
A. Gomaa
Department of Food and Animal Sciences, Alabama A and M University, United States of America
S. Willis
Department of Food and Animal Sciences, Alabama A and M University, United States of America
M. Verghese
Department of Food and Animal Sciences, Alabama A and M University, United States of America
J. Boateng
Department of Food and Animal Sciences, Alabama A and M University, United States of America
LiveDNA: 1.12516
American Journal of Food Technology
Year: 2021 | Volume: 16 | Issue: 1 | Page No.: 18-30







ABSTRACT
Background and Objective: Carob pods are natural sources of fermentable fibers and polyphenols, which have been shown to have health benefits. Konjac is a plant known for its dietary fiber, health benefits and food applications. The utilization of carob and konjac as prebiotic sources could serve as novel substrates for improving probiotics viability and stability. This study examined the effect of carob and konjac (2.5 and 5%) on the growth of four probiotic strains (Bifidobacterium bifidum, Lactobacillus paracasei, Lactobacillus acidophilus and Streptococcus thermophilus) and the effects of fermentation on total phytochemical contents and antioxidant activity. Materials and Methods: Total viable counts and pH were measured for assessing prebiotic activity. Total phenolics, total flavonoids, DPPH, FRAP, TEAC and ORAC were measured in fermented and unfermented samples. The analysis was conducted in triplicates. Results: The highest growth fold (Log CFU) was noted with a combination of L. acidophilus and 5% carob after 48 hrs incubation (2.02 Log CFU) and L. paracasei and 2.5% konjac after 24 hrs incubation (2.13 Log CFU). Total phenolic contents and total flavonoids were highest after 48 hrs fermentation of 2.5% carob with B. bifidum (518.07 mg GA 100 g–1 and 344.37 mg CE 100 g–1). The DPPH, TEAC and FRAP activity ranged from (12.15-92.85%, 4.13-24.8 mM Trolox g–1 and 11.25-31.4 mM FeSO4 g–1, respectively) in all samples. For ORAC, the majority of fermented extracts showed better values (0.04-5.46 uM Trolox g–1). Conclusion: Accordingly, carob pods and konjac extracts may be considered as effective prebiotic for increasing probiotics growth and viability and particularly carob as a source of antioxidants.
PDF Abstract XML References Citation
How to cite this article
A. Gomaa, S. Willis, M. Verghese and J. Boateng, 2021. Probiotic Fermentation of Konjac and Carob Pods Ceratonia siliqua and Observation of Related Antioxidant Activity. American Journal of Food Technology, 16: 18-30.
DOI: 10.3923/ajft.2021.18.30
URL: https://scialert.net/abstract/?doi=ajft.2021.18.30
DOI: 10.3923/ajft.2021.18.30
URL: https://scialert.net/abstract/?doi=ajft.2021.18.30
INTRODUCTION
Probiotic fermented foods and beverages are highly trending in the food industry1. According to the World Health Organization (WHO)2, probiotics are defined as “live strains of strictly selected microorganisms which, when administered in adequate amounts confer a health benefit on the host”. Probiotics have been shown to improve human health and alleviate many chronic diseases such as insulin resistance3,4, obesity5,6, colon cancer7,8, lactose intolerance9 and oxidative stress10,11.
Prebiotics are a type of non-digestible food that beneficially affects human health. The difference between prebiotics and dietary fibers is that prebiotics has to selectively stimulate the growth and/or activity of good microorganisms in the colon12 and is poorly utilized by potentially harmful bacteria. Dietary fibers such as insulin and fructooligosaccharides act beneficially on the human body as a prebiotic substrate for probiotics to enhance its growth and encourage the production of short-chain fatty acids (SCFAs) via fermentation13.
Konjac (Amorphophallus konjac) (Fig. 1) is a perennial plant, growing widely in East Asia. Konjac has been suggested to have many health benefits including, anti-diabetic, anti-inflammatory and prebiotic activity14. Moreover, konjac is utilized in the food industry (Food additive: E 425i and ii) as a film former, stabilizer, thickener and gelling agent. Glucomannan is a water-soluble polysaccharide that accounts for approximately 40% of the of plant’s bulbo-tuber14. Glucomannan is made primarily of mannose and D-glucose as a secondary sugar with β-(1-4) glycosidic bond15. As a highly fermentable fiber, glucomannan are highly fermentable by Bifidobacterium16, favor the growth of probiotics over the growth of pathogens such as Staphylococcus aureus and Salmonella typhimurium17 and attenuate Inflammatory Bowel Disease (IBD) related symptoms18. The ability of konjac to form thermo-irreversible gel and its shear thinning properties makes it a prime candidate for prebiotic selection as it can exhibit stability in different food processing conditions.
Carob (Ceratonia siliqua) (Fig. 2) is the fruit of a Mediterranean evergreen tree belonging to the legume family, Fabaceae19. Carob has been utilized throughout history as a binding agent and medicinal beverage20. Carob is a good source for polyphenols19,21, which have shown to selectively enhance the growth of lactic acid bacteria22 in addition to their stimulation of SCFA production23. Furthermore, carob is rich ingalactomannans24 a type of polysaccharides, which are not digestible in the gastrointestinal tract25 and thus are able to stimulate the growth of lactobacilli and bifidobacteria15,26.
 |
| Fig. 1: | Konjac roots Adapted from https://www.precisionnutrition.com/all-about- glucomannan |
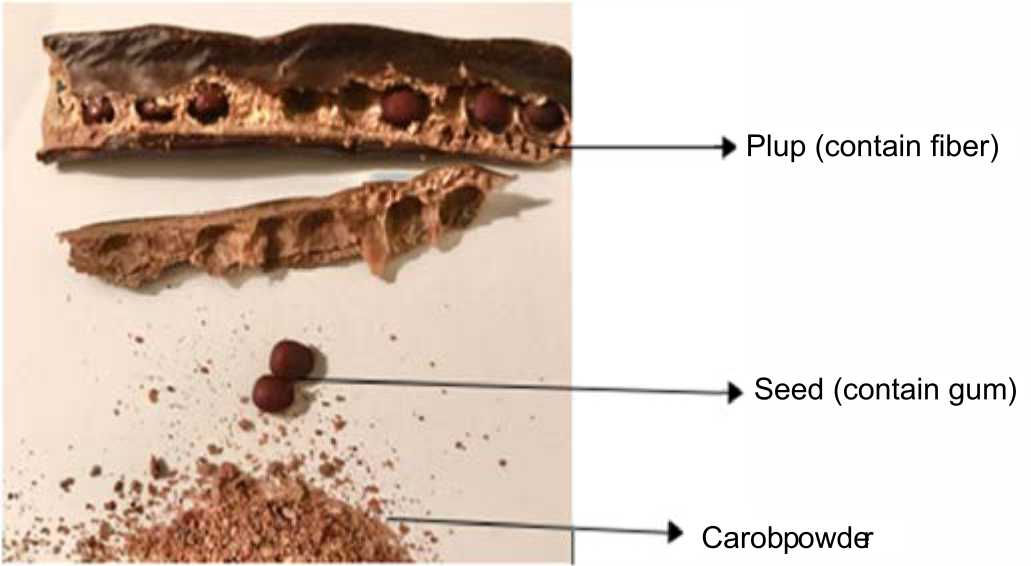 |
| Fig. 2: | Mature carob pod and its constituents |
The predominant polyphenols in carob fruit are phenolic acids, flavonols and condensed tannins (proanthocyanidins)27. Carob pods are a great source for many types of anti-inflammatory and antioxidants as both seeds and pulp contain high concentrations of different polyphenols classes. Carob pulp contains phenolic acids such as 4-hydroxybenzoic acid, gallic, caffeic and ferulic acids, flavonoids such as quercetin rhamnoside, eriodictyol, genistein28 and tannins such as tannic acid. Further, the flour and pods are rich in myricetin, methyl gallate, catechin and ellagitannins29,30.
The aim of this study was to compare the potential of carob pods and konjac root powder as prebiotic substrates for the growth of four probiotic strains: Bifidobacterium bifidum (Danisco Bb-06®), Lactobacillus paracasei (Danisco Lpc-37®), Lactobacillus acidophilus (Danisco La-14®) and Streptococcus thermophilus (Danisco St-21®) and also determine the total phytochemical content (total phenolics content and total flavonoids content) and antioxidant activity (DPPH, FRAP, TEAC and ORAC) of fermented and unfermented substrates.
MATERIALS AND METHODS
Study area: The study was carried out in the Food Science Department at Alabama A and M University, Normal AL from April, 2019 to December, 2019.
