
J.A. Serna-Jimenez
Agroindustrial Processes Research Group, Universidad de La Sabana, Campus del Puente del Comun Km 7, Chia, Cundinamarca, Colombia
M.X. Quintanilla-Carvajal
Agroindustrial Processes Research Group, Universidad de La Sabana, Campus del Puente del Comun Km 7, Chia, Cundinamarca, Colombia
J.M. Rodriguez
Agroindustrial Processes Research Group, Universidad de La Sabana, Campus del Puente del Comun Km 7, Chia, Cundinamarca, Colombia
M.A. Uribe
Agroindustrial Processes Research Group, Universidad de La Sabana, Campus del Puente del Comun Km 7, Chia, Cundinamarca, Colombia
B. Klotz
Alpina Research Institute, Sopo, Cundinamarca, Colombia
American Journal of Food Technology
Year: 2016 | Volume: 11 | Issue: 1-2 | Page No.: 21-28







ABSTRACT
Fungi are the major cause of fresh fermented dairy spoilage adversely affecting the economic performance of the dairy industry. The control of the contamination and the inhibition of the fungal growth is a constant challenge during the production process. The development of natural and effective procedures that guarantee food quality is nowadays an imperative. However, the successful application of the hurdle technology demands a better understanding of the behaviour of microorganisms in relation to the most relevant preservation factors. In this study the effect of temperature, pH and bioprotectants on the growth of Mucor circinelloides was investigated. The mold was isolated from deteriorated yogurt and identified by molecular techniques. Mucor circinelloides was inoculated in potato dextrose agar at two different pH levels (4.5 and 5.8) and five different temperatures (4, 12, 21, 29 and 37°C). The growth was evaluated by a factorial design at different combinations of pH and temperature. The results demonstrated that temperature and pH and the interaction between them were the factors that controlled the growth rate of the microorganism. At pH 4.5 and 21°C, the mycelium growth rate presented the highest growth rate and no evident apparent lag phase. Biological control with the addition of lactic acid bacteria and their metabolites as bioprotectants resulted in a reduction of up to 50% in the mycelial growth rate of M. circinelloides. The present study provides useful information of the kinetic growth parameters of M. circinelloides under different pH and temperatures values and presence of bioprotective agents. The results of the study showed the biocontrol potential of lactic acid bacteria and their metabolites on the growth of molds.
PDF Abstract XML References Citation
Received: July 22, 2015;
Accepted: November 04, 2015;
Published: February 15, 2016
How to cite this article
J.A. Serna-Jimenez, M.X. Quintanilla-Carvajal, J.M. Rodriguez, M.A. Uribe and B. Klotz, 2016. Development of a Combined Temperature and pH Model and the Use of Bioprotectants to Control of Mucor circinelloides. American Journal of Food Technology, 11: 21-28.
DOI: 10.3923/ajft.2016.21.28
URL: https://scialert.net/abstract/?doi=ajft.2016.21.28
DOI: 10.3923/ajft.2016.21.28
URL: https://scialert.net/abstract/?doi=ajft.2016.21.28
INTRODUCTION
Fungi are ubiquitous in nature and have evolved over time to colonize a wide range of ecosystems, including foods. Airborne transfer of fungal spores is now seen as a significant route of contamination in many sectors of the food industry (Stringini et al., 2008). If environmental conditions allow fungal growth, their colonization of foods results in spoiled products and subsequent significant economic losses. Fungal presence in food may adversely affect not only the organoleptic value of the product but, most importantly, its nutritional value and shelf life (Gougouli et al., 2011).
The development of visible mycelia is one of the most frequent quality problems during shelf life in the production of fresh fermented products and has important economic implications for the dairy industry. The refrigerated storage of fresh fermented dairy products is usually not sufficient to prevent spore germination and mycelium growth because most fungal species are psychotrophic and able to grow at low temperatures (Gougouli and Koutsoumanis, 2013). Consequently, the development of control programs and monitoring techniques are necessary to minimize the effects of contamination.
