
U.O. George-Okafor
Department of Applied Microbiology and Brewing, Enugu State University of Science and Technology, Enugu, Nigeria
LiveDNA: 234.23250
ORCID: 0000-0001-8932-6442
F.O. Tasie
Department of Applied Microbiology and Brewing, Enugu State University of Science and Technology, Enugu, Nigeria
N.C. Anyamene
Department of Food Science and Technology, Nnamdi Azikiwe University, Awka, Anambra, Nigeria
American Journal of Food Technology
Year: 2013 | Volume: 8 | Issue: 1 | Page No.: 54-64







ABSTRACT
The need for starch-converting enzymes in various industries has led to enormous interest in their production at a cheap rate. The present study was focused on the examination of various inexpensive substrates and other production parameters for amylase production by the most potential selected strain. It was observed that A. oryzae-SR2 was the best amylase producer (36.31±0.24 mm) among other Aspergillus isolates when screened by starch hydrolysis test. Preliminary submerged fermentation using soluble starch medium confirmed the potentials of A. oryzae-SR2 for amylase production. The maximum enzyme production under submerged fermentation was achieved with 0.5% sugar-cane juice supplemented with 0.5% dry-flakes fermented tapioca, 1% yeast extract, 7% inoculums concentration, pH 7.0, temperature of 55°C and incubation period of 48 h. The enzyme demonstrated strong hydrolytic activities (56.4%) on the tested starchy foods with cassava starch being the most hydrolyzed (101.5%) with 86.90±1.10% conversion efficiency at 70°C. The ability to produce high amount of amylase within a relatively short time (48 h) using cheap substrates and the ability of its enzyme to effectively hydrolyze starchy foods suggests its suitability for biotechnological application especially in starch-converting industries.
PDF Abstract XML References Citation
Received: November 24, 2012;
Accepted: January 14, 2013;
Published: April 13, 2013
How to cite this article
U.O. George-Okafor, F.O. Tasie and N.C. Anyamene, 2013. Studies on Low-cost Substrates and Other Cultural Conditions for Optimal Amylase Yield from Aspergillus oryzae-SR2. American Journal of Food Technology, 8: 54-64.
DOI: 10.3923/ajft.2013.54.64
URL: https://scialert.net/abstract/?doi=ajft.2013.54.64
DOI: 10.3923/ajft.2013.54.64
URL: https://scialert.net/abstract/?doi=ajft.2013.54.64
INTRODUCTION
Amylases are a group of hydrolases that can specifically cleave the O-glycosidic bonds in starch and are therefore employed in the starch processing industries for hydrolysis of starch into simple sugars (Alva et al., 2007; Crabb and Mitchinson, 1997). Currently, amylases are utilized in other fields, including medicinal and analytical chemistry, textile, food processing and brewing industries (Nimkar et al., 2010; Chimata et al., 2010; Anto et al., 2006).
Amylases are widely distributed in nature and can be sourced from plants, animals and microorganisms (Pandey et al., 2005; Reddy et al., 2003). Fungal and bacterial amylases have predominant applications in the industrial sector (Irfan et al., 2012). However, the ease of manipulation and recovery which subsequently leads to reduction in production cost is the major advantage associated with the use of fungi for amylase production (Irfan et al., 2012). Among fungi, many species of Aspergillus and Rhizopus have been reported to be good sources of fungal amylases (Gupta et al., 2010; Varalakshmi et al., 2009).
Aspergillus species produce a large variety of extracellular amylases which are of significant industrial importance (Pandey et al., 2000). Since Aspergillus species have been reported to be a good source of amylase enzyme, it is worthwhile to isolate its suitable strain for efficient utilization. Selection of suitable organisms plays a key role in high yield of desirable enzymes (Negi and Banerjee, 2006) and the selection of suitable production media is also very essential for both the growth of the organism and the production of the enzyme. For instance, cultural and nutritional factors have been reported to have great influence in the production of amylases by moulds (Pedersen and Nielson, 2000). Therefore, the aim of study was to isolate a good strain of Aspergillus species and determine its adequate growth media and other cultural conditions for its maximum enzyme yield.
