
Jing Zhou
College of Life Sciences, China Jiliang University, Hangzhou, Zhejiang, 310018, China
Wen-Wei Chen
College of Life Sciences, China Jiliang University, Hangzhou, Zhejiang, 310018, China
Zheng-Bao Jia
College of Life Sciences, China Jiliang University, Hangzhou, Zhejiang, 310018, China
Guang-Rong Huang
College of Life Sciences, China Jiliang University, Hangzhou, Zhejiang, 310018, China
Yao Hong
College of Life Sciences, China Jiliang University, Hangzhou, Zhejiang, 310018, China
Jun-Jie Tao
College of Life Sciences, China Jiliang University, Hangzhou, Zhejiang, 310018, China
Xiao-Bo Luo
College of Life Sciences, China Jiliang University, Hangzhou, Zhejiang, 310018, China
American Journal of Food Technology
Year: 2012 | Volume: 7 | Issue: 10 | Page No.: 596-608







ABSTRACT
Lipase was one of the most important industrial enzymes and has diverse applications in food industry, detergent industry, fats and oil hydrolysis, peptide synthesis and pharmaceutical industries. The aim of this study was focused on isolation of a lipase producing strain from waste cooking oily soil and characterization of the lipase. The strain, Aspergillus oryzae CJLU-31 was isolated from the waste cooking oily soil and identified by ITS rDNA analysis. The lipase from Aspergillus oryzae CJLU-31 was purified by ammonium sulfate precipitation and two-step chromatography. The results showed that the purified lipase had a specific activity of 11552.2 U mg-1, 23.9-fold purification factor and molecular mass of 27 kDa by Sodium Dodecyl Sulfate-polyacrylamide Gel Electrophoresis (SDS-PAGE). The purified lipase showed higher activity when the waste cooking oil was used as the substrate. It also showed maximum activity and stability at pH 4.0 and 40°C. The lipase retained high activity over ranges of pH (2-5) and temperature (30-50°C). It was enhanced by low concentration metal ions, such as K+, Li+, Mg2+, Zn2+, Mn2+ and Ca2+, while inhibited by Fe2+, Fe3+ and Cu2+. It showed high stability in the presence of lower polarity organic solvents and activated in 0.1% polyvinyl alcohol-124. The Km and Vmax of the lipase acting on olive oil were 0.11 and 0.41 mM min-1, respectively. Heating and cooling refold-treatment activated the lipase. These features provided the lipase from Aspergillus oryzae CJLU-31 as a potential application for lipase-catalyzed synthesis of fatty acid methyl esters (biodiesel) using waste cooking oil.
PDF Abstract XML References Citation
Received: April 27, 2012;
Accepted: July 16, 2012;
Published: October 09, 2012
How to cite this article
Jing Zhou, Wen-Wei Chen, Zheng-Bao Jia, Guang-Rong Huang, Yao Hong, Jun-Jie Tao and Xiao-Bo Luo, 2012. Purification and Characterization of Lipase Produced by Aspergillus oryzae CJLU-31 Isolated from Waste Cooking Oily Soil. American Journal of Food Technology, 7: 596-608.
DOI: 10.3923/ajft.2012.596.608
URL: https://scialert.net/abstract/?doi=ajft.2012.596.608
DOI: 10.3923/ajft.2012.596.608
URL: https://scialert.net/abstract/?doi=ajft.2012.596.608
INTRODUCTION
Waste Cooking Oil (WCO) originated from food restaurants and household disposals, was creating serious problems of food safety and environmental control in China. In order to obtain higher economic benefit, WCO was recovered to produce edible oil by some undesirable businessmen, though this behavior was prohibited by law in China. Production of biodiesel with WCO as feedstock not only could reduce these problems, but, more importantly, would decrease the cost of biodiesel (Chen et al., 2005; Chhetri et al., 2008; Wan Omar et al., 2009).
Lipase of triacylglycerol hydrolase (EC 3.1.1.3) was one of the carboxyl esterases, which catalyzed the hydrolysis of triacylglycerol synthesis to form esters and the long chain fatty acids at the oil-water interface and transesterification of acylglycerols (Zaks and Klibanov, 1985; Susumu et al., 1979). Lipases constitute one of the most important groups of industrial enzymes due to their unique ability of activities in both aqueous and non-aqueous solvent systems (Gaur et al., 2008). Because of these advantages, they became the focus of bio-refinery. In recent years, many scholars had carried out a lot of studies aiming at lipase-catalyzed synthesis of biodiesel (De los Rios et al., 2011; Halim and Kamaruddin, 2008; Yan et al., 2011; Al-Zuhair et al., 2009). Isolation of new lipase with potential industrial applications from novel sources stimulated many researchers interest (Islam et al., 2008).
