
Victoria Olusola Adetunji
Department of Veterinary Public Health and Preventive Medicine, University of Ibadan, Ibadan, Nigeria
LiveDNA: 234.1198
Ismail Ayoade Odetokun
Department of Veterinary Public Health and Preventive Medicine, University of Ibadan, Ibadan, Nigeria
American Journal of Food Technology
Year: 2012 | Volume: 7 | Issue: 10 | Page No.: 582-595







ABSTRACT
The aim of this study was to assess biofilm formation by some pathogenic strains of E. coli and Salmonella sp. using glass and cement coupons, varying nutrient-culture media and two incubation temperatures (refrigeration (11°C) and room (28±2°C)). The Crystal Violet Binding Assay (CVBA) and Bacteria Cell Enumeration (BCE) methods were used to quantify E. coli and Salmonella biofilms in 5 days while Scanning Electron Microscopy (SEM) was utilized in viewing the formed biofilms. Though, it was observed that all tested organisms did not grow at the same rate, generally, Salmonella sp. produced higher biofilms than E. coli O157:H7 strains. More biofilms were formed on cement surfaces than glass and was confirmed with SEM. Biofilms developed on the coupons were higher at room temperature and in most cases higher in media with increased nutrient concentrations. With these results, it can be deduced that there is a higher presence of biofilms formed by pathogenic organisms in our food processing plants where the principles of hygiene are not held in high esteem. Therefore, regular evaluation of operation cleanliness in the food processing environments is highly necessary in order to control these pathogens and their biofilms.
PDF Abstract XML References Citation
Received: May 06, 2012;
Accepted: August 04, 2012;
Published: October 09, 2012
How to cite this article
Victoria Olusola Adetunji and Ismail Ayoade Odetokun, 2012. Assessment of Biofilm in E. coli O157:H7 and Salmonella Strains: Influence of Cultural Conditions. American Journal of Food Technology, 7: 582-595.
DOI: 10.3923/ajft.2012.582.595
URL: https://scialert.net/abstract/?doi=ajft.2012.582.595
DOI: 10.3923/ajft.2012.582.595
URL: https://scialert.net/abstract/?doi=ajft.2012.582.595
INTRODUCTION
Common pathogenic bacteria that may be causes of foodborne diseases include strains of Salmonella and Escherichia coli (Sockett, 1991). Many high risk pathogens that cause diseases in humans are transmitted through various food items (Hassanain, 2008). E. coli O157:H7 infection has considerable economic impacts. Escherichia coli O157:H7 has emerged with increasing frequency as a foodborne pathogen of concern over the last 20 years and is responsible for causing serious illness and severe sequelae in susceptible humans (Bacon and Sofos, 2003). Escherichia coli O157:H7 was first identified as being responsible for causing outbreaks of enteric infection in 1982 (Riley et al., 1983). Symptoms include diarrhoea, haemorrhagic colitis and haemolytic uraemic syndrome (Doyle, 1991; Doyle et al., 2001). Salmonella is ubiquitous geographically and zoologically (Hirsh, 2005). The occurrence of Salmonella can contaminate food anywhere along the farm to fork continuum (D’Aoust, 1997; Small et al., 2006). Poultry products are regarded as the primary vehicles of Salmonella transmission (Geornaras and von Holy, 2000). Ready-to-eat products are typically contaminated during post-processing steps (Mbandi and Shelef, 2002). Post-processing contamination is largely contributed to poor handling practices (De Cesare et al., 2003). According to FDA the number of cases of salmonellosis is 2-4 millions/year in the US and the frequency is rapidly increasing. Especially S. enteritides is rapidly spreading in US and Europe (Enfors, 2008). All foodborne salmonellosis infections are non-typhoidal (Bailey and Maurer, 2005) and this is a major cause of food borne illness (Hassanain, 2008). Non-typhoidal salmonellosis caused more outbreaks and cases of food poisoning than all other bacteria between 1993 and 1997 (CDC, 2000). Escherichia coli O157:H7 is an emerging food pathogen that was first identified as a cause of illness in 1982 (Riley et al., 1983). Rangel et al. (2005) and CDC (2000) reported E. coli O157:H7 to be responsible for about 73,000 illnesses, 2,000 hospitalizations and 60 deaths in the United States each year. A biofilm can be defined as an assemblage or population of microbial cells that is irreversibly associated (not removed by gentle rinsing with a surface) and enclosed in a self-produced matrix of primarily polysaccharide material having possessed the ability to stick to wet surfaces while undergoing a multi-step developmental process (Abee et al., 2011; Donlan, 2002; Kokare et al., 2009; Stepanovic et al., 2004; Van Houdt and Michiels, 2010). Biofilm formed in food processing environments is of special importance as it has the potential to act as the chronic source of microbial contamination that may lead to food spoilage or transmission of diseases (Stepanovic et al., 2004). Attachment and biofilm formation by food-borne pathogens and spoilage microorganisms on food contact surfaces in processing plants are a public health and cross-contamination concern (Ryu and Beuchat, 2005). Biofilm formation comprises a sequence of steps (Breyers and Ratner, 2004). Formation of microbial biofilms on food contact surfaces is a matter of great concern for the food processing industries (Jeyasekaran et al., 