MATERIALS AND METHODS
Isolation of Aspergillus species: Four Aspergillus species were isolated from spoilt corn and rice grains collected from home environment. The grains were carefully homogenized and 1 g of each type was dissolved in 100 mL of sterile distilled water. The diluted suspension (0.1 mL) was inoculated onto Czapek Dox agar plates supplemented with streptomycin (Varalakshmi et al., 2009). They were identified through morphological characteristics by the methods of Campbell and Stewart (1980) and as described by Diba et al. (2007) using Meat extract agar and Czapek Yeast agar. The morphological characteristics studied include the colony diameter, conidia color, colony texture, conidia heads, size and shapes, vesicles shapes and seriation. The observed macroscopic and microscopic features were compared with features of the standard species and that of other described Aspergillus species (Diba et al., 2007; Mohammadi et al., 2009). The culture was maintained at 4°C in Potato Dextrose agar slant (PDA).
Screening for amylase activity: The four isolated Aspergillus species were screened by Starch hydrolysis test (Aneja, 2002). The best isolate (with the disposition number SR2) which demonstrated the highest enzyme activity by producing the greatest zone of clearance was utilized for further study.
Inoculums preparation: Ten millimeter of sterile distilled water was carefully sprinkled onto 5 days old culture of A. oryzae-SR2 maintained on PDA agar slant. A sterile inoculating loop was used to dislodge the clustered spores and shook well to obtain a homogeneous suspension. Five millimeter (5 mL) of the spore suspension equivalent to 5x105 served as the inoculums.
Preliminary amylase production by A. oryzae-SR2: Initial 72 h-shake flask fermentation at 30°C/100 rpm (Mishra and Dadhich, 2010) was carried out with 5% inoculums in a soluble starch-mineral-salt medium containing (w/v), soluble starch (1%), yeast extract (0.5%), CaCl2 (0.03%), MgSO4.7H2O (0.2%) and FeSO4.7H2O (0.05%). At the end of the fermentation, the fungal mass was separated by centrifugation at 4500 rpm for 10 min. The clear supernatant which served as the crude enzyme was assayed.
Preparation of crude enzyme for assay: The crude enzyme was concentrated (75%) by slow addition of solid ammonium sulphate at 4°C (Gupta et al., 2010). The precipitated protein was separated by centrifugation at 1200 rpm for 30 min and thereafter dissolved with 20 mL of citrate phosphate buffer (20 mM; pH 6.0).
Amylase assay: The enzyme activity was determined by Dinitrosalicyclic Acid (DNS) method described by De Moraes et al. (1999), using soluble starch as substrate. The soluble starch was buffered with 2 mM citrate buffer (pH 6.0). The reaction mixture containing equal volumes of the enzyme and 10% substrate was incubated at 35°C for 30 min. It was terminated by the addition of 3 mL of DNS solution (3, 5-dinitrosalicyclic acid). After stopping the reaction, the tubes were placed in boiling water-bath for 5 min and thereafter cooled for absorbance at 540 nm. Blanks were prepared with inactivated enzyme. One unit of amylase was the amount of enzyme that produced one micromole of reducing sugar, measured as glucose under experimental conditions.
Determination of fermentation parameters
Carbon/nitrogen substrates: The carbon sources used were processed and unprocessed rice, corn, edible dry-flakes fermented tapioca and sugar-cane juice, all purchased from Nigeria local market. Soluble starch which served as the control carbon source was provided by the Department. Bambara meal, Soy meal, peanut meal and corn-steep- liquor, purchased and prepared locally served as the local nitrogen sources while yeast extract, peptone, egg albumin and urea purchased from Naberg-Chemical store Nigeria, were the refined sources. All the local substrates were prepared as described by George-Okafor and Mike-Anosike (2012). Each liquid substrate (100 mL) was concentrated (40%) by first stirring with a stirrer at 80 rpm for 5 min to obtain homogenous mixture. Thereafter, the mixture was subjected to air-drying using hot air drying oven at 60°C for 2-3 h prior to use. The inducing effects of these local and refined substrates (0.5-1%) were tested in two phases: (1), as sole carbon/nitrogen sources of the medium and (2) as additional supplement to the best carbon/nitrogen- sources of the basal mineral salt medium earlier described. A control flask was maintained in each test without the addition of either carbon or nitrogen source.