Materials: The M17 and MRS media were obtained from BD DifcoTM and BD BBLTM. Carob pods were purchased from a local store (Brooklyn, NY). Konjac Root powder (90% Glucomannan) was purchased from Hard Eight Nutrition LLC, (Henderson, NV). Oxoid Anaero Gen 2.5 L Sachets, anaerobic atmosphere generation system (Gas Pak, Thermo Scientific, Hampshire, UK). Ethanol, sodium carbonate, sodium hydroxide, aluminum chloride, sodium acetate, acetic acid, 2,4,6-tri(2-pyridyl)-s-triazine (TPTZ), hydrochloric acid, ferric chloride, ferrous sulfate, 2,2’-azinobis (3-ethylbenzothiazoline 6-sulfonate) radical cation, Trolox and potassium persulfate (Fisher Scientific, Waltham, MA). Gallic acid, catechin,1-diphenyl-2-picrylhydrazyl (DPPH) radical and 2,2 azo bis 2-amidinopropane dihydrochloride (AAPH) (Sigma-Aldrich). Folin-Ciocalteu reagent (MP Biomedical).
Probiotic propagation and fermentation of prebiotic substrates: Probiotic cultures (L. acidophilus (La-14), L. paracasei (Lpc-37), S. thermophilus (St-21) and B. bifidum (Bb-06)) were kindly provided by DuPontTM Danisco® Food Ingredients (Copenhagen, Denmark) and were stored and rehydrated according to ATCC® reviving freeze-dried microorganisms methods.
Growth media were prepared and stored according to manufacturer's methods. All the probiotics strains were maintained at 38°C in anaerobic conditions except for S. thermophilus, which was maintained at 40°C. Probiotic strains were added to the perspective media/treatment combination and were fermented for 24 or 48 hrs. After fermentation, pH was taken and CFU was determined following standard protocols.
Carob pods were blended into a homogenous powder. Carob and konjac mixtures were prepared by mixing 2.5 and 5 g (2.5 and 5%) with 100 mL of selected broth (MRS/M17). Each concentration served as a medium for probiotic strains and each mixture was incubated for two time periods, 24 and 48 hrs. Media without carob/konjac served as control (24 and 48 hrs incubation). Samples were taken from each treatment and were serially diluted for microbial analysis.
Total phenolic and flavonoid contents: Total Phenolics Content (TPC) and Total Flavonoids Content (TFC) were determined using Folin-Ciocalteu’s and aluminum colorimetric methods, according to Patel et al.31. For TPC, Gallic Acid (GA) was used as a standard. The absorbance was measured at 750 nm after incubating for 90 min at room temperature. For TFC, the absorbance was measured at 510 nm using catechin (CE) as a standard.
Antioxidant assays: DPPH (2,2-Diphenyl-1-Picrylhydrazyl) radical scavenging activity, ferric reducing antioxidative potential (FRAP) and Trolox equivalent antioxidant capacity (TEAC) was determined according to Patel et al.31 and oxygen radical absorbance capacity (ORAC) was determined according to Haile and Kang32.
Preparation of fermented and unfermented carob extracts for total polyphenol and antioxidant analysis
Unfermented extracts: Five grams of carob/konjac powder was mixed in 50 mL of 80% ethanol for 2 hrs with continuous shaking. The extracts were centrifuged at 5000 g for 20 min. Supernatants were filtered and the remaining residue was washed with 80% ethanol re-extracted. Supernatants were pooled and the solvent was evaporated using a rotary evaporator (Buchi Rotavapor R-215). The concentrates were stored at -80°C until further use.
Fermented extracts: Cell-free supernatant (CFS) of the media/carob mixtures were prepared using a method by Xing et al.33 with some modifications to the centrifugation conditions. Probiotics samples were centrifuged (10,000 g, 20 min, 4°C) and the resulting supernatant was filtered twice with (0.22 μm pore size syringe filters) into 10 mL centrifuge tubes. The CFS was stored at -80°C until use.
Statistical analysis: The experiment was conducted in triplicates and SAS 9.4 software (SAS Institute Inc., Cary, NC, USA) was used. The ANOVA was performed and means separation was done by Student’s two-tailed t-test and Tukey-Kramer range test at a 5% significance level. Data are reported as Means±SEM.