2000). Biofilms formed on surfaces could develop under static conditions (i.e., on floors, walls, etc.) or under dynamic conditions (all situations in which some or all parts are moving). In general, biofilm formation involves attachment, colonization and growth of microorganisms (Forsythe, 2000). Biofilms formed by Escherichia coli O157:H7 on inadequately cleaned and sanitized contact surfaces may be a source of contamination of ground beef and deli meat in processing facilities as well as in food service settings (Sharma et al., 2005). Food processing operations usually take place in wet environment and therefore, ideal for biofilm formation. There is little information on the presence of Salmonella in biofilms in food processing environments. However, various studies (Helke and Wong, 1994; Jones and Bradshaw, 1997; Joseph et al., 2001) show that Salmonella can attach and form biofilms on surfaces found in food processing plants, including plastic, cement and stainless steel. Microaerophilic and CO2-rich conditions provide the best environment for biofilm formation while the least biofilm was formed under anaerobic conditions (Stepanovic et al., 2003). In food systems, the attachment of microorganisms leading to the formation of biofilms may be undesirable and also detrimental (Kumar and Anand, 1998). Biofilms formed in food-processing environments are of special importance as they have the potential to act as a persistent source of microbial contamination (Van Houdt and Michiels, 2010). There are not many reports on biofilm assays in developing countries especially Nigeria. Processing of food animals in most processing plants in Nigeria is far from the ideal and thus the problem of food contamination and diseases transmission are always in manifest. In most abattoirs, cattle, goats, sheep and pigs are slaughtered on bare floor usually made of cement. Coupled with water insufficiency, poor facility designs and irregular inspection and regulation, blood from slaughtered animals usually stand as source of food contamination. It is noteworthy to assay biofilms produced from food contact surfaces in order to pave a way for bacteria control. This study therefore, assessed the pattern of biofilm formation in pathogenic E. coli and Salmonella sp. from animal sources on cement and glass surfaces at 11 and 28±2°C.
MATERIALS AND METHODS
Test strains: Two strains each of E. coli O157:H7 and Salmonella sp. laboratory stock cultures from poultry and soft cheese were used for biofilm assays (Table 1) between the months of November, 2010 and June, 2012.
Biofilm development: Methods as described by Jeyasekaran et al. (2000) and Joseph et al. (2001) with slight modifications were utilized for the formation of biofilms on food contact surfaces (cement and glass). The cement (locally made, 1x2 cmx1.5 mm, PureChem® Cement, Nig.) coupons were each scrubbed with a brush and wash with distilled water thoroughly. Thereafter they were rinsed with 95% Ethanol (AnalaR®, BDH Chemical Ltd., UK), wrapped in foil paper and allowed to dry. The glass (1x2 cmx1.2 mm, Sail brand®, CAT. No. 7101) coupons were washed with detergent (Omo®) to remove grease, rinsed thoroughly with distilled water, wrapped in foil paper and sterilized in the Hot Air Oven (Elektro, HELIOS, Sweden) for 30 min at 120°C. Five different types of media were used for biofilm incubation and they include: Tryptone Soy Broth (TSB) (LAB M, Lancashire, UK), TSB+0.5% glucose, TSB+1% glucose, TSB+2% Sheep blood and TSB+5% Sheep blood. Tryptone Soy Broth (TSB) (LAB M, Lancashire, UK) was used and the producer’s direction was followed in the preparation. Thirty grams of the broth powder was dispersed in 1000 mL of distilled water, soaked for 10 min swirled to mix and warmed to dissolve before sterilized by autoclaving at 121°C for 15 min. The broth was allowed to cool before being used. After the Tryptone Soy Broth (TSB) (LAB M, Lancashire, UK) was prepared as described above, 0.5 and 1.0 g of Glucose (D+) (M and B, UK) as well as 2 and 5 mL Sheep Blood (obtained from apparently healthy sheep from the Sheep and Goat slaughter slab Bodija abattoir) were added to 100 mL of TSB for each particular test strain. These mixtures were thoroughly stirred together. Each test strains were grown in TSB (LAB M, Lancashire, UK) for 24 h at 37°C before they were added to the various media used for incubation and biofilm development. Glass jars were used for the incubation of the test strains at both refrigeration (11°C) and room (28±2°C) temperatures. Glass (1x2 cmx1.2 mm, Sail brand®, CAT. No. 7101) and cement coupons (locally made, 1x2 cmx1.5 mm, PureChem® Cement, Nig.), 13 each were placed in glass jars and 100 mL of the five different media were added. Two milliliter each of the test strain grown in TSB for 24 h at 37°C were then added to the glass jars containing each different media and the coupons. After incubation at refrigeration (11°C) (in the refrigerator (Haier Thermocool, HPZ, Ltd.) and ambient (28±2°C) temperatures for 48 h, the coupons were aseptically removed, washed with distilled water to remove unattached cells and freshly prepared broths were poured into the glass jar (Ren and Frank, 1993). This procedure was repeated on day 4 to complete biofilm formation on day 5. Five glass jars containing the representative media were used as control in which no test strain was added.