Determination of other parameters: The optimum pH was determined by adjusting the pH of the medium at range of 4-10 which was achieved by the addition of 0.2 M citrate phosphate with either HCl or NaOH prior to sterilization. The inoculums size was determined by varying the number of mycelia agar plugs (2-6) obtained using sterile cork borer of 2 cm in diameter. The weight of each mycelium agar was determined and thereafter the percentage (%) mass (4-12%) utilized for the assay was derived by comparing the weight of the portion of mycelia used with that of the total grown mycelia (50 agar plugs) which was taken as 100%. Fermentation time was optimized by subjecting the fermentation to various periods (1-5 day). A temperature range of 25-85°C was considered for the maximum enzyme yield.
Hydrolytic potentials of the enzyme: Starch powder from our local starchy edible food substances (Cassava, Corn, Rice, Yam, Cocoyam) were prepared as described by Ghosh and Ray (2011). The obtained starches were subjected to enzyme action using refined soluble starch as control. The enzyme activity was assayed as earlier described. Thereafter, temperature profile (30-90°C) for the conversion efficiency of the enzyme was determined.
Statistical analysis: All the data were statistically analyzed by ANOVA, using a software application; Statistical Package for Social Sciences (SPSS), version 16.0.
RESULTS AND DISCUSSION
Selection of amylase producers: The result on amylase production by starch hydrolysis test showed that the 4 fungal isolates with their disposition numbers identified as, Aspergillus awamori-SC1, Aspergillus niger-SC2, Aspergillus terreus-SR1 and Aspergillus oryzae-SR2 had potentials for amylase production (>21.15±0.11 mm; Table 1). However, Aspergillus oryzae-SR2 which demonstrated the highest zone of clearance (36.31±0.24 mm) was selected for further studies on amylase production.
Initial amylase production by Aspergillus oryzae-SR2: The organism was able to produce amylase (20.122 IU mL-1; Table 1) through Submerged Fermentation (SMF), utilizing soluble starch and yeast extract as the C/N sources of the basal medium. Good production of amylase by A. oryzae has been severally reported (Farid and Shata, 2011; Esfahanibolandbalaie et al., 2008; Rahardjo et al., 2005; Raqunathan and Swaminathan, 2005) but the reported enzyme production was mostly through solid state fermentation which has some limitations on industrial application. Hence the utilization of SMF in this study for industrial application, as most commercially important enzymes have been produced by this method (Hashemi et al., 2010).
Effect of carbon sources: The use of sugar cane juice as the sole carbon source significantly enhanced amylase production (127.9%) better than the soluble starch (100%) which served as the control (Table 2). The supplementation of other C-sources with sugar-cane (the best sole c-source) indicated dry-flakes fermented tapioca as the best C-supplement, followed by corn (Table 3).
| Table 1: | Screening potentials of the fungal isolates for amylase production |
 | |
| *After screening, the initial submerged fermentation with Aspergillus oryzae-SR2 yielded 20.122 U mL-1 amylase | |
| Table 2: | Effect of various substrates used as sole carbon for amylase production by Aspergillus oryzae-SR2 |
 | |
| *Relative enzyme yield was calculated in relation to soluble starch (100%) which served as the control | |
| Table 3: | Effect of supplementation of various carbon sources (0.5%) to the sugar cane juice medium (0.5%) for amylase production |
 | |
| *Relative enzyme yield was calculated in relation to sugar cane juice (100%) which served as the control (best sole carbon-source), +Sugar cane juice with supplementary carbon source | |
Generally, the supplementation with other C-substrates resulted in a considerable increase in enzyme yield (108.64-119.27%) except for processed rice which slightly repressed enzyme production (91.44%).
