
Jace D. Everette
Department of Agriculture, 1200 North University Drive, 153 Woodward Hall, University of Arkansas at Pine Bluff, Pine Bluff AR 71601, USA
Shahidul Islam
Department of Agriculture, 1200 North University Drive, 153 Woodward Hall, University of Arkansas at Pine Bluff, Pine Bluff AR 71601, USA
American Journal of Food Technology
Year: 2012 | Volume: 7 | Issue: 2 | Page No.: 50-61







ABSTRACT
The antioxidant activities and phenolic contents of the five orange-fleshed sweetpotato genotypes namely SP-122, SP-129, SP-115, SP-323 and SP-425 in relation to the two extraction methods (hydrophilic and lipophilic fraction) were examined. The antioxidant capacity was investigated with three different screening methods: the 2, 2’-azinobis (3-ethyl-benzothiazoline-6-sulfonic acid) (ABTS), 2, 2-diphenyl-1-picrylhydrazyl (DPPH) and Oxygen Radical Absorbance Capacity (ORAC). It was found that the antioxidant activity of sweetpotato extracts in hydrophilic fraction have a significant antioxidant effect when tested by each method. There was a relationship between total polyphenol content and antioxidant function in case of ABTS (r = 0.59) and ORAC (r = 0.35). The hydrophilic ABTS values correlate significantly with the hydrophilic DPPH values (r = 0.84) and the hydrophilic ORAC values correlate reasonably well with the hydrophilic ABTS values (r = 0.85). In case of the hydrophilic DPPH values and hydrophilic ORAC values also showed a strong correlation (r = 0.87). However, antioxidant activities with the lipophilic extracts were not significantly correlated. Among the sweetpotato genotypes studied, the SP-129 had significantly higher antioxidant activity in ORAC and ABTS methods. In case of DPPH methods, the genotypes SP-425 showed higher activity compared to others studied; however, phenolic content of all genotypes was ranged 3.5-4.8 mg TAE/g dry weight. Among the methods examined, ABTS proved the best for antioxidant determination in orange-fleshed sweetpotatoes followed by ORAC method. The information provided by this research will also facilitate the genetic and chemical breeding study for improvement of the desired quality criteria of orange fleshed sweetpotatoes as well as other produces.
PDF Abstract XML References Citation
Received: August 04, 2011;
Accepted: November 08, 2011;
Published: December 20, 2011
How to cite this article
Jace D. Everette and Shahidul Islam, 2012. Effect of Extraction Procedures, Genotypes and Screening Methods to Measure the Antioxidant Potential and Phenolic Content of Orange-fleshed Sweetpotatoes (Ipomoea batatas L.). American Journal of Food Technology, 7: 50-61.
DOI: 10.3923/ajft.2012.50.61
URL: https://scialert.net/abstract/?doi=ajft.2012.50.61
DOI: 10.3923/ajft.2012.50.61
URL: https://scialert.net/abstract/?doi=ajft.2012.50.61
INTRODUCTION
The sweetpotato [Ipomoea batatas L. Lam.] is the seventh most important food crop in the world (FAO, 1997) and is among the crops selected by the U.S. National Aeronautics and Space Administration to be grown in a controlled ecological life support system as a primary food source (Hoff et al., 1982). These food crops have a high content of phenolic antioxidants, especially caffeic and chlorogenic acids (Teow et al., 2007; Islam, 2006, 2008a, 2009). Orange-fleshed sweet potatoes are also rich in carotenoids (Teow et al., 2007; Yoshimoto et al., 2003). Purple-fleshed sweet potatoes also have high anthocyanin content (Yang and Gadi, 2008). Sweet potatoes have also been shown to have antidiabetic (Kusano et al., 2001; Matsui et al., 2002; Islam, 2006) and antimicrobial (Islam, 2008b) properties. Sweetpotato cultivars whose roots are used for a beverage, a paste, a powder, an alcohol drink and a natural colorant have been developed in this decade (Yoshimoto, 2001; Islam, 2006).