Biofilm quantification: To quantify the biofilms formed by the bacteria sp. incubated in the five different broths for 5 day at refrigeration (11°C) and ambient temperatures (28±2°C), the Crystal Violet Binding Assay (CVBA) and Bacteria Cell Enumeration (BCE) methods were used to quantify the biofilms while Scanning Electron Microscopy (SEM) was used to view the growth of the biofilms on the coupons.
| Table 1: | List, serotype and sources of strains utilized for biofilm development |
 | |
Crystal violet binding assay (CVBA): This was carried out as performed by Adetunji and Adegoke (2008) and Stepanovic et al. (2004) with some modifications. A coupon, with its replicate, was picked from each incubated broth and washed 3 times with 5 mL of distilled water. The sessile bacterial cells (biofilms) were fixed with 1 mL of 70% ethanol (AnalaR®, BDH Chemical Ltd., UK) for 15 min. The fixed samples were then air dried and stained with crystal violet. Excess stains were rinsed with running tap water and then air dried. Three milliliter of glacial acetic acid (AnalaR®, BDH Chemical Ltd., UK) was used to re-solubilize each coupon. The re-solubilized liquid was then poured into a cuvette whose wavelength was measured using the Absorption Spectrophotometer (Springfield, UK). Absorbance (OD) reading against the control for a specific media and incubation temperature type was then measured. Rate of absorbance was measured at λ: 520 and 570 nm for the E. coli and for Salmonella sp., respectively.
Bacterial cell enumeration (BCE): To enumerate biofilm cells after 5 days of incubation, the samples, with their replicates, were washed with sterile distilled water to remove unattached cells and the biofilm cells were removed by swabbing with sterile cotton swabs. Sterile peptone water were added into the swabs, shaken vigorously and enumerated by standard spread plate technique with sterile dilution performed to 10-4. Trytone Soy Agar (TSA, Scharlau, Spain) was used for enumeration and plates were incubated at 37°C for 48 h. Biofilm colonies were counted and converted to log CFU cm-2.
Scanning electron microscopy (SEM): Each coupon was picked from each incubated broth and washed 3 times with 5 mL of distilled water. The sessile bacterial cells (biofilms) were fixed with 1 mL of 70% ethanol (AnalaR®, BDH Chemical Ltd., UK) for 15 min. The fixed samples were then air dried, wrapped with foil paper and transported to the, Advanced Physics Laboratory, Sheda Science and Technology Complex (SHETSCO), Abuja, Nigeria to carry out the SEM procedure. The samples for SEM analysis were placed inside the Scanning Electron Microscope (SEM) (Zeiss EVO MA10 Carl Zeiss SMT AG, Germany) and micrographs were taken at a magnification of 1000 μm.
Statistical analysis: The statistical software SPSS for Windows, version 17.0 (SPSS Inc. Chicago, IL) was used. The T-Test was utilized to ascertain the significant differences between the biofilm formed on each coupons for each strain at the two incubation temperatures in all culture types. p values less than 0.05 were considered significant.
RESULTS
Biofilm formation (crystal violet binding assay) by E. coli O157:H7 and Salmonella strains at refrigeration temperature (11°C) on glass and cement coupons (day 5): With crystal violet binding assay, pathogenic E. coli biofilms formed on cement were higher, significantly than produced on glass coupons for all broth types (Table 2, Fig. 1-2). There was increased biofilm formation for both E. coli strains with increased nutrient in the growth media (Table 2) higher than the biofilm grown using TSB medium alone. Biofilms formed best at refrigeration temperature in TSB+0.5% g on both coupons (0.285±0.001 nm in glass and 0.463±0.001 nm in cement).
| Table 2: | Biofilm quantification (crystal violet binding assay): E. coli O157:H7 and Salmonella strains at refrigeration temperature (11°C) on glass and cement coupons (day 5) |
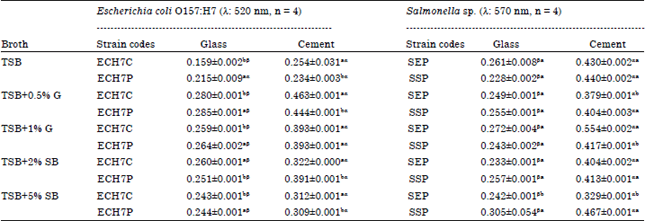 | |
| Values are Mean±SD, ECH7C: E. coli O157:H7 from soft cheese, ECH7P: E. coli O157:H7 from poultry, SEP: Salmonella Enteritidis, SSP: Salmonella spp. Values with the same superscripts (alphabets i.e., aaorbb) between strains for a specific broth type are not statistically significant at p<0.05 along the columns. Values with the same superscripts (symbols i.e., ααorββ) for the coupons on the rows are not statistically significant at p<0.05 | |
 |
| Fig. 1: | E. coli O157:H7 biofilms quantified on glass coupon |
For each strain, there was a difference between the biofilms formed on cement and glass coupons statistically significant a p<0.05 for all broths used. There was a statistical difference significant at p<0.05 in the biofilm quantified between strains on both coupons for each medium except for that noticed in TSB+1% g quantified on cement coupon. Salmonella biofilms developed on cement is significantly higher than generated on glass coupons for all broth types (Table 2, Fig. 3-4). Generally for the Salmonella strains, there is an increase in the quantity of biofilms produced with increased nutrient medium (Table 2). These strains produced biofilms higher than the E. coli O157:H7 strains. Of the two Salmonella strains, Salmonella sp. (SSP) (0.305±0.054 nm), incubated in TSB+5%SB medium formed the highest biofilms on glass coupon while on cement, Salmonella Enteritidis (0.554±0.002 nm) incubated in TSB+1%G medium was the highest biofilm former at refrigeration temperature. For each strain, there is a statistical difference between the biofilms developed on glass and cement coupons significant at p<0.05.