RESULTS
Effect of konjac and carob on pH of probiotics: Table 1 shows the effect of carob and konjac on the pH. All the synbiotic combinations were able to reduce the pH of the medium regardless of the treatment concentrations.
| Table 1: Effect of carob pods and konjac on acidification (pH) of probiotics growth media | |||||
| Probiotics | (Control no prebiotic) | pH 2.5% Carob | pH 5% Carob | pH 2.5% Konjac | pH 5% Konjac |
| 24 hrs | |||||
| L. paracasei | 3.9±0.10d,z | 4.75±0.04a,x | 4.27±0.05b,y | 4.0±0.01d,z | 4.02±0.02cd,z |
| B. bifidum | 4.43±0.01bc,x | 4.03±0.08d,y | 4.17±0.07b,y | 4.39±0.02b,x | 4.43±0.01b,x |
| L. acidophilus | 3.72±0.10e,z | 4.43±0.05b,x | 4.13±0.06bc,y | 4.19±0.01c,y | 4.12±0.03bc,y |
| S. thermophilus | 5.5±0.01a,x | 4.78±0.02a,z | 4.72±0.01a,wz | 4.61±0.09a,z | 5.27±0.03a,y |
| 48 hrs | |||||
| L. paracasei | 3.74±0.08de,y | 3.55±0.06f,z | 3.6±0.04d,z | 3.83±0.00e,xy | 3.89±0.00cd,x |
| B. bifidum | 4.57±0.07b,x | 4.41±0.05b,xy | 4.03±0.02c,y | 4.34±0.01b,xy | 4.23±0.3bc,xy |
| L. acidophilus | 3.91±0.05d,x | 3.86±0.05e,y | 3.72±0.05d,z | 3.90±0.01de,xy | 3.73±0.02d,z |
| S. thermophilus | 4.32±0.03c,z | 4.2±0.01c,w | 4.61±0.01a,x | 4.4±0.01b,y | 4.13±0.01bc,u |
| Results are expressed as means of triplicates±SEM, Subscripts (abc) represent differences amongst columns (p< 0.05). Subscripts (xyz) represent differences between rows (p<0.05) | |||||
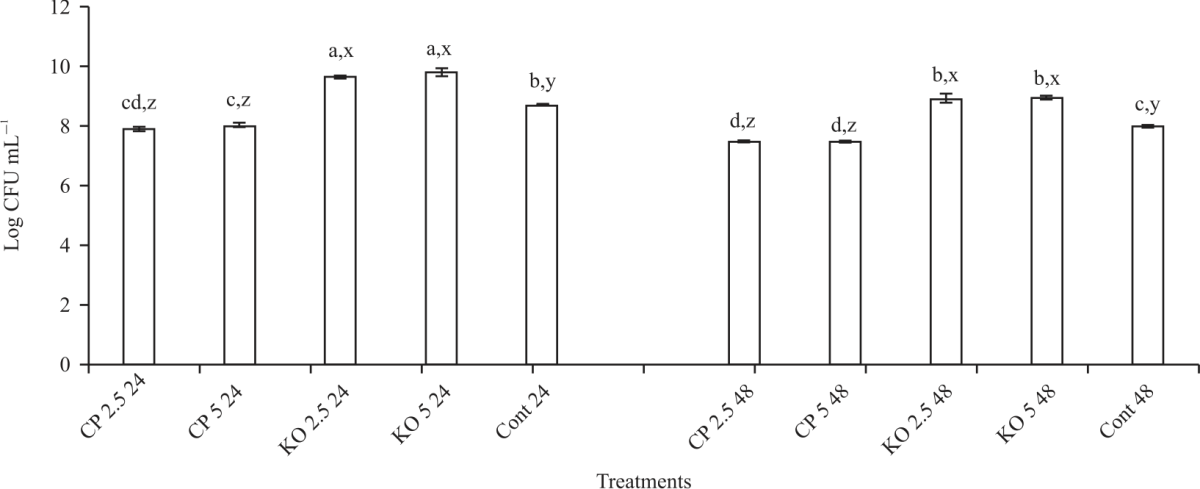 |
| Fig. 3: | Effect of carob and konjac on growth (CFU) of L. acidophilus Values are means (n = 3) ± SEM. Subscripts (abc) represent differences amongst all treatments (p<0.05). Subscripts (xyz) represent differences between treatments within the same incubation time (p<0.05), CP: Carob powder, KO: Konjac, Cont: Control without treatment |
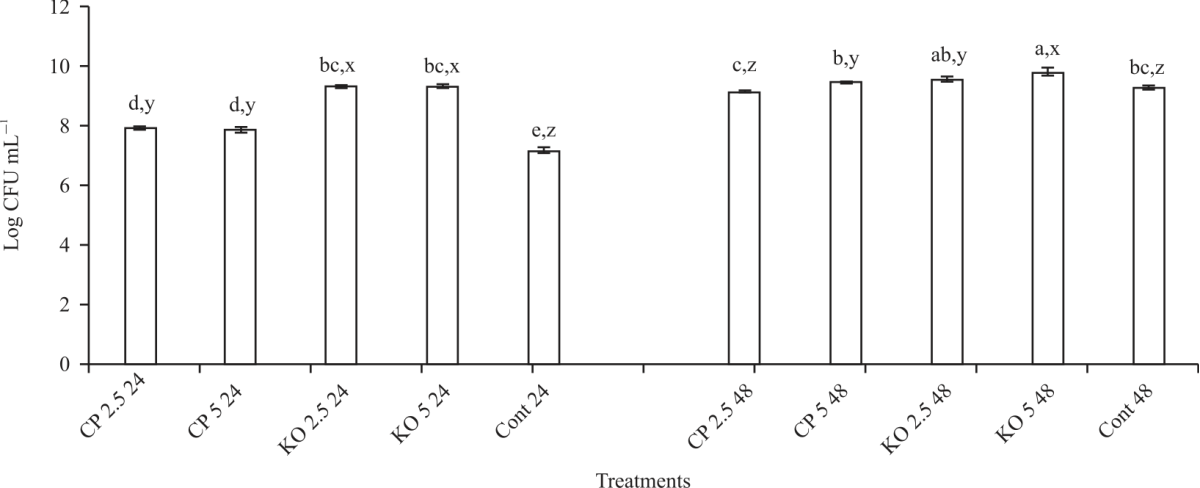 |
| Fig. 4: | Effect of carob and konjac on growth (CFU) of L. paracasei Values are means (n = 3) ±SEM. Subscripts (abc) represent differences amongst all treatments (p<0.05). Subscripts (xyz) represent differences between treatments within the same incubation time (p<0.05). CP: Carob powder, KO: Konjac, Cont: Control without treatment |
After 24 hrs incubation, B. bifidum +2.5 (4.03) and 5% (4.17) carob and S. thermophilus plus all prebiotic combinations showed the most significant (p<0.05) acidification compared to the control (no prebiotic). After 48 hrs incubation, L. paracasei+ 2.5% (3.55) and L. paracasei+ 5% (3.6) showed significant (p<0.05) reduction in pH. For B. bifidum, a significant (p<0.05) drop in pH was noted with+5% carob (4.03).
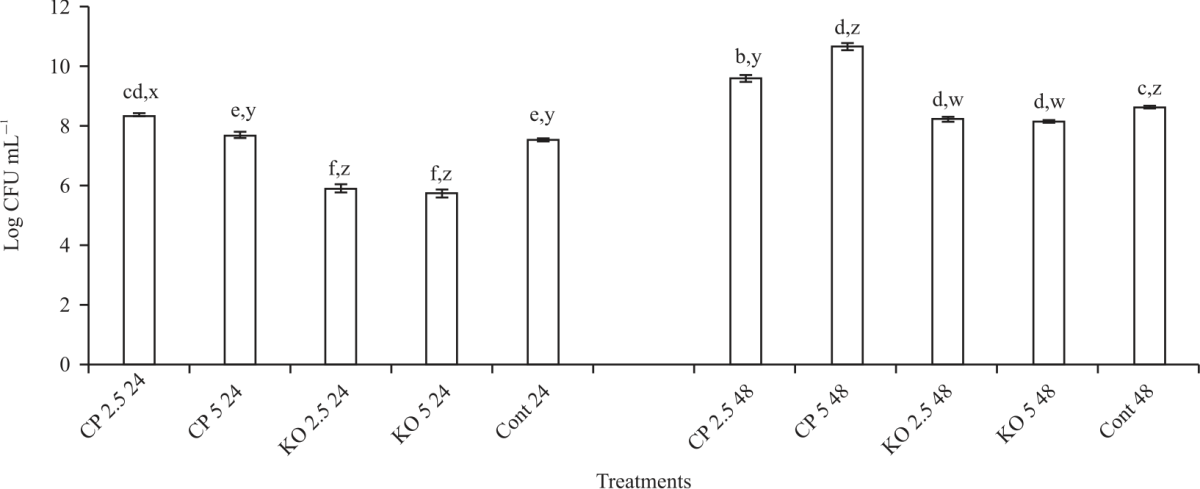 |
| Fig. 5: | Effect of carob and konjac on growth (CFU) of B. bifidum Values are means (n = 3) ±SEM. Subscripts (abc) represent differences amongst all treatments (p<0.05). Subscripts (xyz) represent differences between treatments within the same incubation time (p<0.05), CP: Carob powder, KO: Konjac, Cont: Control without treatment |
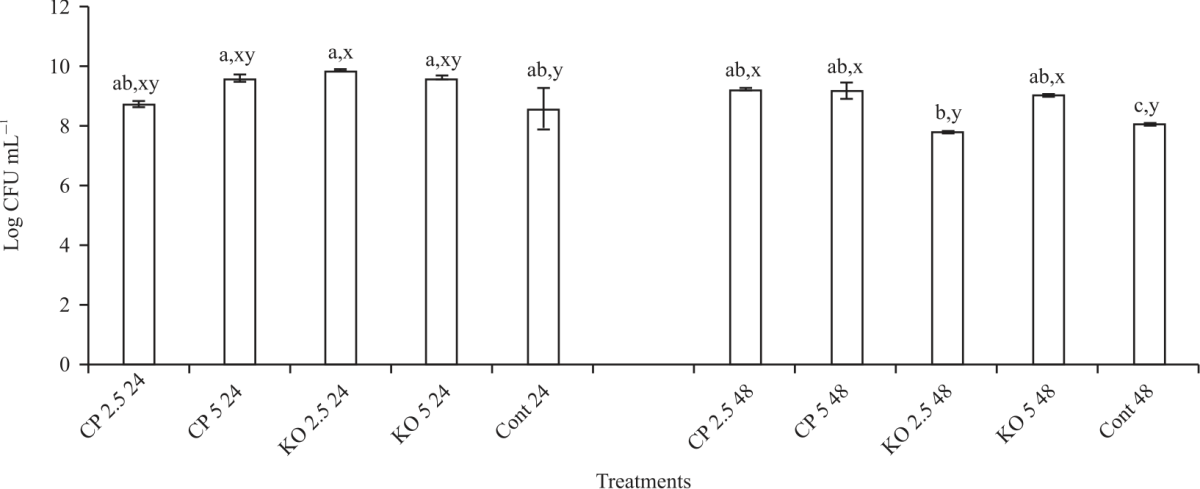 |
| Fig. 6: | Effect of carob and konjac on growth (CFU) of S. thermophilus Values are means (n = 3) ±SEM. Subscripts (abc) represent differences amongst all treatments (p<0.05). Subscripts (xyz) represent differences between treatments within the same incubation time (p<0.05), CP: Carob powder, KO: Konjac, Cont: Control without treatment |
Except for L. acidophilus +2.5 konjac, all the prebiotics types incubated with the probiotic saw significant (p<0.05) decreases in pH when compared to the control (3.91). For S. thermophilus, the combination with 5% konjac indicated significant (p<0.05) decrease in pH (4.13) when compared to the control (4.32). Overall, there were significant (p<0.05) variations in pH with incubation times for the control and the synbiotic treatments. However, the results indicated the lowest pH after 48 hrs incubation for L. paracasei and L. acidophilus with all the prebiotics.