Aspergillus oryzae was one of the extensively investigated lipase-production microorganisms and had been applied widely in lipase production industry or other related industry (Contesini et al., 2010; Lee et al., 2006; Elbashiti et al., 2010; El-Atta et al., 2011). First, they are on the Generally Recognized As Safe (GRAS) list of the Food and Drug Administration (FDA) in the United States. Then they are recognized as the best lipase producers since they produce extracellular lipases, facilitating the extraction from fermentation media (Carvalho et al., 2005). Many studies had optimized the culture compositions, nutritional requirements for production of Aspergillus spp. lipase and purified the homogeneity using a variety of methods involving ammonium sulfate precipitation, ion exchange chromatography followed by gel filtration (Pei et al., 2009; Basheer et al., 2011; Ventura et al., 2012). In addition, lipases from various sources had been characterized in terms of their activity and stability profiles with respect to pH, temperature, metal ions, detergents and organic solvents (Yadav et al., 2011; Zheng et al., 2011; Rivera-Perez et al., 2011) as well as their molecular weights (Jiang et al., 2012; Supakdamrongkul et al., 2010).
In the present study, therefore a strain of Aspergillus oryzae, named CJLU-31, was isolated from WCO waste soil and identified by ITS rDNA (first internal transcribed spacer of rDNA) sequence analysis. This strain was to have high acidic extracellular lipase production ability. Also, the production, purification and characterization of the lipase from Aspergillus oryzae CJLU-31 was investigated.
MATERIALS AND METHODS
Isolation and identification of lipolytic bacterial strain: The oily waste soil sample was collected in September 2010, from Xiasha area, Hangzhou city, Zhejiang province, China. The soil sample (5.0 g) was suspended in 45.0 mL of sterilized normal saline for 24 h and serial dilutions were made. Aliquots (200 μL) of appropriate dilutions were surface plated on the selective plate containing (NH4)2SO4: 2.0 g L-1, K2HPO4: 1.0 g L-1, KCl: 0.5 g L-1, MgSO4.7H2O: 0.5 g L-1, FeSO4: 0.01 g L-1, agar: 20.0 g L-1, bromcresol purple indicator: 0.004 g L-1 and olive oil emulsion (30.0 g L-1 oil emulsified with 90 g L-1 (v/v) polyvinyl alcohol-124): 120.0 g L-1. Plates were then incubated at 30°C and periodically examined for 5-7 days. The size of the yellow color ring was used to evaluate lipase production. The micromorphology of the isolate was studied by viewing spore morphology and microflora in Czapek’s medium plate.
Confirmation of the assigned taxon was carried out by ITS gene sequence analysis. PCR amplification of this gene was carried out using primers ITS1 (5-TCCGTAGGTGAACCTGCGG-3) and ITS4 (5-TCCTCCGCTTATTGATATGC-3). Partial ITS gene sequence thus obtained was submitted to GenBank database at NCBI (GenBank accession No. JN874631.1, Zhou, 2011). Sequence comparisons were performed with the BLAST program (NCBI, USA).
Lipase production: The fermentation condition was optimized through single factor test and orthogonal test. Aspergillus oryzae CJLU-31 was grown in a basal medium containing (NH4)2SO4: 1.0 g L-1, K2HPO4: 1.0 g L-1, MgSO4.7H2O: 1.0 g L-1, beef extract: 30.0 g L-1, polyvinyl alcohol-124: 0.025% and olive oil: 0.125 g L-1 at pH 7.0. Culture conditions were: 100 mL of medium in 250 mL Erlenmeyer flask, strain age 24 h inoculum size 6%, rotation at 150 rpm at 32°C. Biomass in the broth was removed by centrifugation and the supernatant was subjected to lipase activity determination.
Lipase assay: Lipase activity was measured by copper soap calorimetric method (Kwon and Rhee, 1986; Jermsuntiea et al., 2011), using an n-hexane-olive oil micro emulsions with slight modifications. Simply, the reaction mixture (5.80 mL) consisted 0.50 mL olive oil, 4.50 mL n-hexane and 20 mM phosphate buffer 0.8 mL (emulsified with 5% (w/v) gum acacia in buffer (pH 4.0) as substrate. The olive oil substrate incubated for 5 min at 40°C. Then, 0.2 mL of enzyme was added and the reaction was allowed to proceed for 15 min. The reaction was stopped by the addition of 1 mL of 6 M HCl. Then, 0.40 mL supernatant fluid was absorbed to the centrifuge tube, with 4.6 mL toluene and 1.0 mL of 5% (w/v) cupric acetate (pH adjusted to 6.0 with pyridine). The two phases were vortexed for 60 sec and the absorbance of the toluene phase was read at 715 nm using a Apada UV-1200 spectrophotometer. The toluene phase containing the Free Fatty Acids (FFAs) was separated when 1.0 mL of copper reagent was added. Standard curve with oleic acid was used for determination of the free fatty acids. One activity unit of lipase was defined as the amount of enzyme that released 1 μmol of fatty acid per min under assay conditions.