 |
| Fig. 2: | E. coli O157:H7 biofilms quantified on cement coupon |
 |
| Fig. 3: | Salmonella biofilms quantified on glass coupon |
Biofilm formation (crystal violet binding assay) by E. coli O157:H7 and Salmonella strains at room temperature (28±2°C) on glass and cement coupons (day 5): At room temperature (28±2°C), higher biofilm levels for both E. coli O157:H7 and Salmonella strains were quantified than at refrigeration temperature (11°C). On glass coupons, E. coli O157:H7 biofilm formed in the blood media were higher than produced in TSB only on both coupons but a lower quantity was produced in media containing with glucose (Table 3). Biofilm production with increased nutrient concentrations for E. coli O157:H7 on the cement coupons was not consistent. For each strain, there was a difference between the biofilms formed on both coupons statistically significant a p<0.05 for all broths used. Statistical significance was observed between the two E. coli O157:H7 strains for all broth on glass coupons. This was the same as observed for cement except for the coupon incubated in TSB+1% g.
| Table 3: | Biofilm quantification (crystal violet binding assay): E. coli O157:H7 and Salmonella Strains at room temperature (28±2°C) on glass and cement coupons (day 5) |
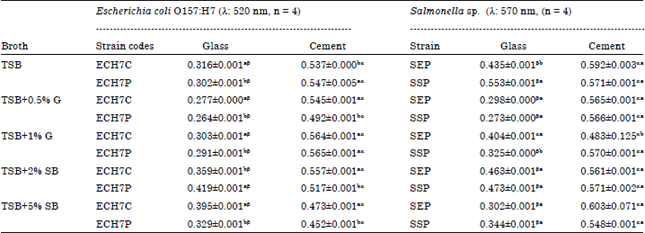 | |
Values are Mean±SD, ECH7C: E. coli O157:H7 from soft cheese, ECH7P: E. coli O157:H7 from poultry, SEP: Salmonella Enteritidis, SSP: Salmonella sp. Values with the same superscripts (alphabets i.e., aaorbb) between strains for a specific broth type are not statistically significant at p<0.05 along the columns. Values with the same superscripts (symbols i.e., ααorββ) for the coupons on the rows are not statistically significant at p<0.05 | |
 |
| Fig. 4: | Salmonella biofilm quantified on cement coupon |
Salmonella strains were better biofilm formers than E. coli O157:H7. Salmonella sp. (SSP) at room temperature (28±2°C) developed the highest level of biofilms (0.553±0.001 nm) on glass coupon in TSB broth while Salmonella Enteritidis developed the highest biofilms (0.603±0.071 nm) on cement coupon when incubated in TSB+SB broth (Table 3). Biofilm development increased with additional glucose levels though higher biofilm level was seen with 2% SB, this decreased when additional blood quantity was added to the TSB broth. Statistically, significant differences (p<0.05) were observed between both coupons for each strain in all broth types. Our results show that Salmonella sp. are better biofilm formers than E. coli O157:H7 strains.
| Table 4: | Biofilm quantification (Bacteria Cell Enumeration): E. coli O157:H7 and Salmonella Strains at refrigeration temperature (11°C) on glass and cement coupons (Day 5) |
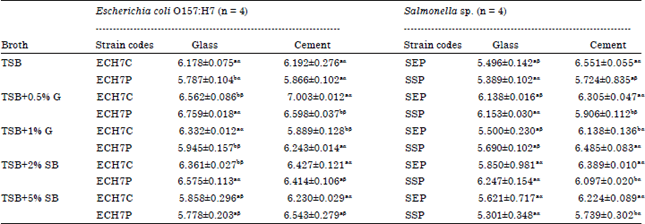 | |
Values are Mean±SD, ECH7C: E. coli O157:H7 from soft cheese, ECH7P: E. coli O157:H7 from poultry, SEP: Salmonella Enteritidis, SSP: Salmonella sp. Values with the same superscripts (alphabets i.e., aaorbb) between strains for a specific broth type are not statistically significant at p<0.05 along the columns. Values with the same superscripts (symbols i.e., ααorββ) for the coupons on the rows are not statistically significant at p<0.05 | |
Biofilm formation (bacterial cell enumeration) by E. coli O157:H7 and Salmonella strains at refrigeration temperature (11°C) on glass and cement coupons (day 5): Biofilms produced by E. coli O157:H7 strains on glass surfaces at refrigeration temperature ranged from 5.778±0.203-6.759±0.018 log CFU cm-2 formed by the poultry isolate (Table 4). On cement coupons, the biofilms developed by the strains ranged from 5.866±0.102-7.003±0.012 log CFU cm-2. Highest biofilms on glass and cement surfaces for E. coli O157:H7 were formed by ECH7P and ECH7C incubated in TSB+0.5% g, respectively. On both coupons, biofilms increased (>0.5 log CFU cm-2) with the initial concentration of nutrients (TSB+0.5% g and TSB+2% SB) added to the TSB broth but with further amplified concentrations (TSB+1%G and TSB+5% SB), biofilm formation decreased. Statistically, significant differences at p<0.05 were noticed between biofilms formed by the two E. coli O157:H7 strains on glass surfaces incubated in all broth types except in TSB+5% SB. On the other hand, on cement coupons, there were no significant differences between the two strains except with coupons incubated in the media with glucose. Biofilms developed by Salmonella strains (SEP and SSP) on glass and cement food contact surfaces at 11°C were presented in Table 4. Averagely, biofilms quantified on cement surfaces were higher than on glass. Increased biofilm production was enumerated on coupons (higher than incubated in TSB media only) with additional nutrients (0.5% g and 2% SB). But with further increase in the concentrations of these nutrients, Salmonella biofilm production decreased. Biofilms formed by Salmonella strains on glass surfaces spanned from 5.301±0.348 (produced by SSP incubated in TSB+2% SB) to 6.247±0.154 (formed by SSP incubated in TSB+5% SB) log CFU cm-2. On cement, the highest biofilm levels were recorded from both strains incubated in TSB only. No significant differences at p<0.05 were noticed between the two Salmonella strains for each broth type quantified on glass surfaces but there were significant differences (p<0.05) between biofilms quantified from the strains on cement coupons for each broth type except in TSB media. Statistical differences significant at 0.05 level of error were observed between food contact surfaces for each strain incubated in TSB media only (except SSP incubated in TSB) and TSB+G. No significant differences were however, seen with the media containing sheep blood.
