerous arbuscules and vesicles and extramatrical mycelium, were easily observed in this in vitro dual culture system.
| Table 5: | Biomass production and nutrient uptake in in vitro raised mycorrhizal plantlets and non-mycorrhizal plantlets of Z. mauritiana |
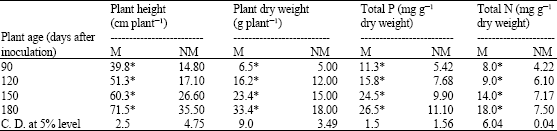 | |
| ANOVA test * Significant at 0.1% level (at each harvest); M = Mycorrhizal, NM = Non-mycorrhizal | |
| Table 6: | VAM spore population and percentage of root colonization in vitro raised mycorrhizal and non-mycorrhizal of Z. mauritiana plantlets |
 | |
| M = Mycorrhizal inoculated, NM = Non-mycorrhizal inoculated | |
| Table 7: | Rate of survival and RFMD of in vitro raised Z. mauritiana plantlets into the field |
 | |
| M = Mycorrhizal, NM = Non-mycorrhizal. RFMD = Relative Field Mycorrhizal Dependency | |
The in vitro sporulated G. constrictum significantly increased p-levels in Z. mauritiana plantlets by efficiently colonizing the roots. The increased p-uptake by VAM endophytes has been well recognized (Koide and Schriener, 1992; Ruiz Lozano et al., 1995). An improved net photosynthetic rate of Z. mauritiana by V A mycorrhizae has been reported under Ex vitro conditions (Mathur and Vyas, 1995b). Hence, improved biomass production of Z. mauritiana during the present study was observed, which could be attributed to improved nutrient uptake and possibly improved photosynthetic rate. The increased nitrogen content in mycorrhizal plants could be due to increased nitrate reductase activity which was attributed to improved p-nutrition provided by a VAM symbiosis (Cliquet and Stewart, 1993; Mathur and Vyas, 1995c). The present study revealed direct correlation between VAM spore populations and percentage of root colonization. These strains that efficiently colonized roots were found to produce a maximum number of spores.
Both the mycorrhizal and non-mycorrhizal plants survived in the field when transferred. However, rate of survival was quite different. During the first harvest 100% survival was observed for both the mycorrhizal and non-mycorrhizal plants, while the samples of the last harvest showed only 45% survival of non-mycorrhizal plants and 92% survival of mycorrhizal plants (Table 7). One of the major impediments to the success of micro-propagation is the very high mortality rate of tissue culture plantlets either during the acclimatization phase or during transfer to field conditions.
This is attributed mainly to certain aberrant features, characteristic of in vitro derived plantlets, which often lead to very high mortality due to desiccation and microbial infection. VA mycorrhizae are important in sustainable agriculture because they improve plant water relations and thus increase drought resistance of the host plant (Mathur and Vyas, 2000), improve disease control (Azcon-Augilar et al., 2002) and increase mineral uptake. All of these benefits provided by the VAM endophyte could be leading towards significantly improved survival of in vitro raised plantlets of Z. mauritiana into the field. Inoculation of micro-propagated plantlets of Z. mauritiana with active cultures of VAM appears to be critical for the survival and growth of in vitro plantlets; this avoids transient transplant shock and stunted growth upon transfer to the field (Lovato et al., 1995).
Hence the present investigation clearly suggests In vitro multiplication of VAM fungi G. constrictum in association with Z. mauritiana. The investigation further proves potentiality of this in vitro proliferated VAM strain on biomass production, nutrient uptake and survival of in vitro raised Z. mauritiana plantlets into the field. This work can be of importance in acclimatizing the in vitro raised plantlets of arid and semi-arid regions into the field, which may help in over-coming the major handicap of tissue culturists of arid areas.
Biological studies of VAM fungi are made difficult by the obligately biotropic relationship they form with vascular plants. The root organ culture technique for VAM fungi has obvious advantages over traditional systems permitting the observation of fungal morphology and development in vitro. Because the spores produced in this culture system are viable and able to colonize new roots, it also permits the propagation of VAM fungi monoaxenically. Thus, the present investigation may be of significance for acclimatizing in vitro raised plantlets of arid and semiarid regions into the field.