Effect of konjac and carob on growth of probiotics: Figure 3 shows the effects of konjac and carob on the growth of the L. acidophilus. After 24 hrs incubation, the following synbiotic combinations showed the most significant (p<0.05) growth, L. acidophilus+2.5% konjac (9.65 Log CFU) and L. acidophilus+5% konjac (9.8 Log CFU). The least growth was noted for L. acidophilus+2.5% carob (7.8 Log CFU) and L. acidophilus+5% carob (8.1 Log CFU). Similar results were noted after 48 hrs of incubation with the prebiotics, although the Log CFU was decreased by 7-8% for konjac and 5-6% for carob treatments. Figure 4 showed the effects of konjac and carbon the growth of L. paracasei. The results show both konjac and carob significantly (p<0.05) increased the growth of L. paracasei after 24 hrs compared to the control. However, konjac was most effective at improving the growth of the probiotic with 9.2 log CFU for both 2.5 and 5%, compared to 7.8 log CFU for both concentrations of carob and the control (7.1 log CFU).
| Table 2: Polyphenols and flavonoids content in probiotic fermented carob pods extracts | ||
| Treatments | Total phenolic content (mg GA 100 g–1) | Total flavonoid content (mg CE 100 g–1) |
| 24 hrs | ||
| L. paracasei 2.5% | 61.31±10.07e,wz | 52.66±0.006ij,w |
| B. bifidum 2.5% | 516.90±5.66a,x | 381.74±13.7b,y |
| L. acidophilus 2.5% | 70.11±6.47de,z | 137.42±2.87ef,z |
| S. thermophilus 2.5% | 517.07±5.63a,x | 429.43±4.79a,x |
| L. paracasei 5% | 40.62±0.62f,e | 38.88±1.04j,w |
| B. bifidum 5% | 258.20±2.87c,y | 160.29±37.20e,z |
| L. acidophilus 5% | 42.53±1.66f,uw | 47.26±3.78ij,w |
| S. thermophilus 5% | 274.55±12.16b,y | 343.31±0.71c,y |
| 48 hrs | ||
| L. paracasei 2.5% | 76.09±0.87de,z | 112.21±0.67efg,uw |
| B. bifidum 2.5% | 518.07±1.06a,x | 428.11±19.87a,x |
| L. acidophilus 2.5% | 77.9±3.92d,z | 125.49±3.58efg,w |
| S. thermophilus 2.5% | 517.90±7.96a,x | 341.10±10.03c,y |
| L. paracasei 5% | 45.26±2.71f,w | 81.52±1.76hi,u |
| B. bifidum 5% | 259.61±2.46bc,y | 274.43±24.08d,z |
| L. acidophilus 5% | 38.46±2.81gf,w | 92.32±1.23gh,uw |
| S. thermophilus 5% | 262.5±3.69bc,y | 306.68±5.16cd,yz |
| No fermentation | ||
| Un Fermented | 24.04±0.42g | 31.08±0.8j |
| Values are means (n = 3)±SEM. Subscripts (abc) represent differences amongst all treatments (p<0.05), Subscripts (xyz) represent differences between treatments within the same incubation time (p<0.05). Abbreviations: GA: Gallic Acid and CE: Catechin | ||
Although, the growth of L. paracasei+konjac (both concentrations) was significantly increased after 24 hrs when compared to carob and the control, this trend was not observed after 48 hrs, in fact, the results showed there was no difference (p<0.05) between konjac and carob treatment and the control. In Fig. 5, B. bifidum+carob showed the highest (p<0.05) growth after 24 hrs (8.3 and 7.6 log CFU for 2.5 and 5%, respectively) and after 48 hrs (9.5 and 10.6 log CFU for 2.5 and 5%, respectively) incubation. On the other hand, konjac decreased (p<0.05) the growth of B. bifidum (5.8 and 8.1 log CFU for both concentrations at 24 and 48 hrs, respectively). Figure 6 shows the effect of carob and konjac on the growth of S. thermophilus. The results showed no significant difference in growth after 24 hrs incubation with both prebiotics. Even though the growth of the probiotic remained unchanged for most treatment combinations after 48 hrs incubation, observed decreased (p<0.05) growth of S. thermophilus+ 2.5% konjac (7.8 log CFU). This was 21% drop in growth after 24 hrs incubation. Overall, incubation with 5% konjac for 24 hrs showed the best growth with L. acidophilus, 2.5% konjac for 24 hrs showed the best growth with L. paracasei, 5% carob for 48 hrs showed the best growth with B. bifidum and 2.5% konjac for 24 hrs showed the best growth with S. thermophilus.
Phytochemical content of fermented and unfermented carob: In preliminary experiments barely detected TPC, TFC and antioxidant activities in konjac root extract. The TPC and TFC in carob pods were significantly affected by the fermentation (Table 2). After 24 hrs fermentation, TPC increased significantly (p<0.05) with all probiotic strains especially, B. bifidum +2.5% carob (516.9 GA mg 100 g–1) and S. thermophilus+2.5% carob (517.07 GA mg 100 g–1) compared to unfermented carob. Among the treatment groups, the lowest (p<0.05) TPC was found in L. paracasei+5% carob (40.62 GA mg 100 g–1) and in L. acidophilus+5% carob. Interestingly, it was observed that TPC was significantly decreased in synbiotics containing high concentrations of carob. Similar observations were made after 48 hrs fermentation. Another interesting note was that TPC