Protein assay: The protein content was determined by the method of Bradford (1976) using bovine serum albumin as the standard.
Purification of lipase: The purification procedure was referred to Shu method (Shu et al., 2006). After 48 h of culture, the cells were separated from the fermented medium by centrifugation at 4°C and 4000 rpm. Ammonium sulphate was added to the filtrate to give final concentration of 60-80% (w/v) saturation at 4°C. The precipitate was collected by centrifugation at 4000 rpm for 4 min. The precipitate was then dissolved in 20 mL of double distilled water and dialyzed for 24 h at 4°C against double distilled water. Then, the crude enzyme solution was loaded onto the DEAE-Separose Fast Flow column (2.6x10 cm) previously equilibrated with 0.02 M phosphate buffer (pH 7.8). Elution of lipase was performed by a linear gradient of NaCl (0-1.0 mol L-1 in equilibrating buffer) at 30 mL h-1. The fractions of 5.0 mL each were collected and assayed for protein and lipase activity. The fractions showing lipase activity were dialyzed against distilled water. The fractions were loaded on Sephadex G-100 column (2.6x80 cm) and followed by elution using 0.02 M phosphate buffer (pH 7.8) at 30 mL h-1. The fractions of 2.5 mL each were collected and assayed for protein and lipase activity.
Determination of molecular weight: Sodium dedocyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was done on a 10% separating gel and 4% stacking gel (0.75 mm, 10x10.5 cm mini gel) under the conditions developed by Laemmli (1970). The protein marker ranging from 14.4-116.0 kDa was used as a standard marker for determination of molecular weight.
Effect of pH and temperature on activity and stability: The effects of pH and temperature on purified lipase were investigated by using olive oil as the substrate. The optimal pH of the purified enzyme was determined at room temperature over a pH range of 2-9-20 mM acetate buffer (pH 2-5), phosphate buffer (pH 6-8) or tris-HCl buffer (pH 8-9). The residual lipase activity was measured. The pH stability was studied by incubating the lipase at each desired pH for 30 min at 40°C.
The optimum temperature was determined by measuring the lipase activity at 30-65°C in 20 mM acetate buffer, pH 4.0. The thermal stability was studied by incubating the lipase mixture at each desired temperature for 30 min, refolding at 4°C.
Effect of metal ions, detergents and organic solvents: Several different molar concentrations of LiCl, KCl, FeSO4, FeCl3, CuSO4, MnCl2, ZnCl2, MgSO4 and CaCl2 were added to 20 mM pH 4.0 acetate buffer enzyme solution at room temperature for 30 min. Similarly, different detergents, such as tween 40, tween 80, SDS, gum acacia and polyvinyl alcohol-124, organic solvents including methanol, ethanol, ether, acetone, propanediol, glycerol and n-butanol, were studied.
Determination of kinetic parameters: The enzymatic kinetic parameters were determined according to Shangguan method (Shangguan et al., 2011). The olive oil emulsion substrate at different concentrations (1-10 mM), temperature of 40°C and pH 4.0 were employed to determine the kinetic parameters. The kinetic parameters were estimated from the Lineweaver-Burk equation plot (1):
| (1) |
where, [S] was the substrate concentration (mM), V was the initial reaction rate of the enzyme (mM min-1), Km was Michaelis-Menten constant and Vmax was maximum reaction rate.
RESULTS AND DISCUSSION
Strains isolation, identification and production of lipase from Aspergillus oryzae CJLU-31: As the result of screening, 31 colonies formed a clear yellow color ring on our plate media, which showed lipase activity.