Limitations and Potentials for Using the Mycorrhizal Root-organ Culture System
Although the use of mycorrhizal root-organ cultures has allowed the elucidation of many aspects of the AM symbiosis, the in vitro system has obvious limitations. Perhaps one of the most important of these is the fact that the plant host is replaced by a root organ. As a result, the symbiotic benefit to the plant is affected by the absence of photosynthetic tissues, a normal hormonal balance and physiological source-sink relationships. Sucrose is added to the culture medium to compensate for the absence of photosynthates. Therefore, the root-fungus interface is bathed in a sugar solution, which does not occur in vivo. In this case, carbohydrates reach the cortex and the vascular system via the epidermis. It is possible that the presence of sugars at this interface modifies the biochemistry of the plant-fungal interaction. This might explain why arbuscules and vesicles are often scarce in Ri T-DNA transformed carrot roots, despite abundant intracortical mycelium. However, this hypothesis is not supported by recent work with M. truncatula hairy roots inoculated with G. intraradices, which exhibit colonization levels of up to 40%, this being mostly arbuscular.
Despite the artificial nature of this in vitro system, there are several legitimate reasons for its continued use in the study of AM fungi. The fungus forms typical colonization structures (i.e., appressoria, arbuscules and vesicles) and produces profuse extraradical mycelium and spores. The production of spores, morphologically and structurally similar to those produced in pot cultures and of intraradical structures capable of initiating new mycorrhizal symbiosis following subculturing indicates that the fungus is able to complete its life cycle. It can, therefore, be assumed that the mechanisms controlling the early colonization steps reflect those occurring in vivo.
This in vitro system has proved to be a useful tool for the cultivation and conservation of a large number of species and isolates of AM fungi (Table 2). It has also allowed many taxonomically important observations. It is likely that methodological improvements will help to establish cultures of some of the more recalcitrant Glomales species. To achieve this, media composition and growth conditions could be optimized. Studies suggest that mycorrhizal roots release com-pounds that are inhibitory for mycelial development and spore production (St-Amaud et al., 1995). Although species such as G. intra radices can survive under the in vitro environment presently used, it is possible that some species or isolates cannot. Knowing the nature of these inhibitors would perhaps permit their elimination, removal, or sequestration, which might help the cultivation of more recalcitrant species.
The validity of the continued use of root-organ cultures for studying some of the most challenging questions regarding biochemical, genetical and physiological relationships between AM fungi and their hosts is supported by the fact that these tissues show the same mycorrhizal characteristics as the plants from which they were developed. For example, hairy roots from non-host plants (e.g., Beta vulgaris L., Brassica nigra (L.) Koch and Brassica kaber (DC) L.C. Wheeler (Becard and Piche, 1990; Schreiner and Koide, 1993). Furthermore, hairy roots and plants transformed with the ENOD 11-Gus gene show a similar gene expression pattern when mycorrhizal and hairy roots developed from certain Medicago sativa L. (alfalfa) clones are, like the whole plants, resistant to G. margarita but receptive to G. intraradices (Douds et al., 1998). From these observations, one can postulate that the establishment of a mycorrhiza and the biotrophic mode of growth of AM fungi on root-organ cultures may be fundamentally similar to that occurring in vivo.
A particularly important field of study concerns the genetical and physiological basis behind the obligate biotrophic nature of AM fungi. In other words, what allows the fungus to complete its life cycle in the presence of a host root ? The in vitro system, which allows control of most parameters and provides root and fungal material at various interactive stages, should permit more in-depth cellular, biochemical and molecular investigations into this aspect. Since hairy roots from nonhost plants also exhibit a Myc- phenotype, this in vitro system could also be used to address the question as to why some plants are inherently non-mycotrophic.