Fungal growth involves spore germination, hyphal extension, formation of mycelia and finally sporulation. Spores can be disseminated in the facility environment and they are principally responsible for the spoilage dairy products. Under favorable substrates and environmental conditions, spores start germination with a swelling process. Thereafter, when the length of the germ tube is between one half and twice the spore diameter, the spore is considered to have germinated (Dantigny et al., 2005). Filamentous fungi are highly adaptable to extreme conditions indicated by their ability to grow easily on substrates that are hostile to other microorganisms, surviving in a wide range of temperatures and in media with high sugar concentrations high acidity (Gougouli and Koutsoumanis, 2013). Mucor spp. and Penicillium spp., have been identified as fungal contaminants of food period Mucor spp., is considered the major food contaminant having the ability to grow in high acid and low temperature environments as well as causing the sensorial deterioration of food products (Gougouli and Koutsoumanis, 2013).
The addition of preservatives to control mold growth has been criticized as consumers demand more natural food products (Dantigny et al., 2005). The use of the natural microflora in the food or microflora added to the food as well as its antimicrobial products to extend the shelf life and safety is a biopreservation practice (Schnurer and Magnusson, 2005). Bioprotectants are currently used for the bioprotection of food preventing specially the growth of pathogenic bacteria (Schnurer and Magnusson, 2005; Garcia et al., 2010; Delavenne et al., 2013). The most common bioprotectants are Lactic Acid Bacteria (LAB) cocktails and their metabolic products, which can also inhibit the growth of molds and yeast. De Muynck et al. (2004) mentioned the antifungal properties of LAB between 1-75%, present antifungal properties and are morphologically and physiologically heterogeneous. These LAB are Generally Recognized As Safe (GRAS) and Qualified Presumption of Safety (QPS). Lactic Acid Bacteria (LAB) are Gram positive and their principal characteristic is the production of lactic acid as the major product of the fermentative metabolism of carbohydrates. In addition, they can produce other substances such as organic acids, peroxide, bacteriocins and other acids with antifungal activity (Schnurer and Magnusson, 2005; Delavenne et al., 2013).
The application of predictive mycology in foods is a field of research in the last decade (Dantigny et al., 2005). The development of modeling tools to predict fungal behavior relies on quantitative data concerning the relative impact of various factors on fungal growth and the germination of fungal spores (Gougouli and Koutsoumanis, 2010). Kinetic models to determine microbial responses in relation to time and environmental conditions and to provide estimates of relevant growth parameters (lag phase (λ) and growth rate (μ)) is a useful tool in food quality and safety control systems. Most of these models are empirical, usually polynomials. Although many other factors in foods affect microbial growth, they account for a small proportion of the total variation in growth compared with the main factors, such as temperature, activity water, pH and the presence of preservatives (Gougouli and Koutsoumanis, 2010). Other factors such as the quality of spores, their ability to germinate and form hyphae and the concentration of spores in the inoculation media might also influence the fungal growth (Bizukojc and Ledakowicz, 2006).
The ability to predict mold growth is fairly more complicated than for bacteria. Molds are not unicellular and thus grow more slowly and in a different way forming surface colonies and hyphae throughout the food (Gougouli and Koutsoumanis, 2010). The growth of different species of Penicillium and Aspergillus has been described using predictive models by Gougouli et al. (2011), Membre et al. (2001), Valik et al. (1999) and Gibson and Hocking (1997). However, predictive mycology studies in the context of biopreservation are not as abundant as bacterial (Henao-Ardila et al., 2015).
Mucor circinelloides can be found in the environment of dairy plants threatening to contaminate goods. Compared to the number of food safety and quality studies concerning pathogenic bacteria, fungal studies are more limited. Thus, the aim of the present study was to determine first the mycelium growth kinetics of M. circinelloides isolated from yogurt at different temperatures and pH values and then, the effect of bioprotectants on the mycelial growth rate of this fungal strain.