was increased or remained unchanged in most synbiotics after 48 hrs fermentation. TFC in all the synbiotics were significantly (p<0.05) increased compared to the control (unfermented sample) after 24 hrs fermentation, except L. paracasei+2.5%. Among the treatments, TFC in S. thermophilus+2.5 and 5% (429.43 and 343.31 CE mg 100 g–1, respectively) were the highest (p<0.05), followed by B. bifidum+at both concentrations. Like TPC, it was noted a decrease in TFC with higher concentrations of carob. A similar trend was observed with 48 hrs fermentations. All the combinations showed significantly (p<0.05) higher values after 48 hrs fermentation compared to 24 hrs fermentation, except for L. acidophilus and S. thermophilus.
Antioxidant activity of fermented and unfermented carob: The DPPH radical scavenging activity of unfermented carob was 92.85%, whereas fermented carob samples varied from 12.5-88.34% (Fig. 7). The highest DPPH (%) after fermentation was observed with the following combinations, L. paracasei +2.5 (88.34%), L. paracasei+5% carob (85.24%) and L. acidophilus+5% carob (85.43%) after 24 hrs incubation, while the lowest DPPH (%) was observed in L. bifidum at both concentrations (23.95 and 30.27 for 2.5 and 5%, respectively).
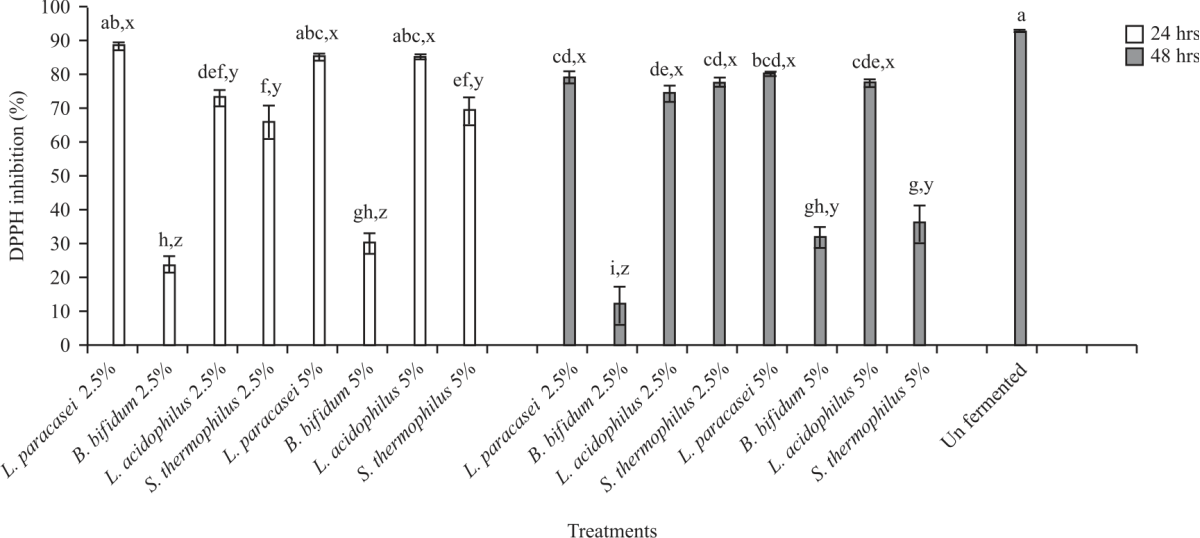 |
| Fig. 7: | Effect of fermented and unfermented carob on DPPH (2,2-diphenyl-1-picrylhydrazyl radical) inhibition (%) Values are means (n = 3) ±SEM. Subscripts (abc) represent differences amongst all treatments (p<0.05). Subscripts (xyz) represent differences between treatments within the same incubation time (p<0.05) |
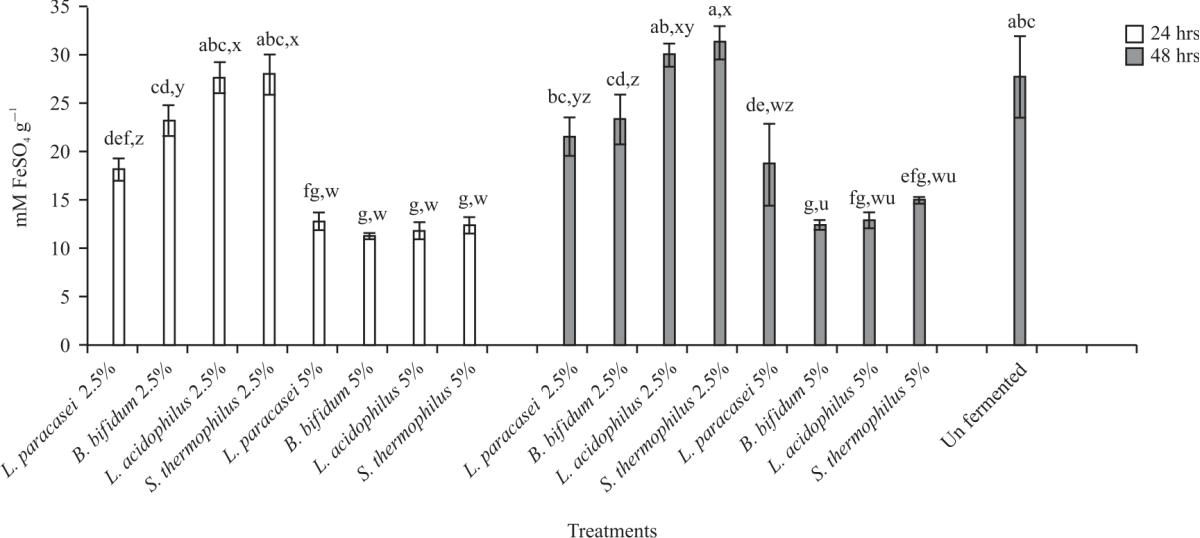 |
| Fig. 8: | FRAP activity of fermented and unfermented carob Values are means (n = 3) ±SEM. Subscripts (abc) represent differences amongst all treatments (p<0.05). Subscripts (xyz) represent differences between treatments within the same incubation time (p<0.05), FRAP: Ferric reducing antioxidant power |
The DPPH (%) decreased with the increase in incubation time in most of the synbiotics treatments. After 24 hrs fermentation, FRAP activity (Fig. 8) in unfermented carob (27.87 mM FeSO4 g–1) was significantly (p<0.05) higher than most of the 24 hrs fermented combinations except B. bifidum+2.5%, L. acidophilus+2.5% and S. thermophilus +2.5%. The results further indicated all the 2.5% combinations were significantly (p<0.05) higher than the 5% combinations except for L. paracasei. After 48 hrs, fermentation did not impact (p<0.05) FRAP activity in all the 2.5% combinations when compared to the unfermented sample, however, 5% combinations saw significant (p<0.05) reductions in FRAP activities. The highest FRAP values after 48 hrs fermentation were noted in S. thermophilus+2.5% (31.4 mM FeSO4 g–1), followed by L. acidophilus +2.5% (30.12). Overall, it was noted, no significant differences in FRAP activities after 24 and 48 hrs fermentations. Trolox equivalence (TEAC) (Fig. 9) of unfermented carob (19.91 Mm Trolox g–1) was significantly (p<0.05) higher compared to all the fermented samples except for S. thermophilus + 2.5% after 24 hrs fermentation (19.2).
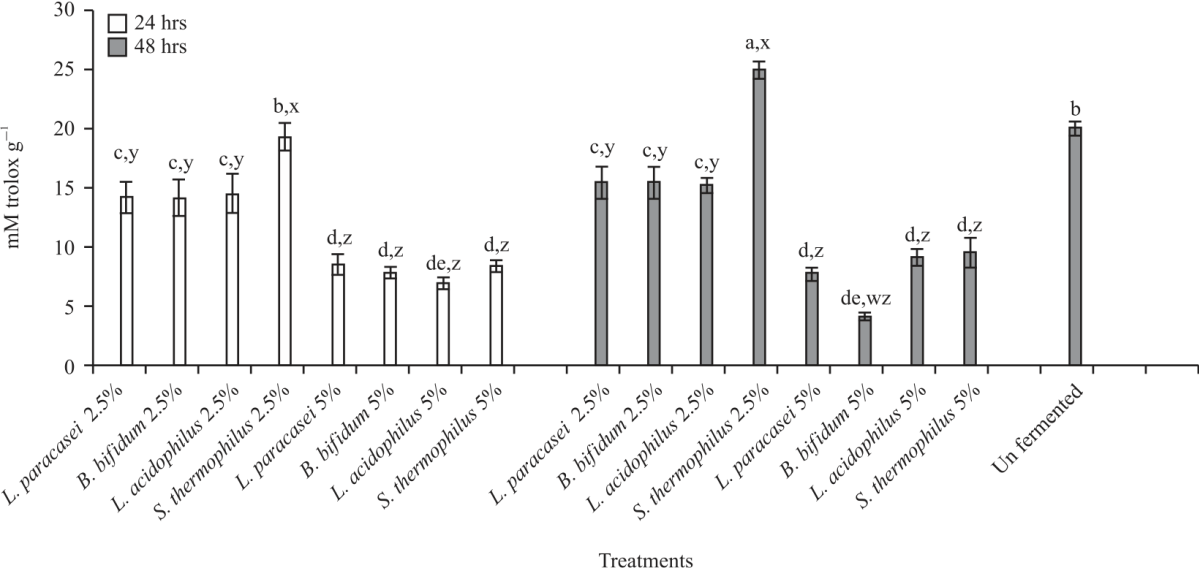 |
| Fig. 9: | TEAC activity of fermented and unfermented carob Values are means (n = 3) ± SEM. ND-not detected. Subscripts (abc) represent differences amongst all treatments (p<0.05). Subscripts (xyz) represent differences between treatments within the same incubation time (p<0.05), TEAC: Trolox equivalent antioxidant capacity |
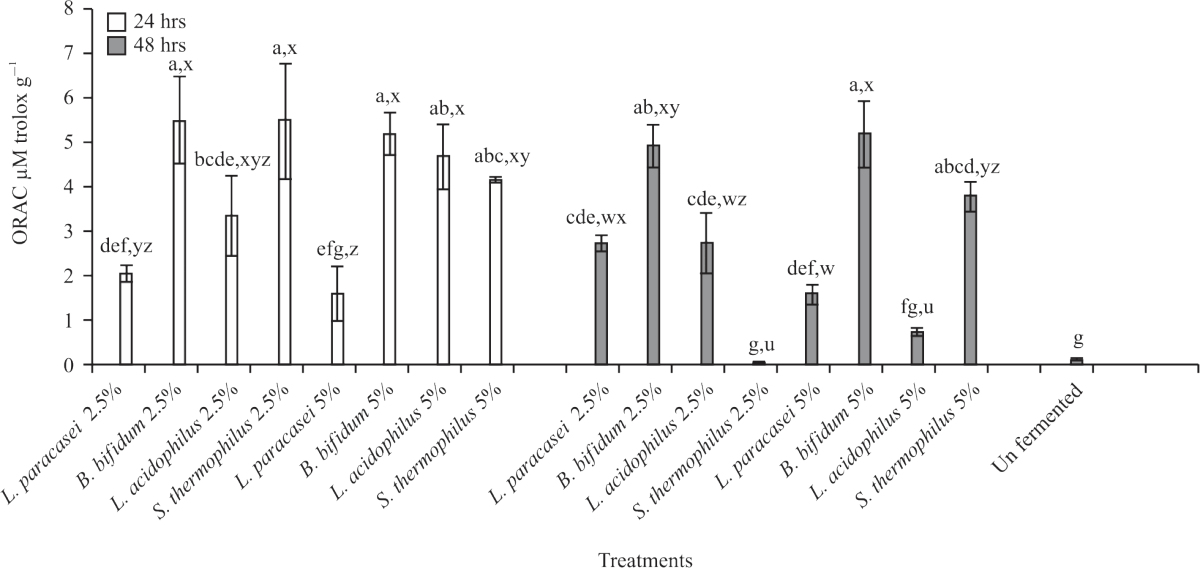 |
| Fig. 10: | ORAC activity of fermented and unfermented carob Values are means (n = 3) ±SEM. ND-not detected. Subscripts (abc) represent differences amongst all treatments (p<0.05). Subscripts (xyz) represent differences between treatments within the same incubation time (p<0.05), ORAC: Oxygen radical absorbance capacity |
Similar observations were made after 48 hrs fermentation, except for S. thermophilus+ 2.5% (24.8). Among the treatments, combinations with 2.5% carob showed significant (p<0.05) increases in TEAC compared to 5% combinations after 24 and 48 hrs fermentation. The ORAC values (Fig. 10) were significantly (p<0.05) increased in all the synbiotic combinations compared to unfermented extract, whereas after 48 hrs fermentation all the combinations were significantly higher (p<0.05) than unfermented carob except for L. acidophilus+ 5% and S. thermophilus +2.5%. Overall, there were no significant (p>0.05) differences in ORAC values among most fermented treatments. In addition, B. bifidum 2.5% and S. thermophilus 2.5% 24 hrs incubation showed the highest ORAC values (5.46 and 5.43 uM Trolox g–1, respectively).