To elucidate which fungal genes are specifically expressed in planta and which plant symbiotic genes are expressed in mycorrhizal roots requires sophisticated molecular analyses using PCR-based subtractive hybridization methods. For example, fungal genes specifically expressed in planta must be selected against the root genes and the extraradical fungal genes. This can only be achieved with a monoxenic system, such as that outlined earlier, which provides a means of harvesting sufficient quantities of mycorrhizal roots and isolated extraradical mycelium.
In addition to the transfer of T-DNA from the wild plasmid of A. rhizogenes to produce hairy roots, the bacterium can also be used as a vector to transfer other genes of interest. Studies on the role and the expression of putative symbiotic genes, using reporter genes, anti-sense and the overexpression strategy, can, therefore, be carried out using transformed hairy roots. These can be obtained in 2-3 weeks. By contrast, the transformation and regeneration of intact plants usually takes 6 months.
The use of the AM root-organ culture technique has important implications for the production of AM inocula for research and commercial purposes. Although the results from most industry-based research are not generally publicly available, recently Moutoglis and Beland (2001) provided, a brief insight into some of the potential techniques and Jolicoeur et al. (1999) and Jolicoeur and Perrier (2001) proposed a bioreactor-based production technique using root-organ cultures. Although the nutritional parameters determining the productivity within these in vitro systems have been studied, further research is needed to optimize productivity and to develop low-cost techniques for the large-scale production of aseptic inocula.
The potential of the in vitro system for the study of interspecific AM fungal competition has not yet been investigated. Compatibility studies between AM fungi using root-organ cultures are possible and the results might suggest improvements that could be made to industrially produced inocula.
At the intraspecific level, in vitro studies (mycelial development and spore production) have highlighted the existence of phenotypic variations between different G. intradices isolates. The in vitro system could, therefore, be used to study mating types and the heredity nature of certain phenotypes within this species, which may also help improve commercial Inocula ( Fig. 1).
The mycorrhizal root-organ culture has proven useful for taxonomists and physiologists and potentially useful for geneticists. It is also promising for the study of interaction with root-born pathogens and other soil organisms. In its present state, however, the AM root-organ culture system is somewhat limiting because the root is bathed in a carbohydrate-rich solution. Studies of direct interactions between AM colonized roots and pathogens and other soil organisms, could perhaps be achieved using an improved version of the root-hypocotyl system (Bunting and Horrocks, 1964; Miller-Wideman and Watrud, 1984), possibly adding auxins (Fortin and Piche, 1979). Briefly, using system, it would be possible to achieve a polarity whereby organic nutrients, which are absorbed by the aerial part of the plant, are translocated to the roots via the vascular system. Because the root system is exposed only to mineral nutrients, this in vitro system would allow interactions between soil organisms involved in polysaccharide decomposition, nitrogen fixation, nitrate reductase, ammonification and phosphate solubilization to be investigated.
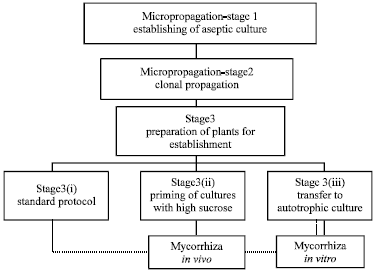 | |
| Fig. 1: | Flow diagram showing alternative strategies for mycorrhization of micro-plants in in vitro and in vivo |
An in vitro tripartite culture system, consisting of the co-culture of in vitro cultivated strawberry plantlets, AM fungi (G. intraradices or Gi. rosea) and a carrot root-organ culture, has proved useful in highlighting the capacity of AM fungi to reduce water stress in micropropagated plantlets (Elmeskaoui et al., 1995). Using this tripartite system, Herninlndez-Sebastia et al. (1999) showed that colonization of in vitro cultivated strawberry plantlets by G. intraradices increased relative plant water content. This effect was related to enhanced water content of the mycorrhizal roots. However, root osmotic potential and dry weight did not significantly differ from that of non-mycorrhizal controls. In an attempt to explain this phenomenon, Herninlndez-Sebastia et al. (2000) proposed that mycorrhizal strawberry plantlets were able to change root cell amino-acid composition and alter cell starch concentrations, to reduce water stress.