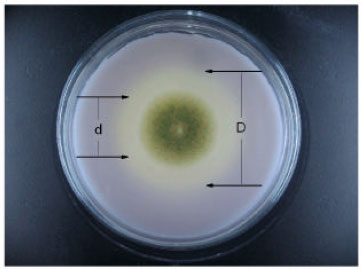 |
| Fig. 1: | Colonial morphology of CJLU-31, D: Diameter of yellow color ring, d: Diameter of colony |
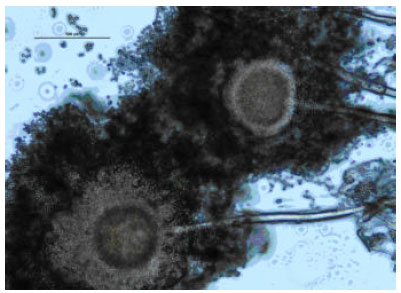 |
| Fig. 2: | Spore morphology of CJLU-31 |
 |
| Fig. 3: | Phylogenetic tree of the lipase genes, it shows the relationship of lipase producing strain Aspergillus oryzae CJLU-31 to other Aspergillus spp. with MAGE 4.0 using the neighbor-joining method, values shown in the parenthesis are accession number |
A fungus (isolate CJLU-31, Fig. 1) exhibited a highest diameter ratio between yellow color ring and colony and it showed a maximum lipase activity (16.0 U mL-1) after fermentation. Therefore, the organism produced an extracellular lipase, which was selected for the production of lipase. The spore morphology showed that the individual smoother spores were black or brown (fading to brown with age), conidiophores were longer average and the sterigmata was usually in 1 series (Fig. 2). The ITS1 sequencing data indicated that the isolate was Aspergillus (Fig. 3). The nucleotide sequence reported here had been assigned an accession number JN874631.1 from NCBI gene bank database. The BLAST result showed that the isolated strain was 100% similar to those of nearest strain Aspergillus oryzae strain 1 (accession number HQ 380782.1), named Aspergillus oryzae CJLU-31.
An appropriate incubation time for Aspergillus oryzae CJLU-31 had to be determined for the purpose of maximum lipase production as it was dependent on the dry cell weight. From the growth time profile as shown in Fig. 4, the highest activity of lipase (460.7 U mg-1) was observed in the beginning of the stationary phase. Thus, the best time period for harvesting lipase with the highest activity was optimized at 48 h, which was faster than producing lipase from Aspergillus carneus (Kaushik et al., 2006) or Aspergillus wentii (Chander et al., 1980).
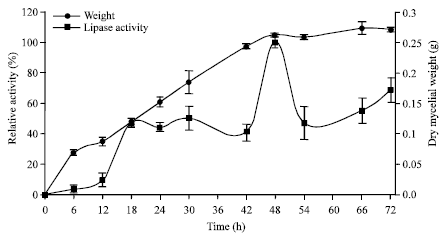 |
| Fig. 4: | Effects of incubation time on lipase activity and cell growth profile, relative activity was calculated by activity at 48 h as 100%, batches were harvested at 6 h intervals to monitor dry mycelial weight and lipase activity, the average of relative value (n = 3) and SD is represented by vertical bar |
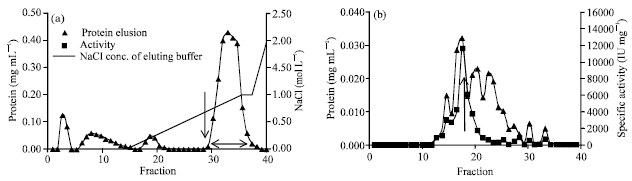 |
| Fig. 5(a-b): | Purification of lipase from Aspergillus oryzae CJLU-31, (a) Chromatography on DEAE-Separose Fast Flow and (b) Chromatography on Sephadex G-100, In each step of chromatography, elution of proteins was followed by measuring absorbance at 595 nm and the activity was followed by copper soap colorimetric method using olive as a substrate, The downward arrows indicate the initiation of elution of lipase, The transverse left-right and upward arrows indicate the fractions collected for further studies |
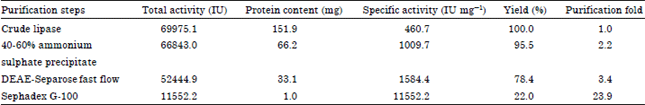
Purification of lipase: In the present study, homogeneous lipase was purified using ammonium sulphate precipitation followed by anion exchange and gel filtration chromatography. By this purification procedure a final yield of 22.0% 23.9-fold purified lipase was achieved. The specific activity of the lipase increased to 11552.2 IU mg-1 (Table 1).