DISCUSSION
This research aimed to assess the prebiotic effects of carob and konjac on the growth of different probiotic bacteria. Moreover, phytochemical and antioxidant contents of fermented carob samples were determined. Carob and konjac extracts showed different effects on pH levels and growth of probiotics. Furthermore, fermentation increased phytochemical contents (TPC and TFC) of carob pods and significantly impacted the antioxidant activity of fermented carob extracts.
As indicated, pH levels of synbiotics, i.e., probiotics+carob or konjac, yielded varied results. This could be possibly ascribed to different metabolism system of the different probiotic strains34 utilized in this study and furthermore, the utilization of treatment substrates35. Also, most of probiotic strains showed lower pH values after longer incubation time which could be attributed to more production of buffering organic compounds36. Some strains such as L. acidophilus showed better acidification with increased concentrations of prebiotics, whereas acidification of other strains such as S. thermophilus was not affected. The fermentability of the major fibers in food plays a vital role in its prebiotic properties and accordingly, the acidification rate, which was observed in the results with some treatments.
An increase in growth (Log CFU) of probiotics was observed in majority of the synbiotics systems. Carob was able to increase growth of L. paracasei, B. bifidum and S. thermophilus whereas konjac showed better prebiotic activity with L. paracasei and L. acidophilus. The fermentable fibers in konjac and carob are structurally similar with mannose core while, the difference is in the secondary sugar moiety, glucomannan (konjac) and galactomannan (carob). Some lactic acid bacteria as S. thermophilus are able to metabolize glucose and galactose37, moreover, mannose has shown some prebiotic activity with some probiotic strains such as Lactobacillus and Bifidobacterium bifidum38. In line with the present results, previous studies indicated konjac supplemented to MRS media were shown to increase numbers of B. bifidum NCIMB 700 compared to the strains grown in MRS media alone and others grew in MRS supplemented with pectic-hydrolysate26. In addition, supplementation with glucomannans resulted in an increase in the size of the colonies for the following strains: B. breve NCIMB 702 258, L. acidophilus NCFB 1748 and L. delbrueckii NCFB 1489. In an animal study by Chen et al.39, konjac glucomannan was able to increase probiotic content in cecal microflora in Balb/c mice and led to high production of SCFAs such as acetate and propionate.
The inability of some treatments to increase growth could be attributed to incubation time as it plays a vital role in the growth of probiotics40,41. Although gums are soluble, locust bean gum in carob is favored more by Bifidobacterium and could be attributed to enzyme production. Bifidobacterium strains produce alpha and beta-mannosidase enzymes42, which could be the reason for their fermentation of carob at a better level than the other probiotics16. Probiotics can utilize carob and konjac as a prebiotic source by secreting enzymes such as cellulase and mannanase, which can utilize carbohydrates in these fibers, leading to an increase in growth and/or activity. The increase in Log CFU numbers was not correlated with reduction in pH except for L. paracasei.
Carob seeds and pulp are a rich source for polyphenolics19,27, dietary fibers19,21 and the pod contains a significant amount of the essential amino acids28. Current findings are in line with studies reporting the beneficial effects of polyphenol sources on probiotic bacteria43,44. Total Polyphenol Content (TPC) was determined before and after fermentation in all carob treatments. Unfermented carob was extracted with ethanol, which is a food-grade solvent with a lower polarity index as compared to water45 and is useful in extraction of majority of phenolic compounds. The TPC of unfermented carob extracts were significantly (p<0.05) lower compared to fermented carob extracts. The reason could be due to the metabolism of polyphenols by probiotic strains, separating sugar molecules from the phenolic compounds leading to more absorbed and soluble form of phenolics32. Proteolytic, tyrosinase and laccase enzymes from lactic acid bacteria46 utilized in the study could have been also have contributed to increasing TPC by modifying the molecules structure. B. bifidum and S. thermophilus showed higher TPC content than other strains which maybe result of better galactosidase activity leading to more release of polyphenols. Total Flavonoid Content (TFC) of unfermented carob pods were significantly (p<0.05) lower compared to fermented pods. The acidification resulted from fermentation could have played a role in improving total flavonoid content as low pH is associated with a higher release of flavonoids and accordingly, higher values after fermentation32. Similar to TPC results, B. bifidum and S. thermophilus showed higher TFC content than the two Lactobacillus strains and also lower concentrations of carob (2.5%) showed better results than 5% for both TPC and TFC. In comparison, Kumazawa et al.47 indicated total phenolics extracted from carob with gallic acid as standard was 0.192 g–1, whereas current results showed unfermented carob powder contains total phenols 0.24 g–1. Flavonoids are class of polyphenols, which are found in plants in the form of glycosides and metabolized by action of microorganisms, this could be the reason TFC of the unfermented sample (29 mg CE 100 g–1) was less than the fermented samples.
Carob is rich in the flavonoid quercetin28, which has been shown to be a strong antioxidant and antidiabetic polyphenol. The antioxidant activity of the samples varied depending on the fermentation process and methods of analysis. Unfermented carob showed a DPPH IC50 scavenging (2.14 mg mL–1), which is lower than that of Goulas and Georgiou48, DPPH IC50 scavenging (2.9-4 mg mL–1). The ORAC values of the carob samples ranged from (0.44-5.46 g–1), which is slightly higher than those reported by Mahtout et al.49, in a carob supplemented kefir (0.194-2.418 g–1). The ORAC is considered a preferable antioxidant assay as it mimics the biological system utilizing a natural occurring radical (peroxyl radical) and is also able to detect antioxidant abilities of nonprotein antioxidants50. Flavonoids content is usually associated with high ORAC values, which can be seen in the results as the majority of synbiotics with high flavonoid content showed a significant increase in ORAC values except for S. thermophilus 2.5% after 48 hrs fermentation. All carob samples showed decent FRAP antioxidation potential. FRAP assay is affected by pH levels, gallic acid which is a major polyphenol in carob has a decreased FRAP antioxidant ability with low pH51. This might be the reason why some fermented samples showed low FRAP values. Moreover, FRAP assay underestimates the antioxidant effect of thiol group52 which is highly found in carob pods. In addition, lactic acid bacteria were found to bio-convert methionine an essential amino acid found in carob28 to free thiols53. High TEAC values were observed in unfermented carob and S. thermophilus 2.5%. The reason S. thermophilus 2.5% had a higher TEAC among all the fermented samples, could be attributed to the high total phenolic contents of the samples. Moreover, most of the fermented samples with the lower concentration of carob showed a higher TEAC value which could be attributed to the fact that they showed a higher TPC and TFC. Different TEAC values among fermented samples could also be due to molecular structure modification of phenolic compounds54 by action of different probiotic strains.
Fermentation can improve phenolic content antioxidant abilities of foods in addition to affecting other physiochemical properties such as texture, color and pH. Khan et al.55 found that fermentation with lactic acid bacteria significantly increased total and free phenolic and flavonoid contents in dried longan pulp. Moreover, antioxidant activity (ORAC and FRAP values) was also increased after fermentation. In addition to their abilities to increase phytochemical content of a food, probiotics such as S. thermophilus, L. acidophilus, L. brevis and Bifidobacterium have intracellular postbiotics contents that have an antioxidant activity56. Curiel et al.57 reported a significant increase in phenolic compounds such as gallic acid, vanillic acid, ellagic acid, myricetin and quercetin in Myrtus communis berries after 48 hrs fermentation with Lactobacillus plantarum. Moreover, fermentation time could also play a role in increasing antioxidant activity of food. In a study by Eom et al.58 antioxidant activity (FRAP value and β-carotene oxidation inhibition percentage) was significantly increased by increasing fermentation time as well as total phenolic content of ginseng marc fermented by Pediococcus acidilactici.