To limit the use of AM toxic compounds in the environment, Wan et al. (1998) and Wan and Rahe (1998) used in vitro AM root-organ cultures to study the sublethal toxicity of a range of pesticides (e.g., benomyl, glyphosate, dimethoate and azadirachtin) on Glomales sp. This method could become a standard test for the regulation of pesticides.
The concept behind the use of excised roots for endomycorrhizal research stems from work on ectomycorrhiza. In return, the technique of in vitro culture on transformed roots developed for endomycorrhizal studies has recently been applied to ectomycorrhizal research. A model based on Ri T-DNA transformed roots of the Mediterranean shrub, Cistus incanus, has been developed for the inoculation of plants with the truffle-producing fungus, Tuber melanosporum (Wenkart et al., 2001). In the past, ectomycorrhizal studies have typically been carried out on fungi grown either saprophytically or with plant hosts but under non-sterile conditions (Read, 1992). The transformed Cistus root model will allow the in vitro study of the effects of different edaphic factors on the growth and development of the extraradical phase of ectomycorrhizal fungi.
Initial experiments on fungal growth and development have shown that transformed Cistus roots can be used to form ectomycorrhizae with many species of ectomycorrhizal fungi and to dramatically increase fungal growth (Coughlan et al., 2001). As observed in studies on the endomycorrhizal symbiosis, the presence of transformed roots stimulates the production of highly branched structures absent on hyphae growing saprophytically (unpublished data). The production of these structures is likely to be important for nutrient uptake and formation of the symbiosis. Work is presently underway to characterize the exudates from these roots and to test their activity on endomycorrhizal fungi in an attempt to determine whether a common signaling pathway exists.
In this review, we have attempted to cover all aspects of the mycorrhizal root-organ culture system and to outline the fact that the only real limit to its use is that of the user’s imagination.
REFERENCES
- Azcon-Aguilar, C., M.C. Jaizme-Vega and C. Calvet, 2002. The Contribution of Arbuscular Mycorrhizal Fungi to the Control of Soil Borne Plant Pathogens. In: Mycorrhizal Technology: From Genes To Bioproducts-Achievement and Hurdles in Arbuscular Mycorrhizal Research, Gianinazzi, S. and H. Schuepp (Eds.). Brikhauser, Basel, Switzerland, ISBN-10: 3764364858, pp: 187-198.
Direct Link - Bago, B., S.P. Bentivenga, V. Brenac, I.C. Dodd, Y. Piche and L. Simon, 1998. Molecular analysis of Gigaspora (Glomales, Gigasporaceae). New Phytol., 139: 581-588.
Direct Link - Bago, B., W. Zipfel, R.M. Williams, H. Chamberland, I.G. Lafontaine, W.W. Web and Y. Piche, 1998. In vivo studies on the nuclear behavior of the arbuscular mycorrhizal fungus Gigaspora rosea grown under axenic conditions. Protoplasma, 203: 1-15.
CrossRef - Bago, B., W. Zipfel, R.M. Williams, W.W. Webb and Y. Piche, 1999. Nuclei of symbiotic arbuscular mycorrhizal fungi, as revealed by in vitro 2-photon microscopy. Protoplasma, 209: 77-89.
CrossRef - Becard, G. and Y. Piche, 1990. Physiological factors determining vesicular-arbuscular mycorrhizal formation in host and non-host Ri T-DNA transformed roots. Can. J. Bot., 68: 1260-1264.
Direct Link - Bonfante, P. and S. Perotto, 1995. Strategies of arbuscular mycorrhizal fungi when infecting host plants. New Phytol., 130: 3-21.
CrossRefDirect Link - Budi, S.W., B. Blal and S. Gianinazzi, 1999. Surface-sterilization of Glomus mosseae sporocarps for studying endomycorrhization in vitro. Mycorrhiza, 9: 65-68.
CrossRef - Chabot, S., G. Becard and Y. Piche, 1992. Life cycle of Glomus intraradix in root organ culture. Mycologia, 84: 315-321.