Thus, the supplementation of dry-flakes fermented tapioca with sugar-cane juice served as the c-source used for further experimental work.
The utilization of sugar-cane juice is encouraging as it is available and affordable in Nigeria. Although it is a little more expensive than its wastes (sugar cane bagasse) which has been reported to be good C-substrate for amylase production (Karamkar and Ray, 2010); its supplementation with cheap dry-flakes fermented tapioca would tremendously reduce the production cost. The use of tapioca is promising and it has been reported to be a good inducer of amylase production (Ghosh and Ray, 2011; Vijayaraghavan et al., 2011). On the other hand, the observed decrease in enzyme yield by processed rice (Table 3), might have resulted from catabolite repression generated by the processed rice.
Effect of nitrogen sources: Varying amounts of amylase yields were achieved by the tested nitrogen sources (Table 4). The refined N-sources induced higher enzyme production (>72.14%) than the local ones (50.9-70.69%). The yeast extract which was the initial N-substrate used in the basal medium still maintained the best N-source for maximum amylase production (100%).
The result obtained (Table 5) showed that the best N-source supplement (Corn-steep liquor) induced some increase in the enzyme yield (88.51%) when compared to the initial result (Table 4) but had no significant effect when compared with the control (100%). This suggests that the catabolites (local N-sources) greatly antagonized with the yeast extract, thereby deregulated nitrogen assimilation process for maximum enzyme production. So the use of yeast extract as the sole N-source is recommended. This result is in contrast to the observation by Pedersen and Nielson (2000) who reported casein hydrolysate instead of yeast extract as the best N-source for their strain of A. oryzae.
| Table 4: | Effect of various nitrogen sources on amylase yield from Aspergillus oryzae-SR2 |
 | |
| *Relative enzyme yield was calculated in relation to yeast extract (100%) which served as the control | |
| Table 5: | Effect of supplementary nitrogen sources on amylase production |
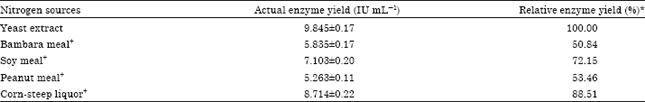 | |
| *Relative enzyme yield was calculated in relation to yeast extract (100%) which served as the control (best nitrogen-source), +Yeast extract with supplementary nitrogen source | |
However, similar results by other researchers (Irfan et al., 2012; Roohi et al., 2011; Oshoma et al., 2010; Valaparla, 2010; Narayana and Vijayalakshmi, 2008) also indicated yeast extract as good organic nitrogen source for amylase production for both fungi and bacteria. The yeast extract seemed to have stimulated the promoter genes for growth and enzyme production due to the presence of vitamin B group.
Effect of pH (4-10) on amylase production by A. oryzae-SR2 is shown in Fig. 1. Maximum enzyme production (100%) was achieved at pH 7.0. Further increase in the pH resulted in the decrease of enzyme yield (<75.8%). This result is closely related to that observed by Sivakumar et al. (2012) and Varalakshmi et al. (2009) with pH 7.0 and 7.5 as best for optimum amylase yield from Bacillus cereus and A. niger J.G.I.24. Conversely, it is higher than that from A. niger-ML-17 and A. oryzae at optimum pH of 5.0 and 6.2, respectively (Irfan et al., 2012; Esfahanibolandbalaie et al., 2008).
Temperature profile on amylase production: The amylase production by A. oryzae-SR2 was maximum at 55°C (Fig. 2), suggesting the enzyme to be thermophilic in nature; an advantage for biotechnological application. This high temperature preference for amylase production by A. oryzae-SR2 is contrary to most of the reported fungal amylase production which ranged between 25-37°C (Karamkar and Ray, 2010; Esfahanibolandbalaie et al., 2008; Peixoto-Nogueira et al., 2008). However, similar reports of high temperature optima (50 and 60°C) for amylase production by A. flavus (NTICC-08143) and A. flavus (NTICC-08150) have been recorded (Morya and Yaday, 2009) and these results resemble high temperature requirement by some Bacillus spp. for enzyme production (George-Okafor and Mike-Anosike, 2012; Sivakumar et al., 2012; George-Okafor and Odibo, 2011; Femi-Ola and Olowo, 2011).