MATERIALS AND METHODS
Microorganism isolation and identification: The mold, M. circinelloides was isolated from deteriorated commercial yogurts and molecular techniques were used for the identification. For ADN isolation, a kit from Invitrogen®, the PureLink Plant Total DNA Purification Kit was used according to the standard protocol. The PCR reactions were performed with specific primers: 5’-TCCGTAGGTGAACCT GCG G-3’ (ITS1 region) and 5’-TCC TCCGCTTATTGATATGC-3’ (ITS4 region) (Diaz et al., 2000). The reaction was performed using a thermal cycler (BioRad®, Hercules, California, USA). The PCR reaction was developed by standard procedures. Standard reaction mixtures were prepared with a volume of 50 μL from a mixture containing 15 μL of PCR water, 20 μL of PCR buffer, 3.2 μL of dNTPs, 3.2 μL of Taq DNA polymerase (BioLine®, United Kingdom), 12 μL of MgCl2 and 5.6 μL of each primer. The reaction cycle was performed as follows; a denaturation cycle at 94°C for 5 min and the following amplification conditions for 30 cycles: denaturation at 94°C for 30 sec, annealing at 58°C for 30 sec and extension at 72°C for 30 sec. Lastly, there was a cycle at 72°C for 7 min.
The amplicons of RAPD-PCR were separated by electrophoresis at 90 V on agarose gels (BioLine®, United Kingdom) in Tris Borato EDTA (TBE 1X) using a horizontal electrophoresis system (BioRad®, Hercules, California, USA). The gels were stained with ethidium bromide, Sybr safe DNA gel stain (Invitrogen, USA), visualized under UV light and digitalized using Universal Hood II (BioRad®, Hercules, California, USA). Hyperladder II (BioLine®, United Kingdom) was used as a marker. The resulting bands were isolated from the gels and sequenced for the identification of the microorganisms.
The PCR product was sequenced and electrophoretogram was edited with Chromas Lite® (http://technelysium.com.au). The sequence was evaluated using the Mycobank database (http://www.mycobank.org/).
Fungal inoculum: Spore suspensions of M. circinelloides were obtained from two week old cultures grown on Potato Dextrose Agar (PDA, Scharlau, Barcelona-Spain, Code: 01 483) at 29°C by adding sterile water. These suspensions were shaken in a vortex to separate spore aggregates. Spore counts were determined by using a Neubauer hemocytometer and adjusted to a concentration of 102 spores/mL (Bosch et al., 1995).
Growth curves at different temperatures and pH: A factorial experimental design was proposed to describe the growth rates of the microorganism at different pH levels (4.5 and 5.8) at optimum pH reported by Dantigny et al. (2005) and temperatures (4, 12, 21, 29 and 37°C) using a commercial statistical package, Design Expert version 8®(Statease Inc., Minneapolis, U.S.A.). Independent assays were run in triplicate and the order of experiments was completely random.
The standard growth medium used in all experiments was PDA. Lactic acid (Merck and Co., Whitehouse Station, N.J., U.S.A.) was added to lower the pH to the value of 4.5. The pH of the media was determined at 25°C before and after inoculation by using a pH meter with a glass electrode (Methrom 888 Titrando, Metrohm, Florida, USA).
Petri dishes (diameter = 90 mm) containing 30 mL of PDA or acidified PDA were inoculated in the center with one 60 μL droplet each of a dense spore suspension of M. circinelloides (102 spores/mL) and were incubated in programmable incubators (Friocell, MMM Medcenter- Einrichtungen GmbH, Germany). The temperatures to be evaluated in the study represent: 4°C, refrigeration temperature (Gougouli and Koutsoumanis, 2010) 12°C, temperature of abuse for dairy products (Gougouli and Koutsoumanis, 2010; Gougouli et al., 2011) 21 and 29°C, optimal temperature for mold growth (Gougouli and Koutsoumanis, 2010) and 37°C, maximum temperature for mold growth (Gougouli et al., 2011).