Direct Link - Cliquet, J.B. and G.R. Stewart, 1993. Ammonia assimilation in Zea mays L. infected with a vesicular arbuscular mycorrhizal fungus Glomus fasciculatum. Plant Physiol., 101: 865-871.
Direct Link - Coughlan, P.A., S. Dube, J. Lavaud, F. Marzitelli and Y. Piche, 2001. Developpement in vitro des genres Tubet; laccaria, cenococcum. In: Les symbioses mycorhiziennes: de la comprehension a I' application en biotechnologie environmentale. Institut De Recherche En Biologic Vegetale and the Jardin Botanique de Montreal. Montreal, Quebec, pp: 26.
- Diop, T.A., G. Becard and Y. Piche, 1992. Long-term in vitro culture of an endomycorrhizal fungus, Gigaspora margarita, on Ri T-DNA transformed root of carrot. Symbiosis, 12: 249-259.
Direct Link - Diop, T.A., C. Plenchette and D.G. Strullu, 1994. Dual axenic culture of sheared-root inocula of vesicular-arbuscular mycorrhizal fungi associated with tomato roots. Mycorrhiza, 5: 17-22.
CrossRef - Diop, T.A., C. Plenchette and D.G. Strullu, 1994. . In vitro culture of sheared mycorrhizal roots. Symbiosis, 17: 217-227.
Direct Link - Douds, D.D. Jr., L. Galvez, G. Becard and Y. Kapulnik, 1998. Regulation of mycorrhizal development by plant host and AM fungus species in alfalfa. New Phytol., 138: 27-35.
Direct Link - Elmeskaoui, A., J.P. Damon, M.J. Poulin, Y. Piche and Y. Desjardins, 1995. A tripartite culture system for endomycorrhizal inoculation of micropropagated strawberry plantlets. Mycorrhiza, 5: 313-319.
CrossRef - Fisher, R.A. and N.C. Turner, 1978. Plant productivity in the arid and semi-arid zones. Ann. Rev. Plant Physiol., 29: 277-317.
Direct Link - Forbes, P.J., S. Millam, J.E. Hooker and L.A. Harrier, 1998. Transformation of the arbuscular mycorrhiza Gigaspora rosea by particle bombardment. Mycol. Res., 102: 497-501.
CrossRef - Fortin, J.A. and Y. Piche, 1979. Cultivation of Pinus strobus root-hypocotyl explants for synthesis of ectomycorrhiza. New Phytol., 83: 109-119.
Direct Link - Furlan, V., H. Biirtschi and J.A. Fortin, 1980. Media for density gradient extraction of endomycorrhizal spores. Trans. Br. Mycol. Soc., 75: 336-338.
Direct Link - Giovannetti, M. and B. Mosse, 1980. An evaluation of techniques to measure vesicular-arbuscular infection in roots. New Phytol., 84: 489-500.
CrossRefDirect Link - Gryndler, M., H. Hrselova, I. Chvatalova and M. Vosatka, 1998. In vitro proliferation of Glomus fistulosum intraradical hyphae from mycorrhizal root segments in maize. Mycol. Res., 102: 1067-1073.
Direct Link - Hernilndez-Sebastiii, C., Y. Piche and Y. Desjardins, 1999. Increase of relative water content in whole strawberry in vitro inoculated with Glomus intraradices in a tripartite system. Plant Sci., 143: 81-91.
Direct Link - Hernilndez-Sebastia, C., G. Samson, P.Y. Bernier, Y. Piche and Y. Desjardins, 2000. Glomus intra radicesM causes differential changes in amino acid and starch concentrations of in vitro strawberry subjected to water stress. New Phytol., 148: 177-186.
Direct Link - Jackson, M.L., 1973. Soil Chemical Analysis. 1st Edn., Prentice Hall Ltd., New Delhi, India, Pages: 498.