Influence of inoculums concentration and fermentation time: Seven percent of the determined inoculums concentration at 2-day fermentation yielded the maximum amylase by the organism (Fig. 3-4).
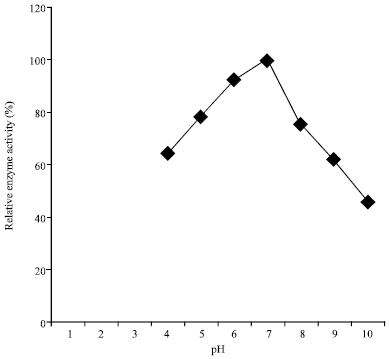 |
| Fig. 1: | pH profile on enzyme yields from A. oryzae-SR2 |
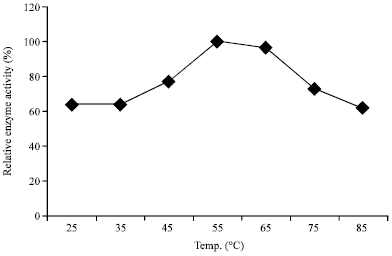 |
| Fig. 2: | Temperature influence on enzyme yield from A. oryzae-SR2 |
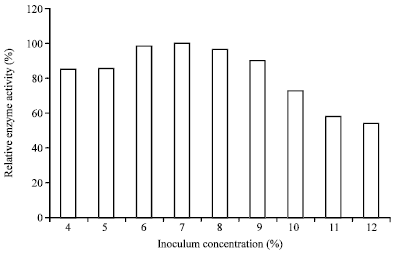 |
| Fig. 3: | Effects of inoculum concentration on enzyme production by A. oryzae-SR2 |
This result is interesting as the organism requires a shorter time for fermentation as compared to other reported fungal amylase producers which utilized up to 5 days for maximum enzyme yield (Farid and Shata, 2011). Similar fermentation period (2 days) was observed from Rhizopus oryzea for endoglucanase production in both solid and liquid states fermentation methods (Karamkar and Ray, 2010). Further increase in the incubation period yielded decrease in amylase production; indicating a peak enzyme yield during the growth phase of the organism. Similarly, a gradual decrease in enzyme yield was observed with the increase in inoculums size and this is also similar to the observation by Esfahanibolandbalaie et al. (2008) which indicated that increasing the inoculums concentration >7-8%, resulted in tremendous decrease in α-amylase production by A. oryzae in submerged fermentation. This shows that high inoculums mass must have reduced the available surface area of the medium, thereby reducing the oxygen concentration required by the organism for fermentation. The reports by other researchers also indicated the adverse effects of high load of fungal mass on enzyme production (Acharya et al., 2008; Chimata et al., 2010).
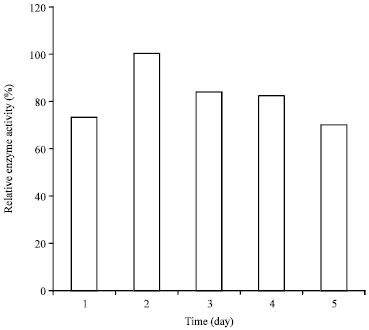 |
| Fig. 4: | Influence of fermentation time on enzyme production by A. oryzae-SR2 |
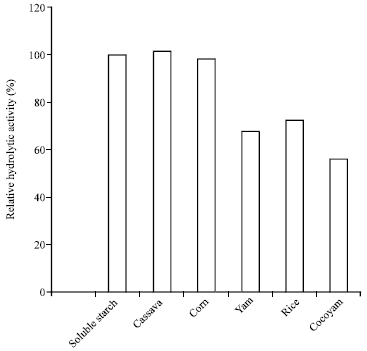 |
| Fig. 5: | Hydrolytic potentials of amylase from A. oryzae-SR2 |
Hydrolysis of starchy food substances: The enzyme was able to saccharify (>56%) all the tested starchy food substances with the cocoyam starch being the least hydrolyzed. Cassava starch was the most hydrolyzed (101.5%) and the hydrolysis was higher than that of the soluble starch (100%) even though the difference was not significant (p<0.5) (Fig. 5). The variation in the hydrolytic activity demonstrated by the enzyme could be linked to the substrate-level saturation and the nature of the starch granules as amylolytic hydrolysis of native starch granules is governed by the specific surface area and not by the mass concentration of each granule (Kim et al., 2008).