| Table 5: | Biofilm quantification (bacteria cell enumeration): E. coli O157:H7 and Salmonella strains at room temperature (28±2°C) on glass and cement coupons (day 5) |
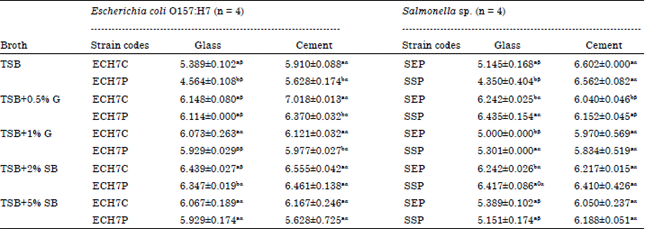 | |
Values are Mean±SD, ECH7C: E. coli O157:H7 from soft cheese, ECH7P: E. coli O157:H7 from poultry, SEP: Salmonella Enteritidis, SSP: Salmonella sp. Values with the same superscripts (alphabets i.e., aaorbb) between strains for a specific broth type are not statistically significant at p<0.05 along the columns. Values with the same superscripts (symbols i.e., ααorββ) for the coupons on the rows are not statistically significant at p<0.05 | |
Biofilm formation (bacteria cell enumeration) by E. coli O157:H7 and Salmonella strains at room temperature (28±2°C) on glass and cement coupons (day 5): Biofilms quantified from E. coli O157:H7 strains on glass and cement surfaces are presented in Table 5 at ambient temperature. Biofilms developed on glass surfaces ranged from 4.564±0.108 log CFU cm-2 (by ECH7P incubated in TSB medium only) to 6.439±0.027 log CFU cm-2 (by ECH7C grown in TSB+2%SB). The limit on cement coupons was from 5.628±0.725 log CFU cm-2 (formed by ECH7P in TSB+5%SB) to 7.018±0.013 log CFU cm-2 (developed by ECH7C in TSB+0.5%G). Supplementary nutrients to the TSB broth yielded increased biofilms formed by the strains on both coupons. Despite the increase, the extra concentrations for both coupons generated lower biofilms than the initial concentrations of TSB+0.5%G and TSB+2%SB. For each broth type, there were no statistical significant differences at p<0.05 in biofilms developed between strains on a particular glass surface except in TSB and TSB+5%SB while on cement coupons, significant differences were not observed with the sheep blood media. On the average, lower biofilms were developed on glass contact surfaces than on cement coupons for Salmonella strains incubated at ambient temperature (Table 5). The biofilms formed by these strains on glass coupons vary extending from 4.350±0.404 (SSP biofilms incubated in TSB) to 6.242±0.025 log CFU cm-2 (SEP biofilms grown in TSB+0.5%G). For cement surfaces, the span was from 5.834±0.519 (SSP biofilms quantified from TSB+1%G) to 6.602±0.000 log CFU cm-2 (SEP biofilms developed in TSB medium only). Generally, with the initial increase in nutrient concentrations, increased biofilm production was observed for both media containing the 0.5%G and 2%SB. The increased biofilm formation was however, reduced with further increase in the concentrations of both nutrients. There were statistical differences at p<0.05 between the biofilms developed by the two Salmonella strains on glass contact surfaces for each broth type with the exception of the strains incubated in TSB+5%SB. For the cement coupons, statistical significance (p<0.05) was only observed between the strains incubated in TSB+0.5%G. Between the food contact surfaces, significant differences (at 0.05 level of error) were not observed with SSP (incubated in TSB+1%SB) and both strains incubated in TSB+2%SB.