The mycelial diameter of the inoculated petri dishes was measured every 24, 48, 72, 96 and 120 h using a Colony Counter ProtoCOL 3 (Synoptic, Frederick, Md, U.S). The mycelial growth rate (μ) was calculated using the methodology reported by Gifford and Schoustra (2013). Two perpendicular measurements were evaluated to obtain the final diameter of each colony. The growth rate (μ) (mm h1–) was estimated as half of the slope of the regression line of colony diameter versus time. The apparent lag time (λ) (h1–) was calculated by an equation reported by Dantigny et al. (2005).
Effect of bioprotectants on mycelial growth rate: The bioprotectants used in this study were MicroGARD™ 100, (cultured skim milk) MicroGARD™ 200 (cultured dextrose) (Danisco, Niebüll, Germany), Lyofast™ FPR2 and Lyofast™ LRB (SACCO, Cadorago, Italy). The MicroGARD products consist of fermentation metabolites and the Lyofast ones are protective cultures of Lactobacillus rhamnosus, Lactobacillus plantarum and Enterococcus faecium. LyofastTM FPR2 contains there species and LyofastTM LRB only L. rhamnosus.
The suspension of the commercial bioprotectants was prepared according to the product’s description in Man Rogosa Sharpe Broth (MRS Broth. Code: 02-135, Scharlau, Barcelona, Spain). Protective cultures were incubated overnight (12 h) at 37°C. Following incubation, the cultures were stored at a cooling temperature (4°C) at a concentration of 108 CFU mL1–.
The inhibition as the reduction of growth diameter was measured using the methodology of Munoz et al. (2010), with some modifications. Five milliliter of the cultures suspension of 3 and 4 at a concentration of 108 CFU mL1– cocktail were mixed with 20 mL of liquid MRS agar and poured into petri dishes. In the case of 1 and 2 the suspensions were prepared following the technical specifications of the products in MRS broth with incubation 12 h at 37°C. The plates were incubated overnight (24 h) at 37°C. The PDA agar overlaid the MRS agar. Wells were made in the center of the overlaid PDA and were each filled with 60 μL of mold inoculum at a concentration of 102 spores mL1–. The plates were incubated for 120 h at 29°C.
This study was developed in 2013 with duration of 6 months.
RESULTS AND DISCUSSION
Molecular identification: The results showed a 100% identity for M. circinelloides. The Mucor spp., belongs to the class Zygomycetes and is considered the major food contaminant, with the ability to grow in high acid and low temperature environments and to cause the sensorial deterioration of the product. Molds can grow in moist environments found in dairy plants and establish on ceilings, walls and floor drains contaminating the air and other surfaces like packaging material, equipment and the food product (Beletsiotis et al., 2011).
Growth model: The different temperature pH combinations had an effect on the growth of M. circinelloides. The size of the diameter of the mycelia in response to the different conditions obtained from the experimental design is presented in Table 1. The experimental data were used to calculate the coefficients of the quadratic equation.
An ANOVA showed that the resulting models adequately represented the experimental data, with a R2 of 0.99 for M. circinelloides, the predicted R2 value of the model was 0.98. It has been suggested that a good thing model should have an R2 of no less than 0.80.
| Table 1: | Experimental design conditions and growth of Mucor circinelloides |
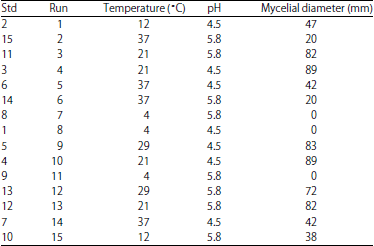 | |
| Table 2: | Factors and predictive equations to estimate the growth of Mucor circinelloides |
 | |
| D: Diameter growth prediction, A: Temperature factor, B: pH factor, µ: Growth rate | |
When the R2 value is close to 1, the empirical model is suitable for fitting the actual data (Little and Hills, 1978; Rodriguez-Bernal et al., 2015). For any of the terms in the model, a large regression co-efficient and a small p-value would indicate a more significant effect on the response variables, the Model F-value was 4510.77 and implies that the model was significant. The factors temperature, pH and interaction between temperature and pH were significant model factors. Table 2 shows the equations for the codified factors, "A"-"B" and the equations used to predict the diameter growth at both pH 4.5 and 5.8.