| Table 6: | Temperature profile for the conversion efficiency of the enzyme |
 | |
The hydrolytic potentials of the enzyme suggest its utilization in starch processing industry where it can be effectively utilized as starch-conversion enzyme in the production of maltodextin, modified starch, glucose syrups or anti-stalling agents in baking.
Temperature profile on starch conversion efficiency: The temperature profile on the hydrolysis of cassava starch (the best hydrolyzed substrate) showed that the maximum conversion efficiency was at 70°C, with the least at 30°C (Table 6). This directly demonstrates the enzyme stability at high temperature range of 50-90°C and thus confirms its thermo-stability property.
CONCLUSION
An increase in amylase yield achieved with the isolated A. oryzae-SR2, using low-cost substrates under submerged fermentation gave an insight for its suitability for biotechnological applications especially in Nigeria where it can replace conventional synthetic processes which are expensive. The stability and starch conversion efficiency of the enzyme at high temperature are additional advantages for its industrial applications. However, further studies on enzyme characterization and immobilization will be carried out in order to confirm its other useful properties for full industrial utilization.
ACKNOWLEDGMENTS
The authors acknowledge the assistance of Mr. S.O. Asogwa of the Department of Mathematics and Statistics, Enugu State University of Science and Technology, for the statistical analysis of the experimental data. Their appreciation also goes to the Departments for the provision of some infrastructural facilities.
REFERENCES
- Acharya, P.B., D.K. Acharya and H.A. Modi, 2008. Optimization for cellulose production by Aspergillus niger using saw dust as substrate. Afr. J. Biotechnol., 7: 4147-4152.
Direct Link - Alva, S., J. Anupama, J. Savla, Y.Y. Chiu and P. Vyshali et al., 2007. Production and characterization of fungal amylase enzyme isolated from Aspergillus sp. JGI 12 in solid state culture. Afr. J. Biotechnol., 6: 576-581.
Direct Link - Anto, H., U.B. Trivedi and K.C. Patel, 2006. Glucoamylase production by solid-state fermentation using rice flake manufacturing waste products as substrate. Bioresour. Technol., 97: 1161-1166.
CrossRefPubMedDirect Link - Crabb, W.D. and C. Mitchinson, 1997. Enzymes involved in the processing of starch tosugars. Trends Biotechnol., 15: 349-352.
CrossRefDirect Link - De Moraes, L.M.P., S.A. Fillo and C.J. Ulhoa, 1999. Purification and some properties of an α-amylase glucoamylase fusion protein from Saccharomyces cerevisiae. World J. Microbiol. Biotechnol., 15: 561-564.
CrossRef - Diba, K., P. Kordbacheh, S.H. Mirhendi, S. Rezale and M. Mahmoudi, 2007. Identification of Aspergillus species using morphological characteristics. Pak. J. Med. Sci., 23: 867-872.
Direct Link - Esfahanibolandbalaie, Z., K. Rostami and S.S. Mirdamadi, 2008. Some studies of α-amylase production using Aspergillus oryzae. Pak. J. Biol. Sci., 11: 2553-2559.
CrossRefPubMedDirect Link - George-Okafor, U.O. and E.E. Mike-Anosike, 2012. Screening and optimal protease production by Bacillus sp. SW-2 using low cost substrate medium. Res. J. Microbiol., 7: 327-336.
Direct Link - George-Okafor, U.O. and F.J.C. Odibo, 2011. Purification and Some Properties of Thermo-stable Alkaline Serine Protease from Thermophilic Bacillus sp. Gs-3. J. Biol. Sci., 11: 299-306.