Direct Link - Jolicoeur, M., R.D. Williams, C. Chavarie, J.A. Fortin and J. Archambault, 1999. Production of Glomus intra radices propagules, an arbuscular mycorrhizal fungus, in an airlift bioreactor. Biotechnol. Bioeng, 63: 224-232.
Direct Link - Karandashov, V.E., I.N. Kuzourina, E. George and H. Marschner, 1999. Monoxenic culture of arbuscular fungi and plant hairy roots. Russ. J. Plant Physiol., 46: 87-92.
Direct Link - Koide, R.T. and R.P. Schriener, 1992. Regulation of the vesicular arbuscular mycorrhizal symbiosis. Ann. Rev. Plant Physiol. Mol. Biol., 43: 557-581.
Direct Link - Mathur, N. and A. Vyas, 1995. In vitro production of Glomus deserticola in association with Ziziphus nummularia. Plant Cell Rep., 14: 735-737.
CrossRefDirect Link - Mathur, N. and A. Vyas, 1996. I: Influence of VA mycorrhizae on net photosynthesis and transpiration of Ziziphus mauritiana. J. Plant Physiol., 147: 328-330.
Direct Link - Mathur, N. and A. Vyas, 1995. II Changes in enzymes of nitrogen metabolism by VA Mycorrhizae in Ziziphus nummularia. J. Plant Physiol., 147: 331-333.
Direct Link - Mathur, N. and A. Vyas, 1996. Biochemical changes in Ziziphus xylopyrus by VA Mycorrhizae. Bot. Bull. Acad. Sin., 37: 209-212.
Direct Link - Mathur, N. and A. Vyas, 1999. Improved biomass production, nutrient uptake and establishment of in vitro raised Ziziphus mauritiana by VA mycorrhiza. J. Plant. Physiol., 155: 129-132.
Direct Link - Mathur, N. and A. Vyas, 2000. Influence of arbuscular mycorrhizae on biomass production, nutrient uptake and physiological changes in Ziziphus mauritiana Lam. under water stress. J. Arid Environ., 45: 191-195.
CrossRefDirect Link - Mosse, B. and C. Hepper, 1975. Vesicular-arbuscular mycorrhizal [Endogone] infections in root organ cultures [Red clover, tomatoes, fungus diseases]. Physicl. Plant. Pathol., 5: 215-223.
Direct Link - Schreiner, R.P. and R.T. Koide, 1993. Stimulation of vesicular-arbuscular fungi by mycotrophic and non mycotrophic plant root systems. Applied Environ. Microbiol., 59: 2750-2752.
Direct Link - Smith, S.E. and D.J. Read, 1997. Mycorrhizal Symbiosis. 2nd Edn., Academic Press, London, ISBN: 0126528403.
Direct Link - Wan, M.T. and J.E. Rahe, 1998. Impact of azadirachtin on Glomus intraradices and vesicular-arbuscular mycorrhiza in roots inducing transferred DNA transformed roots of Daucus carota. Environ. Toxicol. J. Chem., 17: 2041-2050.
Direct Link - Wan, M.T., J.E. Rahe and R.G. Watts, 1998. A new technique for determining the sublethal toxicity of pesticides to the vesicular-arbuscular fungus Glomus intraradices. Environ. Toxicol. Chem., 17: 1421-1428.
Direct Link - Wenkart, S., N. Roth-Bejerano, D. Mills and V. Kagan-Zur, 2001. Mycorrhizal associations between Tuber melanosporum mycelia and transformed roots of Cistus incanus. Plant. Cell. Rep., 20: 369-373.
Direct Link - White, P.R., 1943. A Handbook of Plant Tissue Culture. Jacques Cattell Press, Lancaster, PA., USA.
Direct Link - Strullu, D.G. and C. Plenchette, 1991. The entrapment of Glomus sp. in alginate beads and their use as root inoculum. Mycol. Res., 95: 1194-1196.
Direct Link - Gerdemann, J.W. and T.H. Nicolson, 1963. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans. Br. Mycol. Soc., 46: 235-244.
CrossRefDirect Link
Abdoulaye BEYE Reply
Good article. Thanks for opening access!