Phenolic compounds are a diverse group of secondary metabolites present in higher plants that play important roles in the structure of plants and are involved in a number of metabolic pathways (Harbone,1980; Prior and Cao, 1999). Plant phenolics, because of their diversity and extensive distribution, can be argued to be an important group of natural antioxidants and contribute to organoleptic and nutritional qualities of fruit and vegetables. Phenolic compounds exist universally in most of the vegetables, which are also rich sources of natural antioxidants (Peluso et al., 1995; Chuda et al., 1996; Shimozono et al., 1996; Kaul and Khanduja, 1998; Yoshimoto et al., 1999a,b; Yoshimoto, 2001; Yoshimoto et al., 2003; Murayama et al., 2002; Islam et al., 2002a,b, 2003a,b, 2009; Islam, 2006, 2008a). Dietary antioxidants have attracted special attention because they can protect the human body from oxidative stress, which may cause many diseases including cancer, aging and cardiovascular diseases (Stevens et al., 1995; Hagerman et al., 1998; Kaul and Khanduja, 1998; Prior et al., 1998; Robards et al., 1999; Yoshimoto et al., 1999a, b; Yoshimoto, 2001; Yoshimoto et al., 2003; Islam et al., 2005, 2003a-c; Islam, 2008a, 2009). Therefore, sweetpotato may become an excellent source material for biologically active compounds.
Oxidative stress is known to cause chronic diseases such as cancer, Parkinson’s disease, Alzheimer’s dementia, heart disease, arthritis and many other ailments (Waris and Ahsan, 2006). Oxidative stress occurs as a result of free radicals such as peroxyl, hydroxyl and superoxide radicals (Buechter, 1988). Antioxidants found in fruits and vegetables neutralize free radicals and promote health (Ames et al., 1993). These antioxidants include anthocyanins, polyphenols, carotenoids and flavonoids (Woolfe, 1992; Islam, 2006, 2008b). Therefore, consumption of fruits and vegetables is necessary to combat oxidative stress and prevent degenerative diseases.
Studies have been done which compared the antioxidant content of white, orange, pink, red and purple-fleshed sweet potatoes. These studies indicated that purple fleshed sweet potatoes had the highest overall antioxidant content and color intensity was indicative of antioxidant capacity White fleshed sweet potatoes displayed antioxidant activity that was less than the darker fleshed varieties (Teow et al., 2007). Studies on other foods have also shown that the more highly pigmented varieties are also richer in antioxidant content (Awika et al., 2003; Bao et al., 2005). The purpose of this study is to determine the variation in extraction methods in relation to antioxidant capacity among five genotypes of orange fleshed sweetpotatoes. Also, to find out the suitable antioxidant assays method(s) for orange fleshed sweetpotatoes.
MATERIALS AND METHODS
Chemicals: Trolox (2, 5, 7, 8-tetramethylchroman-2-carboxylic acid), tannic acid, 2, 2-diphenyl-1-picryl hydrazyl (DPPH), 2, 2’-azinobis(3-ethyl-benzothiazoline-6-sulfonic acid) (ABTS), methyl-β-cyclodextrin and sodium persulfate, were purchased from Sigma-Aldrich (St. Louis, MO, USA). 2, 2’Azobis (2-amidino propane) dihydrochloride (AAPH) and fluorescein were obtained from ACROS organics (New Jersey, USA). Folin-Ciocalteau phenol reagent was purchased from MP Biomedicals All other reagents used were of analytical or HPLC grades.
Preparation of sweetpotato samples for analysis: The experiment was conducted during 2009 to 2011 at the University of Arkansas at Pine Bluff, USA. Trials were carried out over 3 years using 25 sweetpotato genotypes grown to screen for suitable genotypes with desireable characteristics. Five orange fleshed genotypes were chosen for further study for biochemical characteristics such as SP-122, SP-129, SP-115, SP-323 and SP-425 of orange-fleshed sweet potato were tested. These genotypes had similar color intensity. The sweetpotato storage roots were washed with tap water and stored at -80°C for 24 h prior to freeze drying. The bulbs were manually cut into small pieces and freeze dried for 60 h in a Millrock MD Freeze-Dryer at -54°C and 96 mT. The samples were then manually ground into a fine powder and stored at -20°C prior to extraction.
Lipophilic and hydrophilic extracts methods: Five grams of freeze-dried powder were stirred for 2 min in 25 mL of hexane and the mixture was filtered using a Büchner funnel. The hexane extraction was repeated twice and the combined lipophilic extracts were evaporated to dryness at 50°C using a rotary evaporator. The dried hexane extract was re-dissolved in 50 mL of 50% acetone/50% water containing 7% methylated β-cyclodextrin. The remaining residue was then extracted twice with 25 mL of acidified methanol (80% methanol/ 7% acetic acid). The combined hydrophilic extracts were made to 50 mL with acidified methanol. This method of extraction was developed by Teow et al. (2007).