The patterns of DEAE sepharose fast flow anion exchange chromatography and Sephadex G-100 gel chromatographies were shown in Fig. 5. The molecular weight was determined by the presence of a single band corresponding to an apparent molecular mass of 27 kDa on SDS-PAGE gels (Fig. 6). The molecular mass of the purified lipase in our study was similar as those reported elsewhere, using P. expansum PED-03 (Dai and Xia, 2006; Lianghua et al., 2007). In the first two process of purification, the enzyme activity was not completely separable from the pigments which were reported by Demir and Tukel (2010).
| Table 1: | Summary of purification procedure of lipase from Aspergillus oryzae CJLU-31 |
 | |
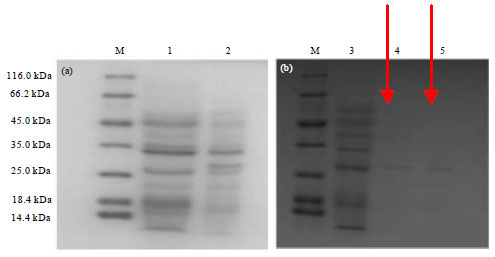 |
| Fig. 6(a-b): | SDS-PAGE of purified lipase, the purified protein was electrophoresed on 12% tris-glycine SDS-PAGE, the markers are β-galactosidase (116.0 kDa), bovine serum albumin (66.2 kDa), ovalbumin (45.0 kDa), lactase dehydrogenase (35.0 kDa), Rease Bsp98I (25.0 kDa), β-lactoglobulin (18.4 kDa) and lysozyme (14.0 kDa), M: Marker, lane 1: Crude lipase, lane 2: 40-60% ammonium sulphate precipitation, lane 3: Purified fraction after DEAE-Separose Fast Flow chromatography, lane 4 and lane 5: Purified enzyme after Sephadex G-100 gel chromatography, The downward arrows indicate the purified lipase for further studies |
The lower lipase activity was obtained in these steps, which might be related with one strain of microorganism screening different lipases. Fernandez-Lorente et al. (2005) purified different lipases from an A. niger strain by using a highly selective adsorption on hydrophobic supports.
Effects of pH and temperature: The optimal pH of purified lipase was about 4.0 (Fig. 7). The lipase activity was not much affected between pH 2-5 where it showed around 80% of residual activity. The lipase was stable in pH range of 2-5 even after incubation for 30 min (Fig. 7). Moreover, the lipase had an optimal temperature of 40°C and was also active in temperature range of 30-50°C (Fig. 8).
Interestingly, a heating and cooling cycle enhanced the activity of the lipases (Fig. 9). The lipase exhibited increased activity at 40°C after 30 min, at 50°C after 120 min and retained 100% activity after 3 h at 30°C, which was different property amongst the lipases isolated from many other fungal species (Heerden et al., 2002; Namboodiri and Chattopadhyaya, 2000). The reason was reported by Zhu et al. (2001), which showed the protein to be trapped in an intermediate, higher energy state, contribute significantly to the stability, structure and activity.
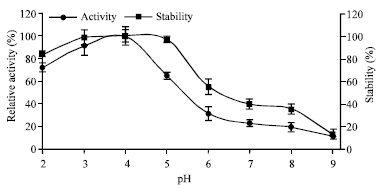 |
| Fig. 7: | Effects of pH on activity and stability of lipase, relative activity was calculated by activity at pH 4.0 without treatment as 100%, data are means of three determinations and standard errors are reported, the average of relative value (n = 3) and SD is represented by vertical bar |
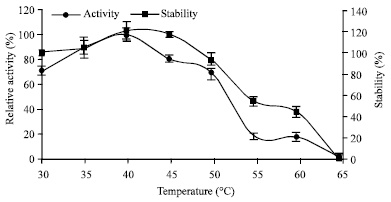 |
| Fig. 8: | Effects of temperature on activity and stability of lipase, relative activity was calculated by activity at 40°C without treatment as 100%, data are means of three determinations and standard errors are reported, the average of relative value (n = 3) and SD is represented by vertical bar |
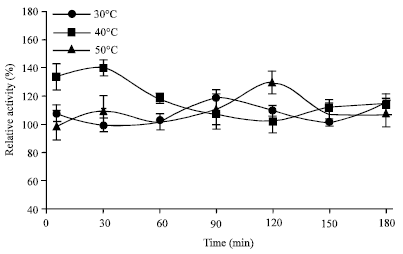 |
| Fig. 9: | Effects of heating (for 30 min) and cooling refold-treatment on activity of lipase, the relative activity at 30, 40 and 50°C for 30, 60, 90, 120, 150 and 180 min were monitored throughout the experiment and were calculated by taking the activity of pH 4.0 and 40°C without treatment as 100%, Data are means of three determinations and standard errors are reported, the average of relative value (n = 3) and SD is represented by vertical bar |
| Table 2: | Effects of detergents on lipase from Aspergillus oryzae CJLU-31 |
 | |
| Experiments were performed in triplicate and activity without detergents was set as 100% | |
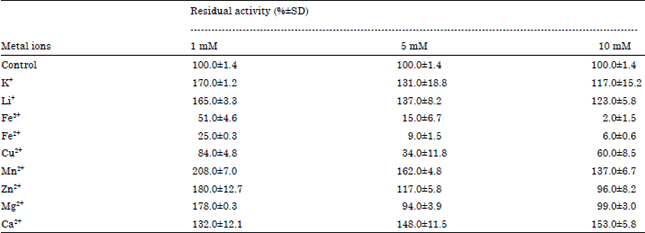
| Table 3: | Effects of metal ions on lipase from Aspergillus oryzae CJLU-31 |
 | |
| Experiments were performed in triplicate and activity without metal ions was set as 100% | |
Effects of detergents and metal ions: Table 2 depicted the effects of detergents on the purified lipase activities. The presence of Polyvinyl alcohol-124 enhanced the lipase activity. The polyvinyl alcohol-124 (0.1%, v/v) was found to increase the lipase activity to 128%. However, the lipase activity was inhibited below 50% by tween-40 and tween-80, which were classified to same kind of nonionic surfactant. Inhibition by SDS suggested that the anionic surfactant intensively decreased the activity of lipases, which was also reported by Ebrahimpour et al. (2011).