Equation 2 and 3 can be used for a diameter growth prediction between 4 and 37°C and are useful tools for determining the control of the growth of the mold. Using the model to obtain the best prediction, it indicated that the minimum temperature for growth in M. circinelloides was -7.67°C and the optimum temperature for growth was from 21-29°C. These results are consistent with previously reported articles. Sautour et al. (2002) and Gougouli et al. (2011) reported an optimum growth temperature range of 25-29°C and a minimum growth temperature range of -3 to -4°C for Mucor spp.
The combination of temperature and pH was considered optimum for growth control if the growth inhibition in millimeter reached minimum values. The values of the response at different combinations of temperature and pH were converted to a desirability function. The minimum and maximum desirability values were 0 and 1, respectively. The maximum desirability function obtained was for pH 5.8 and 4°C.
 |
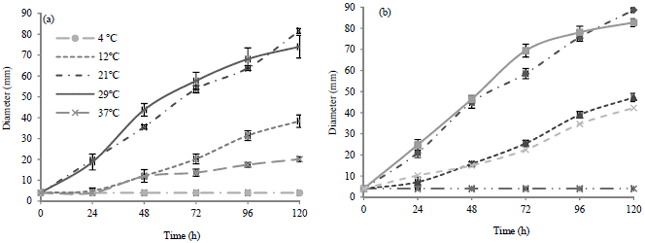
| Fig. 1(a-b): | Mycelial growth of Mucor circinelloides PDA at different temperatures at pH (a) 4.5 and (b) 5.8 |
Figure 1 shows the images of M. circinelloides during 120 h of incubation at the five evaluated temperatures and two pH levels.
The images show the effect in the mycelial morphology at 12 and 37°C. At pH 5.8 the diameters of mycelia were smaller compared to pH 4.5, at pH 4.5 the mold growth increased. Images also confirm the optimal growth conditions for the mycelial growth of M. circinelloides at 21 and 29°C in an acidic media, pH 4.5.
The results obtained showed that the pH and the temperature influenced the growth of M. circinelloides and that their appropriate combination can be used to inhibit or retard the growth of the mold in order to reduce product losses and economic impacts of fungal contamination (Gougouli et al., 2011; In't Veld, 1996). However, as mentioned by McKellar and Hawke (2006) and Gifford and Schoustra (2013), filamentous fungi are adaptive microorganisms that can form spores and hyphae under hostile temperature and pH conditions making necessary additional growth inhibition barriers.
Estimates of Mycelium Growth Rate (MGR): Using an empirical description of colony growth on solid medium, it estimated the mycelia growth model for the filamentous fungus M. circinelloides based on physiological growth parameters. The mycelium growth rate (μ) was estimated using a linear regression of colony diameter over time at the three temperatures and two pH levels. Figure 2 shows the change in diameter at the different times evaluated. The term μ was determined as half of the slope of the regression line (Gifford and Schoustra, 2013).
The mycelium growth data for M. circinelloides in PDA at pH 5.8 is shown in Table 3. At pH 5.8, after an initial lag period, the increase in mycelium diameter over time was a straight line for temperatures tested. At pH 4.5, the lag phase was shorter and the adaptation was higher. The estimated growth kinetic parameters for the tested mold at the different storage temperatures and pH levels are presented in Table 3. The lowest storage temperature at which growth was observed was 12°C. At 37°C the mycelium presented a different morphology (white and yellow without spore formation). At last temperature, a very slow increase of the mycelium diameter was observed after a lag period of about 3 h. The maximum temperature at which growth occurred was between 21 and 29°C at these temperatures, the mycelia presented a typical morphology (green with spore formation). At the refrigeration temperature (4°C) the mold was not able to grow during the time evaluated. The growth parameters were determined with high accuracy by using the linear model as described in Table 3. The regression coefficients were in the range of 0.90-0.98 for pH 5.8 and 0.93-0.99 for pH 4.5. The above linearity of fungal growth has been reported in several studies for different fungi (Gougouli and Koutsoumanis, 2013; Gougouli et al., 2011; Gifford and Schoustra, 2013).