DISCUSSION
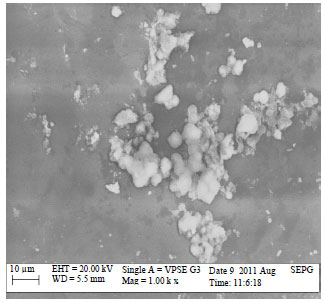
Biofilm formation on glass and cement coupons: Though cement surfaces are widely used in most processing environment in the Nigerian food industry, glass is increasingly being utilized recently, few reports are available locally on biofilms developed on glass (Adetunji and Adegoke, 2008; Adetunji and Isola, 2011) with none assayed on cement surfaces. The significantly higher biofilms (p<0.05) developed on cement (also demonstrated by the Scanning Electron Micrographs) noticed in all test strains was as a result of several factors. This finding is in agreement with earlier studies that reported that biofilms are easily developed on static than dynamic surfaces and may be formed in all areas of food processing environments (Donlan, 2002; Sofos, 2009). The hydrophobicity of bacteria and surface which are important criteria in biofilm formation (Manijeh et al., 2008) also supports the findings in this study. Cement is more hydrophilic than glass which thus facilitates bacteria adhesion and subsequent biofilm formation. The surface characteristics of the cement surfaces also support the higher biofilm levels observed than in glass (Donlan, 2002; Faille et al., 2002; Jullien et al., 2003; Ryu and Beuchat, 2005). The contact time (5 days) between the bacteria cells and the coupons was sufficient enough to allow easy irreversible adhesion of the cells and subsequent biofilm formation. The strains in this study have also been shown to possess fimbriae, flagella, exopolysaccharides and curli which are important extracellular surface structures produced by bacteria which may affect their attachment on food contact surfaces and consequent biofilm formation (Van Houdt and Michiels, 2010). Type 1 fimbriae have been demonstrated to play critical role in initial stable cell-to-surface attachment for Salmonella enterica serovar Enteritidis (Austin et al., 1998) during biofilm formation. Some poultry isolates of Salmonella were found to be biofilm formers on cement surfaces (Joseph et al., 2001) while also, other Escherichia coli strains such as Escherichia coli O111 have been confirmed to survive on cement surfaces forming biofilms (Ghazani et al., 2009). These findings also corroborate the biofilm forming ability of the strains in this study on cement.
Using five culture media with increased concentrations: Results of this study indicated in most cases that with additional nutrients concentrations, the biofilms quantified on the food contact surfaces increased. Higher levels of biofilms were formed with glucose than blood though higher than biofilms grown in TSB broths only. The increased biofilm proliferation was expected because biofilm production increased with nutrients. This finding is in agreement with earlier study where it was reported that sequestration to a nutrient-rich area (colonization) is one of the reasons that drive bacteria to produce biofilms (Jefferson, 2004). In general EPS production increased under conditions where growth was extended by the high glucose content in the medium (Fleming and Wingender, 2001) which is in agreement with this study. In addition E. coli strains have been shown to be strong biofilm formers with increased glucose concentrations (Rodrigues et al., 2010). Though, it has since been found that initial attachment, detachment and characteristics of biofilms of E. coli O157:H7 were affected by the nutrient status of the medium in which the biofilm was developed (Dewanti and Wong, 1995). The findings in this study is nearly at par with earlier report since with increase in the glucose concentration from 0.04% to 0.1 and subsequently 1% there was no significant effect on the number of adherent bacteria as quantified by plate counts (Dewanti and Wong, 1995). A positive correlation between glucose and biofilm formation in Salmonella Enteritidis has been established (Bonafonte et al., 2000) which is in agreement with findings in this study. On the contrary, trehalose and mannose which are disaccharides allow poor biofilm formation (Chmielewski and Frank, 2003). Generally, Salmonella sp. produced more biofilm in nutrient-poor medium grown on plastic surfaces (Stepanovic et al., 2004), this is at par with findings in this study, although the attachment surface was glass and cement in this study.
There is a strong indication that all bacteria strains do not grow at the same rate as seen in the result. The disparity in biofilm developed both in varying glucose and blood concentrations might be due to various factors. biofilm formation and adherence are not accomplished by the same mechanism in the different media; the strains probably utilize different strategies and express different biofilm-promoting genes depending on available environmental resources (Hancock et al., 2011). Biofilm development in blood concentrations is scare and thus, we were unable to compare our results with other findings. The reduced biofilm quantified by some E. coli and Salmonella strains might be due to the toxic influence of blood digestion molecules after blood breakdown on the biofilms. Increase repulsive forces between bacteria cells and the surfaces might result from the actions of these molecules.
Incubating at refrigeration (11°C) and room (28±2°C) temperatures: The results generated in this study clearly demonstrated that higher biofilm levels were formed at ambient (28±2°C) than at refrigeration (11°C) temperature. Increased curli production in E. coli O157:H7 strains might be responsible for the higher biofilm development noticed at room than at refrigeration temperature. Curli production by E. coli O157:H7 enhances its ability to form biofilm (Ryu et al., 2004) and these curli increases at higher temperatures. Generally, the optimal cultivation temperature for the production of most EPS molecules was estimated between 26 and 31°C (Gandhi et al., 1997; Lory, 1992) thus, subsequently enhancing biofilm formation at ambient temperature. This can explain the increase in biofilm formation at 28±2°C in this study.