Measured antioxidant activity in the ORAC assay: The procedure for the ORAC assay was established by Prior (2003). Trolox was dissolved in a phosphate buffer (0.075 M, pH 7.4). The phosphate buffer was used as a blank and solvent used to make necessary dilutions. Fluorescence measurements were taken using a Synergy HT Multi-Mode microplate reader (BioTek Instruments, Inc. Winooski, Vermont). The lipophilic fractions were diluted 10-fold and the hydrophilic fractions were diluted 100-fold. For the diluted samples, 20 μL were added to 200 μL fluorescein (incubated at 37°C). Then, 20 μL AAPH were added rapidly to each well via a multi-channel pipette. Immediately after addition of AAPH, the plate was agitated for 1 sec before the first reading. Readings were taken for 2 h at 1 min intervals. Excitation and emission wavelengths were set at 480 and 520 nm, respectively. The area under curves was measured using Image J software. Data were expressed as μmoles Trolox equivalent per gram dry weight (μmol TE -1 dw).
Measured radical scavenging activity in the ABTS assay: The radical-scavenging activity of the extracts was measured by following the procedure reported by Walker and Everette (2009). Stock solutions of ABTS (5.00x10-4 M) and sodium persulfate (6.89 x10-3 M) in PBS (pH = 8.0) were prepared. The ABTS radical cation solution was prepared by adding 1 mL of sodium persulfate to 99 mL of stock ABTS. The solution was stored for 16 h. This produced a solution of ABTS+ which gave an absorbance of approximately 0.85 at 734 nm. A 10 mM stock solution of Trolox was prepared for every sample tested. Dilutions were prepared for each sample tested. Dilution strength was dependent upon each extract’s relative antioxidant capacity. For each dilution, 20 μL were added to 2.5 mL of ABTS.+ solution and incubated in a dry bath at 37°C for 30 min. Absorbances were measured at 734 nm on an ASYS UVM 340 plate reader. TEAC values were measured by comparing the slopes of sample plots compared to that of Trolox.
Measured radical scavenging activity in the DPPH assay: The radical-scavenging activity of the extracts was measured according to a slightly modified method described by Brand-Williams et al. (1995). A stock solution of DPPH (6 mM) was prepared by dissolving 0.0263 g in 10 mL of ethanol (or methanol). The stock solution is diluted to prepare a 60 μM working solution. Again, a 10 mM stock solution of Trolox was prepared for every sample tested. Dilutions were prepared for each sample tested. Dilution strength was dependent upon each extract’s relative antioxidant capacity. For each dilution, 20 μL were added to 2.5 mL of DPPH. solution and incubated in a dry bath at 37°C for 30 min. Absorbances were measured at 520 nm on an ASYS UVM 340 plate reader. TEAC values were measured by comparing the slope of sample plots to the slope of Trolox. Antioxidant activity was reported as μmoles Trolox equivalent per gram dry weight sample (μmol TE g-1 dw).
Determination of total phenols: Total phenolic content of each extract was measured using a slightly modified method reported by Singleton et al. (1999). The extracts were diluted with distilled water, followed by the addition of 0.25 mL of Folin-Ciacalteau reagent (1 N). Then, 1.25 mL of sodium carbonate is added to each dilution and incubated at room temperature for 40 min. The following dilutions were made for each sample: 1:100, 1:50, 1:33, 1:25 and 1:20. The absorbance was measured at 725 nm on an ASYS UVM 340 plate reader. Distilled water was measured as the blank. Tannic acid was used as a standard. Total phenolic content was expressed as milligrams Tannic acid equivalents per gram dry weight sample (mg TAE g-1 dw). TAE were measured by comparing the slope of samples to the slope tannic acid.
Statistical analysis: The data were analyzed as a combined series of CRDs for laboratory experiments. Data were subjected to analysis of variance (ANOVA). Results are presented as means of four individual experiments. Pearson correlations and t-tests were performed in Microsoft Excel. Group differences were evaluated using t-tests with p<0.05 considered to be an indication of a statistically significant difference.