The relationship between the growth rate and temperature are shown in Fig. 3. There was a pH effect over the growth rates. At pH 4.5 the growth rates were higher than the growth rates at pH 5.8. Additionally, the optimum temperatures were consistent with the results obtained by the factorial design (between 21 and 29°C), where the μ values were nearly 0.4 mm h1– at pH 4.5. Gougouli et al. (2011) reported higher μ values for M. circinelloides (1.530 mm h1–) isolated from the air of a yogurt production unit using medium malt extract agar acidified with lactic acid. However, it is well recognized that microbial growth rates are influenced by extrinsic and intrinsic factors and are strain specific.
The lag phase was not present in all cases at 21 and 29°C the lag phase for the two pH levels evaluated was not present.
 |
| Fig. 2(a-b): | Relationship between colony diameter and time of growth for two pH levels (a) 4.5 and 5.8 and (b) Five temperatures (4, 12, 21, 29 and 37°C). Points are the average diameter of 6 replicates |
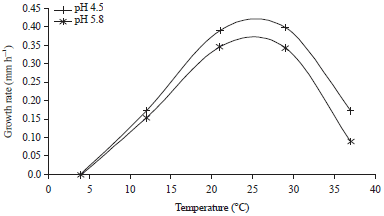 |
| Fig. 3: | Effect of temperature and pH on the growth rate of Mucor circinelloides |
| Table 3: | Growth kinetic parametersof Mucor circinelloidesat different temperatures calculated and linear regression. |
 | |
| ±: Corresponding to the standard error of the diameter measured | |
In the case of acidic pH, the lag phase was shorter than at a pH of 5.8, at which the longest lag phase that occurred was 3 h for 21°C. The lag phase is necessary for the microorganisms to adapt to the environmental conditions to survive and grow. Gougouli et al. (2011) reported that 37°C was the maximum temperature at which the growth of Mucor spp., can be observed, however, M. circinelloides continued to grow at this temperature.
Effect of bioprotectants on mycelial growth rate: The results of the obtained growth rate of M. circinelloides using bioprotectants can be observed in Table 4. Important differences were presented between the μ with and without bioprotectants evaluated at the optimal conditions for mycelial growth (21°C and pH 4.5).
Growth rates dropped dramatically showing an inhibitory effect of 45-47% on the mycelial growth rate using MicroGARD 100™ and Lyofast™ LRB, respectively.
| Table 4: | Growth kinetic parameters of Mucor circinelloides in the presence of bioprotectants and linear regression MicroGARD 100™ (1), MicroGARD™ 200 (2), Lyofast™ FPR2 (3) and Lyofast™ LRB (4) |
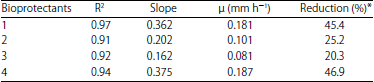 | |
| *Control 0.399 mm h1–, µ: Growth rate | |
MicroGARD™ 200 and Lyofast™ FPR2 also reduced mycelial growth by 25 and 20%, respectively.