Higher biofilms levels were also recorded in this study at room temperature (28±2°C) for the Salmonella isolates tested. The capacity shown for biofilm production by Salmonella sp. at room temperature as a result is of particular interest to the food industry which again emphasizes the necessity for regular and appropriate cleaning (Stepanovic et al., 2003). Though some observers have stated that Salmonellae display multicellular-characteristics in terms of biofilm formation, this multi-cellular behaviour is associated with biofilm formation at various incubation temperature (Stepanovic et al., 2003) and is regulated by environmental conditions that target the agfD promoter (Gerstel and Romling, 2001; Romling et al., 2000). Expression of agfD leads to the production of polymers, thin aggregative fimbriae and cellulose which form the extracellular matrix (Gerstel and Romling, 2001).
It has been proposed that the increased hydrophobicity at high temperatures (e.g., 37°C) may enhance the initial cell adherence, contributing to a higher biofilm density (Di Bonaventura et al., 2008). However, biofilm cells may generate and secrete more extracellular polymeric substances in response to temperature and other factors which would also be seen as an increase in Crystal Violet (CV) absorbance in the microplate assay (Pan et al., 2010). More importantly, it should be noted that the optimum temperature for a microorganism is associated with an increase in nutrient intake resulting in a rapid formation of biofilm (Stepanovic et al., 2003).
CONCLUSION
In conclusion, the results of this study demonstrate that the pathogens under study did not grow at the same rate and higher incubation temperatures and nutrient influenced biofilm formation. In addition E. coli O157:H7 strains were observed to be lower biofilm formers when compared to Salmonella sp. Cement surfaces also adhered more biofilms than glass surfaces. Of particular significance in this study is the high biofilm forming ability of these foodborne isolates, thereby increasing the risk of bacterial contamination in food processing. Further studies on the ability of these pathogens to form biofilms on ceramic are necessary since it is a more durable material than glass in food processing environments.
REFERENCES
- Abee, T., A.T. Kovacs, O.P. Kuipers and S. van der Veen, 2011. Biofilm formation and dispersal in Gram-positive bacteria. Curr. Opin. Biotechnol., 22: 172-179.
PubMed - Adetunji, V.O. and G.O. Adegoke, 2008. Formation of biofilm by strains of Listeria monocytogenes isolated from soft cheese wara and its processing environment. Afr. J. Biotechnol., 7: 2893-2897.
Direct Link - Adetunji, V.O. and T.O. Isola, 2011. Crystal violet binding assay for assessment of biofilm formation by Listeria monocytogenes and Listeria spp on wood, steel and glass surfaces. Global Vet., 6: 6-10.
Direct Link - Austin, J.W., G. Sanders, W.W. Kay and S.K. Collinson, 1998. Thin aggregative fimbriae enhance Salmonella Enteritidis biofilm formation. FEMS Microbiol. Lett., 162: 295-301.
CrossRef - Bonafonte, M.A., C. Solano, B. Sesma, M. Alvarez, L. Montuenga, D. Garcia-Ros and C. Gamazo, 2000. The relationship between glycogen synthesis, biofilm formation and virulence in Salmonella enteritidis. FEMS Microbiol. Lett., 191: 31-36.
PubMed - CDC, 2000. Surveillance for foodborne - disease outbreaks United States, 1993-1997. MMWR Morb. Mortal. Weekly Rep., 49: 1-72.
Direct Link - Chmielewski, R.A.N. and J.F. Frank, 2003. Biofilm formation and control in food processing facilities. Comprehens. Rev. Food Sci. Food Safe., 2: 22-32.
CrossRefDirect Link - De Cesare, A., B.W. Sheldon, K.S. Smith and L.A. Jaykus, 2003. Survival and persistence of Campylobacter and Salmonella species under various organic loads on food contact surfaces. J. Food Prot., 66: 1587-1594.
PubMed - Dewanti, R. and A.C.L. Wong, 1995. Influence of culture conditions on biofilm formation by Escherichia coli O157:H7. Int. J. Food Microbiol., 26: 147-164.
CrossRefPubMedDirect Link - Di Bonaventura, G., R. Piccolomini, D. Paludi, V. D'Orio, A. Vergara, M. Conter and A. Ianieri, 2008. Influence of temperature on biofilm formation by Listeria monocytogenes on various food‐contact surfaces: Relationship with motility and cell surface hydrophobicity. J. Applied Microbiol., 104: 1552-1561.
CrossRefPubMedDirect Link - Donlan, R.M., 2002. Biofilms: Microbial life on surfaces. Emerg. Infect. Dis., 8: 881-890.
CrossRefPubMedDirect Link - Doyle, M.P., 1991. Escherichia coli O157: H7 and its significance in foods. Int. J. Food Microbiol., 12: 289-302.
PubMed - Faille, C., C. Jullien, F. Fontaine, M.N. Bellon-Fontaine, C. Slomianny and T. Benezech, 2002. Adhesion of Bacillus spores and Escherichia coli cells to inert surfaces: Role of surface hydrophobicity. Can. J. Microbiol., 48: 728-738.
PubMed - Flemming, H.C. and J. Wingender, 2001. Relevance of microbial Extracellular Polymeric Substances (EPSs)-Part I: Structural and ecological aspects. Water Sci. Technol., 43: 1-8.
PubMedDirect Link - Gandhi, H.P., R.M. Ray and R.M. Patel, 1997. Exopolymer production by Bacillus species. Carbohydr. Polym., 34: 323-327.