RESULTS AND DISCUSSION
Total phenols: The phenolic content of each extract was also evaluated (Fig. 1). The hydrophilic extract for SP115 demonstrated the most total phenols (4.8 mg TAE g-1 dw) whereas the SP-323 hydrophilic extract demonstrated the least (3.5 mg TAE g-1 DW). However, statistical analysis indicated the five varieties were not significantly different from one another (p>0.05). The lipophilic extract for SP-129 contained the most phenols (0.64 mg TAE g-1 DW) whereas the SP-122 lipophilic extract contained the least (0.37 mg TAE g-1 DW). Similar to the hydrophilic extracts, statistical analysis indicated the three varieties were not significantly different from one another (p>0.05). Several authors reported that highest phenolic content is typically an indication of highest antioxidant activity (Islam et al., 2003a, 2011; Islam, 2006, 2008a). The SP-129 extracts repeatedly displayed higher antioxidant activity than the other two varieties. However, the hydrophilic SP-129 extract contained a lower amount of phenols than the other two hydrophilic extracts for SP-122 and SP-115. Therefore, the phenolic content results suggest the three varieties have very similar amounts of polyphenols. Polyphenols constitute one of the most numerous and ubiquitous groups of plant metabolics and are an integral part of the human diet. Orange fleshed sweetpotatoes are known to possess a variety of antioxidant properties (Yoshinaga et al., 1999; Yoshimoto et al., 2001).
Antioxidant capacity in ABTS, DPPH and ORAC assays: The antioxidant activity of five varieties of orange fleshed sweetpotatoes was evaluated in three antioxidant assays.
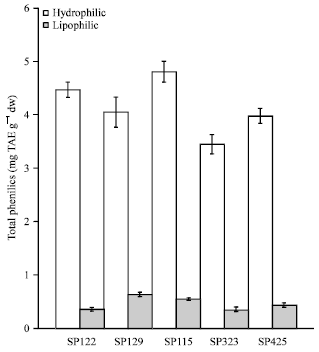 |
| Fig. 1: | Total phenolic content of sweet potato extracts (Bars indicate standard error) |
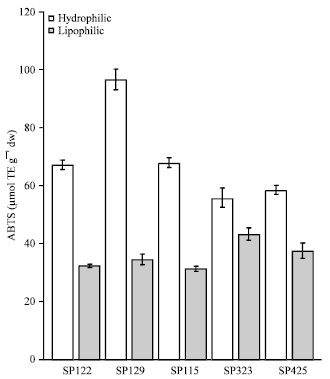 |
| Fig. 2: | Antioxidant activity of sweet potato extracts towards the ABTS radical cation (Bars indicate standard error |
Although, we have found a good correlation among all the methods used here for assessing antioxidant capacity, using more than one antioxidant assay is strongly recommended a single method will provide basic information about antioxidant properties, but a combination of methods describes the antioxidant properties of the sample in more detail. The ABTS radical scavenging capacity was examined and both fractions of SP-129 followed by SP-115 demonstrated the highest activity of the genotypes studied (p<0.05) (Fig. 2). The DPPH radical scavenging capacity was measured for the five genotypes studied (Fig. 3).
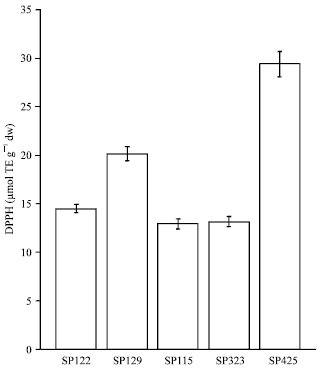 |
| Fig. 3: | Antioxidant activity of hydrophilic sweet potato extracts towards the DPPH radical (Bars indicate standard error) |
The genotypes SP-425 showed the most activity followed by SP-129. The results suggested that the lipophilic extracts were not active in the DPPH assay for orange colored sweetpotatoes. There were no significant differences among SP-122, SP-115 and SP-323 (p>0.05). In the ORAC assay, the antioxidant activity measured was again comparable to the other two assays. Further, the rank order for the hydrophilic ORAC results indicated SP-129 had the highest activity while SP-323 had the lowest. All five genotypes had statistically different (p<0.05) activities from each other (Fig. 4). The lipophilic ORAC results indicated SP-129 was also the most active variety and SP-122 was the least active. In addition, statistical analysis indicated SP-129 was significantly different from the other four varieties (p<0.05); however, SP-122 and SP-115 were not significantly different from each other (p>0.05). The antioxidant activities of the hydrophilic and lipophilic extracts ranged from 45.41 to 23.44 μmol TE g-1 dw and 1.23 to 0.723 μmol TE g-1 dw, respectively (Fig. 4). The study showed SP-129 demonstrated the greatest antioxidant activity among the five genotypes tested in all antioxidant models in both hydrophilic and lipophilic fractions. As expected, the hydrophilic fractions displayed considerably more activity than the lipophilic. The fact that SP-129 displayed considerably more activity than SP-122 and SP-115 which had similar color intensity indicated that considerable variation may occur among orange-fleshed sweet potatoes of similar color intensity.