Metal ions at different concentrations were used to evaluate their effects on the lipase activity (Table 3). The monovalent metal ion such as K+ and Li+ were observed as promoters of the lipase. Meanwhile, lower concentration of K+ and Li+ were more effective to enhance the lipase activity. Some of divalent metal ions, such as Fe2+ and Cu2+, reduced the lipase activity more than 40%, but the other divalent metal ions increased significantly the lipase activity at 1 mM Mn2+ (208%), Zn2+ (180%) and Mg2+ (178%), respectively. Meanwhile, when treated with Ca2+ at increased concentrations, there was obvious enhancement of the lipase activity. Dheeman et al. (2011) reported a significant enzyme activity enhancement of 17%, when a purified microbial lipase PEL was treated with Ca2+.
Effect of organic solvents: Lipase hydrolysis reaction must occur at a water-oil interface. In order to investigate the effects of organic solvents on lipase activity, the lipase solution was incubated with organic solvents (10-30%, v/v) at room temperature, 150 rpm for 30 min and the residual activity of the lipase was shown in Table 4.
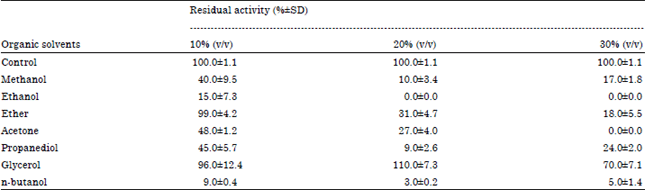
| Table 4: | Effects of organic solvents on lipase from Aspergillus oryzae CJLU-31 |
 | |
| Experiments were performed in triplicate and activity without organic solvents was set as 100% | |
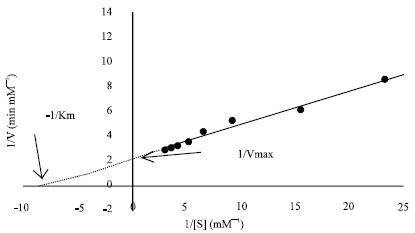 |
| Fig. 10: | Lineweaver-Burk plot of purified lipase from Aspergillus oryzae CJLU-31 using olive oil as a substrate, data are means of three determinations |
The lipase activity was inhibited with the whereas, ethanol, n-butanol strongly inhibited the lipase activity. It was believed that water-miscible organic solvents strip water from the enzymes, leading to the unfolding of the molecule with exposure of the inner hydrophobic residues and that this denaturation occurred at a much faster rate than in a pure aqueous system (Azevedo et al., 2001). In addition, lipase retained 40% of activity after incubation at 10% (v/v) methanol, which exhibited the tolerance of methanol. 20% (v/v) glycerol in the solvent, as the product of the lipase hydrolysis reaction, enhanced its activity. These two points indicated that lipase of Aspergillus oryzae CJLU-31 may be used in industrial application as catalysis.
Kinetic parameters of the lipase: Lineweaver-Burk plots in this study showed that apparent Km and Vmax values of the purified acidic lipase were 0.11 and 0.41 mM min-1, respectively (Fig. 10). The lower value of Km represents higher affinity between enzymes and substrates, while Vmax represents the higher catalytic efficiency of lipase (Sharma et al., 2001).
CONCLUSION
The strain, Aspergillus oryzae CJLU-31 was isolated from the oily (WCO) waste soil, then identified using ITS rDNA sequencing and phylogenetic analysis. The lipase showed a good activity of the waste cooking oil as the substrate and exhibited an enhanced activity after heating and cooling refold-treatment at appropriate temperature. The lipase from Aspergillus oryzae CJLU-31 might be a potential application for using waste cooking oil to produce biodiesel.