CrossRefDirect Link - Geornaras, I. and A. von Holy, 2000. Bacterial counts associated with poultry processing at different sampling times. J. Basic Microbiol., 40: 343-349.
PubMed - Gerstel, U. and U. Romling, 2001. Oxygen tension and nutrient starvation are major signals that regulate agfD promoter activity and expression of the multicellular morphotype in Salmonella typhimurium. Environ. Microbiol., 3: 638-648.
CrossRef - Ghazani, M.H.M., A.R. Karam and J. Dolgharisharaf, 2009. Biofilm Formation of Escherichia coli O111 on food contact cement surfaces. Res. J. Biomedical. Sci., 4: 113-115.
Direct Link - Hassanain, N.A., 2008. Detection of antibodies against zoonotic food borne pathogens in sera of food handlers. Global Vet., 2: 285-289.
Direct Link - Jefferson, K.K., 2004. Mini review: What drives bacteria to produce a biofilm. FEMS Microbiol. Lett., 235: 163-173.
Direct Link - Jones, K. and S.B. Bradshaw, 1997. Synergism in biofilm formation between Salmonella Enteritidis and a nitrogen-fixing strain of Klebsiella pneumoniae. J. Applied Microbiol., 82: 663-668.
PubMed - Joseph, B., S.K. Otta and I. Karunasagar, 2001. Biofilm formation by Salmonella spp. on food contact surfaces and their sensitivity to sanitizers. Int. Food Microbiol., 64: 367-372.
CrossRefDirect Link - Kokare, C.R., S. Chakraborty, A.N. Khopade and K.R. Mahadik, 2009. Biofilm: Importance and applications. Ind. J. Biol. Technol., 8: 159-168.
Direct Link - Lory, S., 1992. Determinants of extracellular protein secretion in Gram-negative bacteria. J. Bacteriol., 174: 3423-3428.
Direct Link - Manijeh, M., J. Mohammad and K.K. Roha, 2008. Biofilm formation by Salmonella enteritidis on food contact surfaces. J. Boil. Sci., 8: 502-505.
CrossRefDirect Link - Mbandi, E. and L.A. Shelef, 2002. Enhanced antimicrobial effects of combination of lactate and diacetate on Listeria monocytogenes and Salmonella spp. in beef Bologna. Int. J. Food Microbiol., 76: 191-198.
PubMed - Pan, Y., F. Breidt and L. Gorski, 2010. Synergistic effects of sodium chloride, glucose and temperature on Biofilm formation by Listeria monocytogenes Serotype 1/2a and 4b Strains. Appl. Environ. Microbiol., 76: 1433-1441.
CrossRef - Rangel, J.M., P.H. Sparling, C. Crowe, P.M. Griffin and D.L. Swerdlow, 2005. Epidemiology of Escherichia coli O157: H7 outbreaks, United States, 1982-2002. Emerg. Infect. Dis., 11: 603-609.
CrossRefDirect Link - Riley, L.W., R.S. Remis, S.D. Helgerson, H.B. McGee and J.G. Wells et al., 1983. Hemorrhagic colitis associated with a rare Escherichia coli serotype. N. Engl. J. Med., 308: 681-685.
PubMedDirect Link - Rodrigues, L.B., L.R. Santos, V.Z. Tagliari, N.N. Rizzo and G. Trenhago et al., 2010. Quantification of biofilm production on polystyrene by Listeria, Escherichia coli and Staphylococcus aureus isolated from a poultry slaughterhouse. Braz. J. Microbiol., 41: 1082-1085.
Direct Link - Ryu, J.H., H. Kim, J.F. Frank and L.R. Beuchat, 2004. Attachment and biofilm formation on stainless steel by Escherichia coli O157:H7 as affected by curli production. Lett. Applied Microbiol., 39: 359-362.
CrossRefPubMedDirect Link - Ryu, J.H. and L.R. Beuchat, 2005. Biofilm Formation by Escherichia coli O157: H7 on stainless steel: Effect of exopolysaccharide and curli production on its resistance to chlorine. Applied Environ. Microbiol., 71: 247-254.
CrossRefDirect Link - Small, A., C. James, S. James, R.H. Davies and E. Liebana et al., 2006. Presence of Salmonella in the red meat abattoir lairage after routine cleansing and disinfection and on carcasses. J. Food Prot., 69: 2342-2351.
PubMed - Sockett, P.N., 1991. The economic implications of human Salmonella infection. J. Applied Bacteriol., 71: 289-295.
Direct Link - Stepanovic, S., I. Irkovi, V. Mija and M. Vabi-Vlahovi, 2003. Influence of the incubation temperature, atmosphere and dynamic conditions on biofilm formation by Salmonella sp. Food Microbiol., 20: 339-343.
Direct Link - Stepanovic, S., I. Cirkovic, L. Ranin and M. Svabic-Vlahovic, 2004. Biofilm formation by Salmonella spp. and Listeria monocytogenes on plastic surface. Lett. Applied Microbiol., 38: 428-432.
CrossRefPubMedDirect Link - Kumar, C.G. and S.K. Anand, 1998. Significance of microbial biofilms in food industry: A review. Int. J. Food Microbiol., 42: 9-27.
CrossRefDirect Link