There are many reports attempting to rank the antioxidant properties of different plant materials using different methods (Pellegrini et al., 2003; Prior et al., 2003; Huang et al., 2002, 2005) including ORAC (Ou et al., 2002a), DPPH (Islam et al., 2009), ABTS (Walker and Everette, 2009). The antioxidant capacity assays measure the combined effect of many antioxidants present in the sample which are able to scavenge free radicals generated in the assays. Interactions between antioxidant are also reflected in the assay value. Both ORAC and ABTS are indicators of the free-radical scavenging ability of antioxidant against peroxyl radical, using the same mechanism of hydrogen atom transfer.
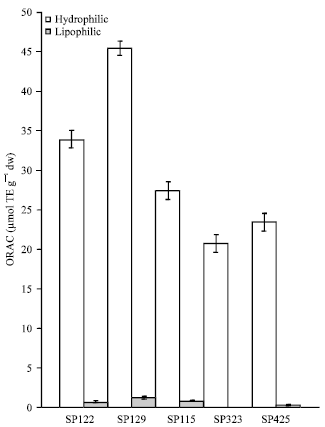 |
| Fig. 4: | Measurement of sweet potato extracts oxygen radical absorbance capacity (Bars indicate standard error) |
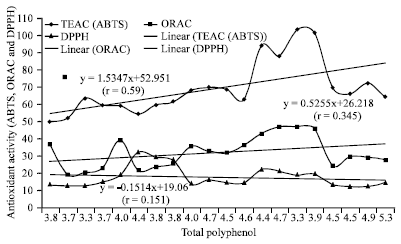 |
| Fig. 5: | Correlation between Total phenolics and different antioxidant methods (n = 20) |
On the other hand, ORAC measures the meta-chelating properties of antioxidant thereby expressing a radical prevention ability of the sample (Huang et al., 2005). Nevertheless in contrast to Ou et al. (2002b) and Huang et al. (2002), who did not find any general agreement among the antioxidant methods used, our data correlated well. A slight difference among the results obtained by the three assays might relate to the different conditions of measurements and the sensitivity of the assays.
Correlations: There was a direct relationship between total polyphenol contents antioxidant function in case of ABTS (r = 0.59) and ORAC (r = 0.35) (Fig. 5).
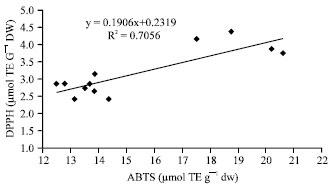 |
| Fig. 6: | Correlation between hydrophilic ABTS values and hydrophilic DPPH values |
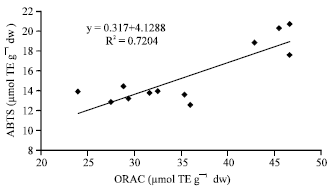 |
| Fig. 7: | Correlation between hydrophilic ORAC values and hydrophilic ABTS values |
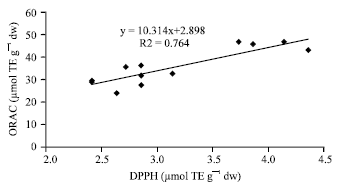 |
| Fig. 8: | Correlation between hydrophilic DPPH values and hydrophilic ORAC values |
These data are in accordance witht hat of other authors who have shown that high total polyphenol content increases antioxidant function and that there is a liner correlation between polyphenol and antioxidant activity (Gorinstein et al., 2003; Islam et al., 2003a; Islam, 2006, 2008b). The three different antioxidant methods used in this study to examine antioxidant activity were compared. The antioxidant activity seen for SP-129 was the highest in all three assays. As shown in Fig. 6 and 7, the hydrophilic ABTS values correlate reasonably well with the hydrophilic DPPH values (r = 0.84) and the hydrophilic ORAC values correlate reasonably well with the hydrophilic ABTS values (r = 0.85). In Fig. 8, the hydrophilic DPPH values and hydrophilic ORAC values showed a significant correlation (r = 0.87). However, antioxidant activity with the lipophilic extracts correlated poorly when compared in all methods studied.