CrossRef - Ghosh, B. and R.R. Ray, 2011. Extra-cellular isoamylase production by Rhizopus oryzae in solid-state fermentation of agro wastes. Brazil. Arch. Biol. Technol., 54: 867-876.
CrossRefDirect Link - Gupta, A., N. Gautam and D.R. Modi, 2010. Optimization of α-amylase production from free and immobilized cells of Aspergillus niger. J. Biotechnol. Pharma. Res., 1: 1-8.
Direct Link - Kim, J.C., B.W. Kong, M.J. Kim and S.H. Lee, 2008. Amylolytic hydrolysis of native starch granules affected by granule surface area. J. Food Sci., 73: C621-C624.
CrossRef - Roohi, K.M., I.Z. Ahmad and J.M. Arif, 2011. Production of coldactive extracellular α-amylase by newly isolated Microbacterium foliorum GA2 from gangotri glacier, Western Himalaya, India. Asian. J. Biotechnol., 3: 449-459.
CrossRef - Mishra, B.K. and S.K. Dadhich, 2010. Production of amylase and xylanase enzymes from soil fungi of Rajasthan. J. Adv. Dev. Res., 1: 21-23.
Direct Link - Mohammadi, A.H., Z. Banihashemi and M. Haghdel, 2009. Identification and prevalence of Aspergillus species in soils of Fars and Kerman provinces of Iran and evaluation of their Aflatoxin production. Rostaniha, 10: 9-30.
Direct Link - Narayana, K.J.P. and M. Vijayalakshmi, 2008. Production of extracellular α-amylase by Streptomyces albidoflavus. Asian J. Biochem., 3: 194-197.
CrossRefDirect Link - Oshoma, C.E., E.E. Imarhiagbe, M.J. Ikenebomeh and H.E. Eigbaredon, 2010. Nitrogen supplements effect on amylase production by Aspergillus niger using cassava whey medium. Afri. J. Biotechnol., 9: 682-686.
Direct Link - Pandey, A., P. Nigam, C.R. Soccol, V.T. Soccol, D. Singh and R. Mohan, 2000. Advances in microbial amylases. Biotechnol. Applied Biochem., 31: 135-152.
PubMedDirect Link - Pedersen, H. and J. Nielsen, 2000. The influence of nitrogen sources on alpha amylase productivity of Aspergillus oryzae in continuous culture. Applied Microbiol. Biotechnol., 53: 278-281.
CrossRefDirect Link - Peixoto-Nogueira, S.C., V.C. Sandrim, H.S. Guimaraes, J.A. Jorge, H.F. Terenzi and M.L.T.M. Polizeli, 2008. Evidence of thermostable amylolytic activity from Rhizopus microsporus var. rhizopodiformis using wheat bran and corncob as alternative carbon source. Bioprocess Biosys. Eng., 31: 329-334.
CrossRef - Rahardjo, Y.S.P., F.J. Weber, S. Haemers, J. Tramper and A. Rinzema, 2005. Aerial mycelia of Aspergillus oryzae accelerate α- amylase production in a model solid state fermentation system. Enzyme Microb. Technol., 36: 900-902.
CrossRef - Raqunathan, R. and K. Swaminathan, 2005. Growth and amylase production by Aspergillus oryzae during solid state fermentation using banana wastes as substrates. J. Environ. Biol., 26: 653-656.
PubMed - Varalakshmi, K.N., B.S. Kumudini, B.N. Nandini, J. Solomon, R. Suhas, B. Mahesh and A.P. Kavitha, 2009. Production and characterization of α-amylase from Aspergillus niger JGI 24 isolated in Bangalore. Pol. J. Microbiol., 58: 29-36.
PubMedDirect Link - Femi-Ola, T.O. and B.M. Olowe, 2011. Characterization of α amylase from Bacillus subtilis BS5 isolated from Amitermes evuncifer silvestri. Res. J. Microbiol., 6: 140-146.
CrossRefDirect Link