CONCLUSION
In conclusion, the utilization of carob and konjac with probiotics as a synbiotic mixture may be reasonable for increasing probiotic count and viability in human and animal body and food products because these compounds and other prebiotics improve probiotics tolerability to environmental factors by maintaining acidic medium, facilitating further fermentation and acting as an extra energy source. Carob powder was able to increase the CFU of Streptococcus thermophilus and Bifidobacterium bifidum, whereas konjac extract increased CFU of Lactobacillus acidophilus and Lactobacillus paracasei. Fermented carob had higher levels of polyphenols and flavonoids compared to unfermented samples. The antioxidative potential of fermented and unfermented carobs was different through different assays.
SIGNIFICANCE STATEMENT
This study discovered the prebiotic effect of carob pods and konjac roots which can be useful in increasing beneficial bacteria in foods and improve their stability. The research showed that lactic acid bacteria fermentation was able to enhance the phytochemical and antioxidant potential of carob pods which can be utilized in the fermented and functional foods industry. The utilization of carob and konjac as prebiotic in the functional foods and nutraceuticals industry may help increase the viability and bioavailability of probiotics during storage and digestion.
ACKNOWLEDGMENTS
This work was funded in part by USDA Capacity Building Grant award 201-38821-27757 and the Alabama A & M University Agricultural Experiment Station Normal AL 35762. We thank Dr. Armitra Jackson-Davis (Alabama A and M University, Normal, AL) for providing access to the microbiology laboratory.
REFERENCES
- Neffe-Skocińska, K., A. Rzepkowska, A. Szydłowska and D. Kołożyn-Krajewska, 2018. Trends and possibilities of the use of probiotics in food production. Altern. Replacement Foods, 2018: 65-94.
CrossRefDirect Link - Markowiak, P. and K. Sliżewska, 2017. Effects of probiotics, prebiotics and synbiotics on human health. Nutrients, Vol. 9, No. 9.
CrossRefDirect Link - Kijmanawat, A., P. Panburana, S. Reutrakul and C. Tangshewinsirikul, 2019. Effects of probiotic supplements on insulin resistance in gestational diabetes mellitus: A double-blind randomized controlled trial. J. Diabetes Investig., 10: 163-170.
CrossRefDirect Link - Eslamparast, T., H. Poustchi, F. Zamani, M. Sharafkhah, R. Malekzadeh and A. Hekmatdoost 2014. Synbiotic supplementation in nonalcoholic fatty liver disease: A randomized, double-blind, placebo-controlled pilot study. Am. J. Clin. Nut. 99: 535-542.
CrossRefDirect Link - Sharafedtinov, K.K., O.A. Plotnikova, R.I. Alexeeva, T.B. Sentsova and E. Songisepp et al., 2013. Hypocaloric diet supplemented with probiotic cheese improves body mass index and blood pressure indices of obese hypertensive patients-a randomized double-blind placebo-controlled pilot study. Nutr. J., Vol. 12.
CrossRefDirect Link - Jung, S., Y.J. Lee, M. Kim, M. Kim and J.H. Kwak et al., 2015. Supplementation with two probiotic strains, Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032, reduced body adiposity and Lp-PLA2 activity in overweight subjects. J. Funct. Foods, 19: 744-752.
CrossRefDirect Link - Jin-Hee Chang, Youn Young Shim, Seong-Kwan Cha, Martin J. T. Reaney, Kew Mahn Chee 2012. Effect of Lactobacillus acidophilus KFRI342 on the development of chemically induced precancerous growths in the rat colon. J. Med. Microbiol., 61: 361-368.
CrossRefDirect Link - Zhang, M., X. Fan, B. Fang, C. Zhu, J. Zhu and F. Ren, 2015. Effects of Lactobacillus salivarius Ren on cancer prevention and intestinal microbiota in 1, 2-dimethylhydrazine-induced rat model. J. Microbiol., 53: 398-405.
CrossRefDirect Link - Roškar, I., K. Švigelj, M. Štempelj, J. Volfand, B. Štabuc, Š. Malovrh and I. Rogelj, 2017. Effects of a probiotic product containing Bifidobacterium animalis subsp. animalis IM386 and Lactobacillus plantarum MP2026 in lactose intolerant individuals: Randomized, placebo-controlled clinical trial. J. Funct. Foods, 35: 1-8.
CrossRefDirect Link - Kang, C.H., S.H. Han, J.S. Kim, Y.G. Kim, Y. Jeong, H.M. Park, N.S. Paek, 2019. Inhibition of nitric oxide production, oxidative stress prevention and probiotic activity of lactic acid bacteria isolated from the human vagina and fermented food. Microorganisms, Vol. 7.
CrossRefDirect Link - Zamani, B., A. Sheikhi, N. Namazi, B. Larijani and L. Azadbakht, 2020. The effects of supplementation with probiotic on biomarkers of oxidative stress in adult subjects: A systematic review and meta-analysis of randomized trials. Probiotics Antimicro. Prot., 12: 102-111.
CrossRefDirect Link - Davani-Davari, D., M. Negahdaripour, I. Karimzadeh, M. Seifan and M. Mohkam et al., 2019. Prebiotics: Definition, types, sources, mechanisms and clinical applications. Foods, Vol. 8.
CrossRefDirect Link - Alexander, C., K.S. Swanson, G.C. Fahey and K.A. Garleb, 2019. Perspective: Physiologic importance of short-chain fatty acids from nondigestible carbohydrate fermentation. Adv. Nut., 10: 576-589.
CrossRefDirect Link - Devaraj, R.D., C.K. Reddy and B. Xu, 2019. Health-promoting effects of konjac glucomannan and its practical applications: A critical review. Int. J. Biol. Macromol., 126: 273-281.
CrossRefDirect Link - Tester, R. and F. Al-Ghazzewi, 2017. Glucomannans and nutrition. Food Hydrocolloids, 68: 246-254.
CrossRefDirect Link - Tungland, B.C. and D. Meyer, 2002. Nondigestible oligo- and polysaccharides (dietary fiber): Their physiology and role in human health and food. Comprehen. Rev. Food Sci. Food Saf., 1: 90-109.
CrossRefDirect Link - Al‐Ghazzewi, F.H., R.F. Tester and K. Alvani, 2012. The synbiotic effects of konjac glucomannan hydrolysates (GMH) and lactobacilli on the growth of Staphylococcus aureus and Salmonella typhimurium. Nut. Food Sci., 42: 97-101.
CrossRefDirect Link - Suwannaporn, P., K. Thepwong, R. Tester, F. Al-Ghazzewi and J. Piggott et al., 2013. Tolerance and nutritional therapy of dietary fibre from konjac glucomannan hydrolysates for patients with inflammatory bowel disease (IBD). Bioact. Carbohyd. Dietary Fibre, 2: 93-98.
CrossRefDirect Link - Avallone, R., M. Plessi, M. Baraldi and A. Monzani, 1997. Determination of chemical composition of carob (Ceratonia siliqua): Protein, fat, carbohydrates and tannins. J. Food Comp. Anal., 10: 166-172.
CrossRef - Maier, H., M. Anderson, C. Karl, K. Magnuson and R.L. Whistler, 1993. Guar, locust bean, tara and fenugreek gums. Indus. Gums, 1993: 181-226.
CrossRefDirect Link - Rtibi, K., M.A. Jabri, S. Selmi, A. Souli and H. Sebai et al., 2015. Carob pods (Ceratonia siliqua L.) inhibit human neutrophils myeloperoxidase and in vitro ROS-scavenging activity. RSC Adv., 5: 84207-84215.
CrossRefDirect Link - Pacheco-Ordaz, R., A. Wall-Medrano, M.G. Goñi, G. Ramos-Clamont-Montfort, J.F. Ayala-Zavala and G.A. González-Aguilar, 2018. Effect of phenolic compounds on the growth of selected probiotic and pathogenic bacteria. Lett. Appl. Microbiol., 66: 25-31.
CrossRefDirect Link - Molino, S., N.A. Casanova, J.Á.R. Henares and M.E.F. Miyakawa, 2019. Natural tannin wood extracts as a potential food ingredient in the food industry. J. Agric. Food Chem., 68: 2836-2848.
CrossRefDirect Link - Majeed, M., S. Majeed, K. Nagabhushanam, S. Arumugam, S. Natarajan, K. Beede and F. Ali, 2018. Galactomannan from Trigonella foenum-graecumL. seed: Prebiotic application and its fermentation by the probiotic Bacillus coagulans strain MTCC 5856. Food Sci. Nutr., 6: 666-673.
CrossRefDirect Link - Al-Ghazzewi, F.H., S. Khanna, R.F. Tester and J. Piggott, 2007. The potential use of hydrolysed konjac glucomannan as a prebiotic. J. Sci. Food Agric., 87: 1758-1766.