Few studies have been reported about the application of LAB and their metabolites as bioprotective agents for control of moulds and yeast in fresh fermented dairy. These bacteria compete with other species by acidifying their environment and rapidly depleting the nutrients in the environment. In addition to these relatively simple antagonistic mechanisms, some LAB also produce potent a ntimicrobial compounds via complex secondary metabolism pathways. Among these are bacteriocins (e.g., nisin), antibiotics (e.g., reutericyclin) and small antibiotic-like molecules, such as reuterin (De Muynck et al., 2004). Most of the antifungal capacity of the LAB described in these reports is due to the production of an antifungal protein or proteinaceous compound (De Muynck et al., 2004; Schnurer and Magnusson, 2005). Other bacteria, such as L. plantarum and L. sanfrancisco, produce special organic acids (3-phenyl-L-lactic acid and caproic acid, respectively) that have antifungal properties (De Muynck et al., 2004), Delavenne et al. (2013) and Gerez et al. (2013) reported on the antifungal activity of LAB. The use of LAB increases the lifetime of dairy fermented beverages due to their production of enzymes and proteolytic exopolysaccharide (Welman and Maddox, 2003) and several secondary metabolites that behave like antimicrobials (Leroy and De Vuyst, 2007). Organic acids can penetrate the microbial cell membrane in their undissociated form, ideally at a pH beneath their pKa. Inside the cell, organic acids dissociate and lower the pH of the cytoplasm causing metabolic disorders (De Muynck et al., 2004; Gougouli and Koutsoumanis, 2010; Leroy and De Vuyst, 2007).
CONCLUSION
The application of the hurdle technology with inclusion of bioprotectants is an ecological and consumer-friendly approach to improve food safety and shelf-life with great potential in the dairy industry. In this study, we demonstrated that the temperature and the pH influenced the growth behavior of M. circinelloides. Low temperatures and pH values decreased the growth of the mold. The commercial bioprotectants assessed were effective reducing the mycelial growth rate at optimal conditions up to 50%.
ACKNOWLEDGMENTS
This study was conducted with the financial support of the Alpina Product Aliembticios S-A in the frame work of R and D incentives of the Administrative Department of Science, Technology and Innovation (COLCIENCIAS, Colombia). We thank Instituto Aplina de Investigación for providing us with access to useful information for this study.
REFERENCES
- Bosch, A., R.A. Maronna and O.M. Yantorno, 1995. A simple descriptive model of filamentous fungi spore germination. Process Biochem., 30: 599-606.
CrossRefDirect Link - Beletsiotis, E., D. Ghikas and K. Kalantzi, 2011. Incorporation of microbiological and molecular methods in HACCP monitoring scheme of molds and yeasts in a Greek dairy plant: A case study. Procedia Food Sci., 1: 1051-1059.
CrossRefDirect Link - Bizukojc, M. and S. Ledakowicz, 2006. A kinetic model to predict biomass content for Aspergillus niger germinating spores in the submerged culture. Process Biochem., 41: 1063-1071.
CrossRefDirect Link - Dantigny, P., A. Guilmart and M. Bensoussan, 2005. Basis of predictive mycology. Int. J. Food Microbiol., 100: 187-196.
CrossRefDirect Link - De Muynck, C., A.I.J. Leroy, S. De Maeseneire, F. Arnaut, W. Soetaert and E.J. Vandamme, 2004. Potential of selected lactic acid bacteria to produce food compatible antifungal metabolites. Microbiol. Res., 159: 339-346.
CrossRefDirect Link - Delavenne, E., R. Ismail, A. Pawtowski, J. Mounier, G. Barbier and G. Le Blay, 2013. Assessment of lactobacilli strains as yogurt bioprotective cultures. Food Control, 30: 206-213.
CrossRefDirect Link - Diaz, M.R., T. Boekhout, B. Theelen and J.W. Fell, 2000. Molecular sequence analyses of the intergenic spacer (IGS) associated with rDNA of the two varieties of the pathogenic yeast, Cryptococcus neoformans. Syst. Applied Microbiol., 23: 535-545.
CrossRefDirect Link - Garcia, D., A.J. Ramos, V. Sanchis and S. Marin, 2010. Modelling mould growth under suboptimal environmental conditions and inoculum size. Food Microbiol., 27: 909-917.
CrossRefDirect Link - Gerez, C.L., M.J. Torres, G.F. de Valdez and G. Rollan, 2013. Control of spoilage fungi by lactic acid bacteria. Biol. Control, 64: 231-237.