The antioxidant properties depend on several structural features of the molecule of polyphenols in its base structure and are primarily attributed to the high reactivity of hydroxyl substituents. The B-ring hydroxyl configuration is the most significant determinant of scavenging of ROS. 30, 40-di OH (catechol) structure in the B-ring strongly enhances lipid peroxidation inhibition. A free 3-hy-droxyl group and 30, 40-catechol (dihydroxy) structure, a C2-C3 double bond and a 4-oxo group on the C ring endow the flavonoid with potent antioxidant function. The superiority of quercetin in inhibit- ing both metal and nonmetal-induced oxidative damage is partially ascribed to its free 3-OH substituent which is thought to increase the stability of the flavonoid radical (Davies, 2000). Fenton-induced oxidation is strongly inhibited by flavonoids with catechol, 4-oxo and 50-OH arrangements (Cao et al., 1997; Cheng and Breen, 2000; Yoshimoto et al., 1999a; Yoshimoto, 2001; Islam, 2006, 2008a, 2009) which is superior to isoforms that lack these features. Future research should include the use of in vivo techniques such as molecular marker analysis. Additional research on polyphenolic should include a standardization of methods for quantification, evaluation of physiological activities and bioavailability. The interactions of the various polyphenolics with pharmaceutical should also be emphasized in future studies. The results may help in future conventional and chemical breeding program for specific constituents to enhance or reduce the bioactive phytochemicals and for improvement of the desired quality criteria of orange fleshed sweetpotatoes as well as other produces.
ACKNOWLEDGMENT
The authors would like to thank Dr. Richard B. Walker, Professor of Biochemistry, University of Arkansas at Pine Bluff for his help and valuable comments. The author also thankful to Mr. Z. Adam and L. Anderson for their assistance during the period of this study. This work has been funded by United State Department of Agriculture.
REFERENCES
- Ames, B.N., M.K. Shigenaga and T.M. Hagen, 1993. Oxidants, antioxidants and the degenerative diseases of aging. Proc. Nat. Acad. Sci. USA, 90: 7915-7922.
CrossRefDirect Link - Awika, J.M., L.W. Rooney, X. Wu, R.L. Prior and L. Cisneros-Zevallos, 2003. Screening methods to measure antioxidant activity of sorghum (Sorghum bicolor) and sorghum products. J. Agric. Food Chem., 51: 6657-6662.
CrossRefDirect Link - Bao, J., Y. Cai, M. Sun, G. Wang and H. Corke, 2005. Anthocyanins, flavonols, and free radical scavenging activity of Chinese bayberry (Myrica rubra) extracts and their color properties and stability. J. Agric. Food Chem., 53: 2327-2332.
CrossRefDirect Link - Brand-Williams, W., M.E. Cuvelier and C. Berset, 1995. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol., 28: 25-30.
CrossRefDirect Link - Buechter, D.D., 1988. Free radicals and oxygen toxicity Pharmacol. Res., 525: 253-260.
PubMedDirect Link - Cao, G., E. Sofic and R.L. Prior, 1997. Antioxidant and prooxidant behavior of flavonoids: Structure-activity relationships. Free Radical Biol. Med., 22: 749-760.
CrossRefDirect Link - Cheng, I.F. and K. Breen, 2000. On the ability of four flavonoids, baicilein, luteolin, naringenin and quercetin to suppress the fenton reaction of the iron-ATP complex. Biometals, 13: 77-83.
PubMedDirect Link - Gorinstein, S., O. Martin-Belloso, E. Katrich, A. Lojek and M. Ciz et al., 2003. Comparison of the contents of the main biochemical compounds and the antioxidant activity of some Spanish olive oils as determined by four different radical scavenging tests. J. Nutr. Biochem., 14: 154-159.
PubMedDirect Link - Hagerman, A.E., K.M. Riedl, G.A. Jones, K.N. Sovik, N.T. Ritchard, P.W. Hartzfeld and T.L. Riechel, 1998. High molecular weight plant polyphenolics (Tannins) as biological antioxidants. J. Agric. Food Chem., 46: 1887-1892.