CrossRefDirect Link - Stavrou, I.J., A. Christou and C.P. Kapnissi-Christodoulou, 2018. Polyphenols in carobs: A review on their composition, antioxidant capacity and cytotoxic effects and health impact. Food Chem., 269: 355-374.
CrossRefDirect Link - Goulas, V., E. Stylos, M. Chatziathanasiadou, T. Mavromoustakos and A. Tzakos, 2016. Functional components of carob fruit: Linking the chemical and biological space. Int. J. Mol. Sci., Vol. 17.
CrossRefDirect Link - Custódio, L., A.L. Escapa, E. Fernandes, A. Fajardo and R. Aligué et al., 2011. Phytochemical profile, antioxidant and cytotoxic activities of the carob tree (Ceratonia siliqua L.) Germ flour extracts. Plant Foods Hum. Nutr., 66: 78-84.
CrossRefDirect Link - Patel, P., R. Sunkara, L.T. Walker abnd M. Verghese, 2016. Effect of drying techniques on antioxidant capacity of guava fruit. Food Nut. Sci., 07: 544-554.
CrossRefDirect Link - Haile, M. and W. Kang, 2019. Antioxidant activity, total polyphenol, flavonoid and tannin contents of fermented green coffee beans with selected yeasts. Fermentation, Vol. 5.
CrossRefDirect Link - Xing, J., G. Wang, Q. Zhang, X. Liu and Z. Gu et al., 2015. Determining antioxidant activities of lactobacilli cell-free supernatants by cellular antioxidant assay: A comparison with traditional methods. PLoS ONE, Vol. 10.
CrossRefDirect Link - Gao, Y., N. Hamid, N. Gutierrez-Maddox, K. Kantono and E. Kitundu, 2019. Development of a probiotic beverage using breadfruit flour as a substrate. Foods, Vol. 8.
CrossRefDirect Link - Charalampopoulos, D., S.S. Pandiella and C. Webb, 2002. Growth studies of potentially probiotic lactic acid bacteria in cereal-based substrates. J. Applied Microbiol., 92: 851-859.
CrossRefDirect Link - Casarotti, S.N. and A.L.B. Penna, 2015. Acidification profile, probiotic in vitro gastrointestinal tolerance and viability in fermented milk with fruit flours. Int. Dairy J., 41: 1-6.
CrossRefDirect Link - Cui, Y., T. Xu, X. Qu, T. Hu, X. Jiang and C. Zhao, 2016. New insights into various production characteristics of Streptococcus thermophilus strains. Int. J. Mol. Sci., Vol. 17.
CrossRefDirect Link - Korneeva, O.S., I.V. Cheremushkina, A.S. Glushchenko, N.A. Mikhaĭlova, A.P. Baturo, E.E. Romanenko and S.A. Zlygostev, 2012. Prebiotic properties of mannose and its effect on specific resistance. Zh. Mikrobiol. Epidemiol. Immunobiol., 5: 67-70.
Direct Link - Chen, H.L., Y.H. Fan, M.E. Chen and Y. Chan, 2005. Unhydrolyzed and hydrolyzed konjac glucomannans modulated cecal and fecal microflora in Balb/c mice. Nutrition, 21: 1059-1064.
CrossRef - Al-Otaibi, H.S., R. Gashgari, A. Mohammed, S. Almojel, M. Elobeid and J. Abrahaim, 2016. Investigation of the growth ability of probiotic (Lactobacillus and bifidobacterium) in infant’s milk under different environmental conditions. Biomed. Pharmacol. J., 9: 451-462.
CrossRefDirect Link - Ostlie, H.M., H.M. Helland and J.A. Narvhus, 2003. Growth and metabolism of selected strains of probiotic bacteria in milk. Int. J. Food Microbiol., 87: 17-27.
CrossRef - O'Callaghan, A. and D. van Sinderen, 2016. Bifidobacteria and their role as members of the human gut microbiota. Front. Microbiol., Vol. 7.
CrossRefDirect Link - Bialonska, D., S.G. Kasimsetty, K.K. Schrader and D. Ferreira, 2009. The effect of pomegranate (Punica granatum L.) byproducts and ellagitannins on the growth of human gut bacteria. J. Agric. Food Chem., 57: 8344-8349.
CrossRefDirect Link - China, R., S. Mukherjee, S. Sen, S. Bose and S. Datta et al., 2012. Antimicrobial activity of Sesbania grandiflora flower polyphenol extracts on some pathogenic bacteria and growth stimulatory effect on the probiotic organism Lactobacillus acidophilus. Microbiol. Res., 167: 500-506.
CrossRefDirect Link - Kleiman, M., K.A. Ryu and A.P. Esser-Kahn, 2016. Determination of factors influencing the wet etching of polydimethylsiloxane using tetra-n-butylammonium fluoride. Macromol. Chem. Phys., 217: 284-291.
CrossRefDirect Link - Matthews, A., A. Grimaldi, M. Walker, E. Bartowsky, P. Grbin and V. Jiranek, 2004. Lactic acid bacteria as a potential source of enzymes for use in vinification. Appl. Environ. Microbiol., 70: 5715-5731.
CrossRefDirect Link - Kumazawa, S., M. Taniguchi, Y. Suzuki, M. Shimura, M.S. Kwon and T. Nakayama, 2002. Antioxidant activity of polyphenols in carob pods. J. Agric. Food Chem., 50: 373-377.
CrossRefDirect Link - Goulas, V. and E. Georgiou, 2019. Utilization of carob fruit as sources of phenolic compounds with antioxidant potential: extraction optimization and application in food models. Foods, Vol. 9.
CrossRefDirect Link - Mahtout, R., F. Zaidi, L. Saadi, S. Boudjou, B.D. Oomah and F. Hosseinian, 2016. Carob (Ceratoniasiliqua L.) supplementation affects kefir quality and antioxidant capacity during storage. Int. J. Eng. Techn., 2: 168-177.
Direct Link - Cao, G., H.M. Alessio and R.G. Cutler, 1993. Oxygen-radical absorbance capacity assay for antioxidants. Free Radic. Biol. Med., 14: 303-311.
CrossRefDirect Link - Wong, C.W., W.S.M. Cheung, Y.Y. Lau, A.A.S.B. de la Torre and R. Owusu-Apenten, 2015. A FRAP assay at pH 7 unveils extra antioxidant activity from green, black, white and rooibos tea but not apple tea. F. Nut. Reprt., 1: 16-23.
CrossRefDirect Link - Payne, A.C., A. Mazzer, G.J.J. Clarkson and G. Taylor, 2013. Antioxidant assays-consistent findings from FRAP and ORAC reveal a negative impact of organic cultivation on antioxidant potential in spinach but not watercress or rocket leaves. Food Sci. Nutr., 1: 439-444.
CrossRefDirect Link - Dias, B. and B. Weimer, 1998. Conversion of methionine to thiols by lactococci, lactobacilli and brevibacteria. Appl. Environ. Microbiol., 64: 3320-3326.
CrossRefDirect Link - Kwaw, E., Y. Ma, W. Tchabo, M.T. Apaliya and M. Wu et al., 2018. Effect of lactobacillus strains on phenolic profile, color attributes and antioxidant activities of lactic-acid-fermented mulberry juice. Food Chem., 250: 148-154.
CrossRefDirect Link - Khan, S.A., L. Liu, T. Lai, R. Zhang and Z. Wei et al., 2018. Phenolic profile, free amino acids composition and antioxidant potential of dried longan fermented by lactic acid bacteria. J. Food Sci. Technol., 55: 4782-4791.
CrossRefDirect Link - Aguilar-Toalá, J.E., R. Garcia-Varela, H.S. Garcia, V. Mata-Haro, A.F. González-Córdova, B. Vallejo-Cordoba and A. Hernández-Mendoza, 2018. Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Technol., 75: 105-114.
CrossRefDirect Link - Curiel, J.A., D. Pinto, B. Marzani, P. Filannino, G.A. Farris, M. Gobbetti and C.G. Rizzello, 2015. Lactic acid fermentation as a tool to enhance the antioxidant properties of Myrtus communis berries. Microb. Cell Fact., Vol. 14.
CrossRefDirect Link - Eom, S.J., J.E. Hwang, K.T. Kim and H.D. Paik, 2018. Increased antioxidative and nitric oxide scavenging activity of ginseng marc fermented by Pediococcus acidilactici KCCM11614P. Food Sci. Biotechnol., 27: 185-191.
CrossRefDirect Link