ACKNOWLEDGMENT
This study was supported by grants from the Natural Science Foundation of China, Zhejiang province (Grant No. Y307589).
REFERENCES
- Al-Zuhair, S., A. Dowaidar and H. Kamal, 2009. Dynamic modeling of biodiesel production from simulated waste cooking oil using immobilized lipase. Biochem. Eng. J., 44: 256-262.
CrossRef - Azevedo, A.M., D.M.F. Prazeres, J.M.S. Cabral and L.P. Fonseca, 2001. Stability of free and immobilised peroxidase in aqueous-organic solvents mixtures. J. Mol. Catal. B: Enzym., 15: 147-153.
CrossRef - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Carvalho, P.D.O., F.J. Contesini, R. Bizaco and G.A. Macedo, 2005. Kinetic properties and enantioselectivity of the lipases produced by four Aspergillus species. Food Biotechnol., 19: 183-192.
CrossRef - Chander, H., V.K. Batish, S.S. Sannabhadti and R.A. Srinivasan, 1980. Factors affecting liapse production in Aspergillus wentii. J. Food Sci., 45: 598-600.
CrossRef - Chhetri, A.B., K.C. Watts and M.R. Islam, 2008. Waste cooking oil as an alternate feedstock for biodiesel production. Energies, 1: 3-18.
CrossRef - Contesini, F.J., D.B. Lopes, G.A. Macedo, M.D.G. Nascimento and P.D.O. Carvalho, 2010. Aspergillus sp. lipase: Potential biocatalyst for industrial use. J. Mol. Catal. B: Enzym., 67: 163-171.
CrossRef - Dai, D. and L. Xia, 2006. Effect of lipase immobilization on resolution of (R, S)-2-octanol in nonaqueous media using modified ultrastable-Y molecular sieve as support. Appl. Biochem. Biotechnol., 134: 39-50.
CrossRef - De los Rios, A.P., F.J.H. Fernandez, D. Gomez, M. Rubio and G. Villora, 2011. Biocatalytic transesterification of sunflower and waste cooking oils in ionic liquid media. Process Biochem., 46: 1475-1480.
CrossRef - Demir, B.S. and S.S. Tukel, 2010. Purification and characterization of lipase from Spirulina platensis. J. Mol. Catal. B: Enzym., 64: 123-128.
CrossRef - Dheeman, D.S., G.T.M. Henehan and J.M. Frias, 2011. Purification and properties of Amycolatopsis mediterranei DSM 43304 lipase and its potential in flavour ester synthesis. Bioresour. Technol., 102: 3373-3379.
CrossRef - Ebrahimpour, A., R.N.Z.R.A. Rahman, M. Basri and A.B. Salleh, 2011. High level expression and characterization of a novel thermostable, organic solvent tolerant, 1,3-regioselective lipase from Geobacillus sp. strain ARM. Bioresour. Technol., 102: 6972-6981.
CrossRefDirect Link - El-Atta, H.A., I.M. Aref and S.A. Khalil, 2011. Increased gum arabic production after infestation of Acacia senegal with Aspergillus flavus and Pseudomonas pseudoalcaligenes Transmitted by Agrilus nubeculosus. Biotechnology, 10: 159-166.
CrossRefDirect Link - Elbashiti, T., A. Fayyad and A. Elkichaoui, 2010. Isolation and identification of Aspergillus oryzae and the production of soy sauce with new aroma. Pak. J. Nutr., 9: 1171-1175.
CrossRefDirect Link - Gaur, R., A. Gupta and S.K. Khare, 2008. Purification and characterization of lipase from solvent tolerant Pseudomonas aeruginosa PseA. Process Biochem., 43: 1040-1046.
CrossRefDirect Link - Halim, S.F.A. and A.H. Kamaruddin, 2008. Catalytic studies of lipase on FAME production from waste cooking palm oil in a tert-butanol system. Process Biochem., 43: 1436-1439.
CrossRef - Heerden, E.V., D. Litthauer and R. Verger, 2002. Biochemical characterisation and kinetic properties of a purified lipase from Aspergillus niger in bulk phase and monomolecular films. Enzyme Microb. Technol., 30: 902-909.
CrossRef - Islam, M.A., N. Absar and A.S. Bhuiyan, 2008. Isolation, purification and characterization of lipase from grey mullet (Liza parsia Hamilton, 1822). Asian J. Biochem., 3: 243-255.