CrossRefDirect Link - Huang, D., B. Ou, M. Hampsch-Woodill, J.A. Flanagan and E.K. Deemer, 2002. Development and validation of oxygen radical absorbance capacity assay for lipophillic antioxidants using randomly methylated beta-cyclodextrin as the solubility enhancer. J. Agric. Food Chem., 50: 1815-1821.
CrossRefDirect Link - Huang, D., B. Ou and R.L. Prior, 2005. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem., 53: 1841-1856.
CrossRefPubMedDirect Link - Islam, S., M. Jalaluddin and N. Hettiarachchy, 2011. Bio-active compounds of bitter melon genotypes (Momordica charantia L.) in relation to their physiological functions. Functional Foods Health Dis., 2: 61-74.
Direct Link - Islam, S., 2008. Antimicrobial activities of Ipomoea batatas (L.) Leaf. J. Food Agric. Environ., 6: 14-17.
Direct Link - Islam, S., 2006. Sweetpotato (Ipomoea batatas L.) Leaf: Its potential effect on human health and nutrition. J. Food Sci., 71: R13-R21.
CrossRefDirect Link - Islam, S., Y. Yoshimoto, O. Yamakawa, 2003. Distribution and physiological function of caffeoylquinic acid derivatives in Leaves of sweetpotato genotypes. J. Food Sci., 68: 111-116.
CrossRef - Islam, M.S., M. Yoshimoto, K. Ishigure, S. Okuno and O. Yamakawa, 2003. Effect of artificial shading and temperature on radical scavenging activity and polyphenolic composition in sweetpotato (Ipomoea batatas L.) leaves. J. Am. Soc. Horticul. Sci., 128: 182-187.
Direct Link - Islam, M.S., M. Yoshimoto, N. Terahara and O. Yamakawa, 2002. Anthocyanin compositions in sweetpotato (lpomoea batatas L.) leaves. Biosci. Biotechnol. Biochem., 66: 2483-2486.
PubMed - Islam, S., M. Yoshimoto, S. Yahara, S. Okuno, K. Ishiguro and O. Yamakawa, 2002. Identification and characterization of foliar polyphenolic composition in sweetpotato (Ipomoea batatas L.) genotypes. J. Agric. Food Chem., 50: 3718-3722.
CrossRefDirect Link - Kaul, A. and K.L. Khanduja, 1998. Polyphenols inhibit promotional phase of tumorigenesis: Relevance of superoxide radicals. Nutr. Cancer, 32: 81-85.
Direct Link - Kusano, S., H. Abe and H. Tamura, 2001. Isolation of antidiabetic components from white-skinned sweet potato (Ipomoea batatas L.). Biosci. Biotechnol. Biochem., 65: 109-114.
PubMed - Matsui, T., S. Ebuchi, M. Kobayashi, K. Fukui, K. Sugita, N. Terahara and K. Matsumoto, 2002. Anti-hyperglycemic effect of diacylated anthocyanin derived from Ipomoea batatas cultivar ayamurasaki can be achieved through the α-glucosidase inhibitory action. J. Agric. Food Chem., 50: 7244-7248.
CrossRefPubMedDirect Link - Murayama, T., H. Yada, M. Kobori, H. Shinmoto and T. Tsushida, 2002. Evaluation of three antioxidants and their identification and radical scavenging activities in edible chrysanthemums (Dendranthema). J. Jpn. Soc. Hortic. Sci., 71: 236-242.
Direct Link - Ou, B., M. Hampsch-Woodill, J. Flanagan, E.K. Deemer, R.L. Prior and D. Huang, 2002. Novel fluorometric assay for hydroxyl radical prevention capacity using fluorescein as the probe. J. Agric. Food Chem., 50: 2772-2777.
PubMed - Ou, B., D. Huang, M. Hampsch-Woodill, J.A. Flanagan and E.K. Deemer, 2002. Analysis of antioxidant activities of common vegetables employing Oxygen Radical Absorbance Capacity (ORAC) and ferric reducing antioxidant power (FRAP) assays: A comparative study. J. Agric. Food Chem., 50: 3122-3128.