CrossRefDirect Link - Gifford, D.R. and S.E. Schoustra, 2013. Modelling colony population growth in the filamentous fungus Aspergillus nidulans. J. Theor. Biol., 320: 124-130.
CrossRefDirect Link - Gougouli, M., K. Kalantzi, E. Beletsiotis and K.P. Koutsoumanis, 2011. Development and application of predictive models for fungal growth as tools to improve quality control in yogurt production. Food Microbiol., 28: 1453-1462.
CrossRefDirect Link - Gougouli, M. and K.P. Koutsoumanis, 2010. Modelling growth of Penicillium expansum and Aspergillus niger at constant and fluctuating temperature conditions. Int. J. Food Microbiol., 140: 254-262.
CrossRefDirect Link - Gougouli, M. and K.P. Koutsoumanis, 2013. Relation between germination and mycelium growth of individual fungal spores. Int. J. Food Microbiol., 161: 231-239.
CrossRefDirect Link - In't Veld, J.H.J.H., 1996. Microbial and biochemical spoilage of foods: An overview. Int. J. Food Microbiol., 33: 1-18.
CrossRefDirect Link - Leroy, F. and L. De Vuyst, 2007. Modelling microbial interactions in foods. Modell. Microorganisms Food, 11: 214-227.
Direct Link - Valik, L., J. Baranyi and F. Gorner, 1999. Predicting fungal growth: The effect of water activity on Penicillium roqueforti. Int. J. Food Microbiol., 47: 141-146.
CrossRefDirect Link - McKellar, R.C. and A. Hawke, 2006. Assessment of distributions for fitting lag times of individual cells in bacterial populations. Int. J. Food Microbiol., 106: 169-175.
CrossRefDirect Link - Membre, J.M., M. Kubaczka and C. Chene, 2001. Growth rate and growth-no-growth interface of Penicillium brevicompactum as functions of pH and preservative acids. Food Microbiol., 18: 531-538.
CrossRefDirect Link - Gibson, A.M. and A.D. Hocking, 1997. Advances in the predictive modelling of fungal growth in food. Trends Food Sci. Technol., 8: 353-358.
CrossRefDirect Link - Munoz, R., M.E. Arena, J. Silva and S.N. Gonzalez, 2010. Inhibition of mycotoxin-producing Aspergillus nomius VSC 23 by lactic acid bacteria and Saccharomyces cerevisiae. Braz. J. Microbiol., 41: 1019-1026.
CrossRefDirect Link - Sautour, M., C.S. Mansur, C. Divies, M. Bensoussan and P. Dantigny, 2002. Comparison of the effects of temperature and water activity on growth rate of food spoilage moulds. J. Ind. Microbiol. Biotechnol., 28: 311-315.
CrossRefDirect Link - Schnurer, J. and J. Magnusson, 2005. Antifungal lactic acid bacteria as biopreservatives. Trends. Food Sci. Technol., 16: 70-78.
CrossRefDirect Link - Stringini, M., F. Comitini, M. Taccari and M. Ciani, 2008. Yeast diversity in crop-growing environments in Cameroon. Int. J. Food Microbiol., 127: 184-189.
CrossRefDirect Link - Welman, A.D. and I.S. Maddox, 2003. Exopolysaccharides from lactic acid bacteria: Perspectives and challenges. Trends Biotechnol., 21: 269-274.
CrossRefDirect Link - Little, T.M. and F.J. Hills, 1978. Agricultural Experimentation: Design and Analysis. John Wiley and Sons, New York, United State, ISBN: 9780471023524, Pages: 350.
Direct Link - Rodriguez-Bernal, J.M., E. Tello, E. Flores-Andrade, M. de Jesus Perea-Flores, A.A. Vallejo-Cardona, G.F. Gutierrez-Lopez and M.X. Quintanilla-Carvajal, 2015. Effect of borojo (Borojoa patinoi Cuatrecasas) three-phase composition and gum Arabic on the glass transition temperature. J. Sci. Food Agric., (In Press).
CrossRefDirect Link