CrossRefDirect Link - Jermsuntiea, W., T. Aki, R. Toyoura, K. Iwashita, S. Kawamoto and K. Ono, 2011. Purification and characterization of intracellular lipase from the polyunsaturated fatty acid-producing fungus Mortierella alliacea. New Biotechnol., 28: 158-169.
CrossRef - Jiang, J., B. Shi, D.Q. Zhu, Q.X. Cai and Y.R. Chen et al., 2012. Characterization of a novel bacteriocin produced by Lactobacillus sakei LSJ618 isolated from traditional Chinese fermented radish. Food Control, 23: 338-344.
CrossRefDirect Link - Kaushik, R., S. Saran, J. Isar and R.K. Saxena, 2006. Statistical optimization of medium components and growth conditions by response surface methodology to enhance lipase production by Aspergillus carneus. J. Mol. Catal. B: Enzym., 40: 121-126.
CrossRefDirect Link - Kwon, D.Y. and J.S. Rhee, 1986. A simple and rapid colorimetric method for determination of free fatty acids for lipase assay. J. Am. Oil Chem. Soc., 63: 89-92.
CrossRefDirect Link - Lee, K., S.K. Lee and B.D. Lee, 2006. Aspergillus oryzae as probiotic in poultry: A review. Int. J. Poult. Sci., 5: 1-3.
CrossRefDirect Link - Lianghua, T., X. Liming, S. Min and G. Huaying, 2007. Purification and application of a lipase from Penicillium expansum PED-03. Appl. Biochem. Biotechnol., 142: 194-199.
CrossRef - Namboodiri, V.M. and R. Chattopadhyaya, 2000. Purification and biochemical characterization of a novel thermostable lipase from Aspergillus niger. Lipids, 35: 495-502.
PubMed - Pei, Y., J. Wang, K. Wu, X. Xuan and X. Lu, 2009. Ionic liquid-based aqueous two-phase extraction of selected proteins. Sep. Purif. Technol., 64: 288-295.
CrossRef - Rivera-Perez, C., M.A.N. del Toro and F. Garcia-Carreno, 2011. Purification and characterization of an intracellular lipase from pleopods of whiteleg shrimp (Litopenaeus vannamei). Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol., 158: 99-105.
CrossRef - Shu, S.C., C.J. Xu and G.C. Lin, 2006. Purification and partial characterization of a lipase from Antrodia cinnamomea. Process Biochem., 41: 734-738.
CrossRef - Supakdamrongkul, P., A. Bhumiratana and C. Wiwat, 2010. Characterization of an extracellular lipase from the biocontrol fungus, Nomuraea rileyi MJ and its toxicity toward Spodoptera litura. J. Invertebr. Pathol., 105: 228-235.
CrossRef - Susumu, O., I. Mieko and T. Yoshio, 1979. Synthesis of various kinds of esters by four microbial lipases. Biochim. Biophys. Acta, 575: 156-165.
CrossRef - Ventura, S.P.M., R.L.F. de Barros, J.M. de Pinho Barbosa, C.M.F. Soares, A.S. Lima and J.A.P. Coutinho, 2012. Production and purification of an extracellular lipolytic enzyme using ionic liquid-based aqueous two-phase systems. Green Chem., 14: 734-740.
CrossRefDirect Link - Wan Omar, W.N.N., N. Nordin, M. Mohamed and N.A.S. Amin, 2009. A two-step biodiesel production from waste cooking oil: Optimization of pre-treatment step. J. Applied Sci., 9: 3098-3103.
CrossRefDirect Link - Yadav, K.N., M.G. Adsul, K.B. Bastawde, D.D. Jadhav, H.V. Thulasiram and D.V. Gokhale, 2011. Differential induction, purification and characterization of cold active lipase from Yarrowia lipolytica NCIM 3639. Bioresour. Technol., 102: 10663-10670.
CrossRefPubMedDirect Link - Yan, J., Y. Yan, S. Liu, J. Hu and G. Wang, 2011. Preparation of cross-linked lipase-coated micro-crystals for biodiesel production from waste cooking oil. Bioresource Technol., 102: 4755-4758.
PubMed - Zaks, A. and A.M. Klibanov, 1985. Enzyme-catalyzed processes in organic solvents. Proc. Natl. Acad. Sci. USA., 82: 3192-3196.
Direct Link - Zheng, Y.Y., X.H. Guo, N.N. Song and D.C. Li, 2011. Thermophilic lipase from Thermomyces lanuginosus: Gene cloning, expression and characterization. J. Mol. Catal. B: Enzym., 69: 127-132.
CrossRef - Sharma, R., Y. Chisti and U.C. Banerjee, 2001. Production, purification, characterization and applications of lipases. Biotechnol. Adv., 19: 627-662.
CrossRef