CrossRefDirect Link - Pellegrini, N., M. Serafini, B. Colombi, D. Del Rio and S. Salvatore et al., 2003. Total antioxidant capacity of plant foods, beverages and oils consumed in Italy assessed by three different in vitro assays. J. Nutr., 133: 2812-2819.
PubMedDirect Link - Prior, R.L., G. Cao, A. Martin, E. Sofic and J. McEwen et al., 1998. Antioxidant capacity as influenced by total phenolic and anthocyanin content, maturity and variety of Vaccinium species. J. Agric. Food Chem., 46: 2686-2693.
CrossRefDirect Link - Prior, R.L., 2003. Fruits and vegetables in the prevention of cellular oxidative damage. Am. J. Clin. Nut., 78: 570S-578S.
Direct Link - Prior, R.L. and G. Cao, 1999. In vivo total antioxidant capacity: Comparison of different analytical methods. Free Radic. Biol. Med., 27: 1173-1181.
CrossRefPubMedDirect Link - Prior, R.L., H. Hoang, L. Gu and X. Wu and M. Bacchiocca et al., 2003. Assays for hydrophilic and lipophilic Antioxidant capacity (oxygen radical absorbance capacity (ORACFL) of plasma and other biological and food samples. J. Agric. Food Chem., 51: 3273-3279.
CrossRefDirect Link - Singleton, V.L., R. Orthofer and R.M. Lamuela-Raventos, 1999. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. In: Methods in Enzymology, Burslem, G.L. (Ed.), Academic Press, Cambridge, Massachusetts, ISBN: 9780121822002, pp: 152-178.
CrossRefDirect Link - Robards, K., P.D. Prenzeler, G. Tucker, P. Swatsitang and W. Glover, 1999. Phenolic compounds and their role in oxidative processes in fruits. Food Chem., 66: 401-436.
CrossRefDirect Link - Stevens, K.L., R.E. Wilson and M. Friedman, 1995. Inactivation of a tetrachloroimide mutagen from simulated processing water. J. Agric. Food Chem., 43: 2424-2427.
CrossRefDirect Link - Teow, C.C., V.D. Truong, R.F. McFeeters, R.L. Thompson, K.V. Pecota and G.C. Yencho, 2007. Antioxidant activities, phenolic and β-carotene contents of sweet potato genotypes with varying flesh colours. Food Chem., 103: 829-838.
CrossRefDirect Link - Walker, R. and J.D. Everette, 2009. Camparative reaction rates of various antioxidants with the ABTS radical cation. J. Agric. Food Chem., 57: 1156-1161.
PubMedDirect Link - Waris, G. and H. Ahsan, 2006. Reactive oxygen species: Role in the development of cancer and various chronic conditions. J. Carcinog., 5: 14-21.
CrossRef - Yang, J. and R.L. Gadi, 2008. Effects of steaming and dehydration on anthocyanins, antioxidant activity, total phenols and color characteristics of purple-fleshed sweet potatoes (Ipomoea batatas). Am. J. Food Technol., 3: 224-234.
CrossRefDirect Link - Yoshimoto, M., S. Okuno, M. Yoshinaga, O. Yamakawa, M. Yamaguchi and J. Yamada, 1999. Antimutagenicity of sweetpotato (Ipomoea batatas) roots. Biosci. Biotechnol. Biochem., 63: 537-541.
PubMedDirect Link - Yoshimoto, M., S. Okuno, T. Kumagai, M. Yoshinaga and O. Yamakawa, 1999. Distribution of antimutagenic components in colored sweetpotatoes. Jap. Agric. Res. Q., 33: 143-148.
Direct Link - Yoshimoto, M., S. Okuno, M. Yamaguchi and O. Yamakawa, 2001. Antimutagenicity of deacylated anthocyanins in purple-fleshed sweetpotato. Biosci. Biotechnol. Biochem., 65: 1652-1655.
CrossRefPubMedDirect Link - Yoshinaga, M., O. Yamakawa and M. Nakatani, 1999. Genotypic diversity of anthocyanin content and composition in purple-fleshed sweetpotato (Ipomoea batatas (L.) Lam). Breed Sci., 49: 43-47.
Direct Link - Islam, I., A.U. Shaikh and I.M. Shahidul, 2009. Antioxidative and antimutagenic potentials of phytochemicals from Ipomoea batatas (L.) Lam. Int. J. Cancer Res., 5: 83-94.
CrossRefDirect Link