
Abdoulaye Coulibaly
State Key Laboratory of Food Science and Technology, Jiangnan University, Wuxi, Jiangsu 214122, People`s Republic of China
Jie Chen
State Key Laboratory of Food Science and Technology, Jiangnan University, Wuxi, Jiangsu 214122, People`s Republic of China
American Journal of Food Technology
Year: 2011 | Volume: 6 | Issue: 1 | Page No.: 40-51







ABSTRACT
This study was conducted to asses the energetic compounds (protein, fat and carbohydrate), some vitamins, minerals, antioxidant capacity, phytase and amylase activity during the germination of foxtail millet. Germination has been found to increase nutritive qualities of foxtail millet. One day soaking and germination up to 8 days increased significantly soluble sugars (reducing and total sugars), amylase activity from the first day to third day and phytase activity from first day to seventh day. The proteins were not changing during germination. Vitamins and minerals increased, however fat content and total phenolic content decreased through germination. Germination of foxtail millet for three days allowed obtaining flour with high amylolytic activity, high DPPH scavenging activity and high concentration of minerals, thus it can be added to different flour to initiate starch degradation and reduce viscosity, to control the minerals balance in children diet.
PDF Abstract XML References Citation
How to cite this article
Abdoulaye Coulibaly and Jie Chen, 2011. Evolution of Energetic Compounds, Antioxidant Capacity, Some Vitamins and Minerals, Phytase and Amylase Activity During the Germination of Foxtail Millet. American Journal of Food Technology, 6: 40-51.
DOI: 10.3923/ajft.2011.40.51
URL: https://scialert.net/abstract/?doi=ajft.2011.40.51
DOI: 10.3923/ajft.2011.40.51
URL: https://scialert.net/abstract/?doi=ajft.2011.40.51
INTRODUCTION
Germination is the set of processes by which a seed grows and gives birth to a new plant; it is the resumption of development and metabolism (absorption of water (imbibition), respiration, enzyme activity) of an embryo of seed plants.
It actually comprises 3 steps:
| • | A phase-dipping (imbibition) which will allow the seed to absorb the water they need and disable the enzyme inhibitors which prevent germination to occur |
| • | The second step is germination itself, it is irreversible |
| • | The last step also irreversible, in turn leads to a decrease in reserves of the seed and the exaltation of a seedling (Heller, 1995) |
In fact, the embryo uses the reserves of the seed to begin its development. The reserves consist of seeds of carbohydrates (mainly starch), lipids (fats) and proteins in variable proportions. Thus, the starchy grains like cereals contain mainly starch. The consumption of seed reserves by the embryo will provide energy and raw materials to train the young seedlings.
The nutrients are reduced to smaller units such as carbohydrates into simple sugars, proteins into amino acids, facilitating their uptake by our bodies.
In addition, it should be noted that these grains contain minerals and vitamins. Their quantities therein are not only insufficient to cover our needs, but also their bioavailability is low. Indeed, anti-nutrients like phytates are naturally present in cereal grains. They chelate minerals such as iron, zinc, calcium and therefore limit their absorption and utilization by the body (Gibson et al., 1998).
During germination, many enzymatic activities have been identified, among other amylolytic activity with the production of maltose, maltotriose and dextrin from starch hydrolysis (Weil, 1990), phytase activity. Phytases are responsible in plants from the release of inorganic phosphorus during the germination of seeds (reserve of phosphorus in seeds are in the form of phytate) (Hubel et al., 2001) and the production of free myo-inositol (non phosphate) is an important growth factor. Hydrolysis of phytate also releases cations (minerals) that could be linked to phytate chelating due to its negative charge.
Indeed malnutrition in infants, young children is the most serious health problems in developing world. The largest share of malnutrition in the world is made micronutrient deficiencies, which includes all minerals and vitamins (and some to add essential fatty acids). Malnutrition has been called the hidden hunger by the united state, affecting two billion people suffer from deficiencies in minerals and vitamins, can cause fatal diseases. In 2006, more than 3.5 billion people suffer from iron deficiency; 200 million preschool children suffer from vitamin A deficiency. In developing countries, children endure damaging growth delay to their physical and intellectual development. Adults are also affected by this problem, especially women. Anemia caused by iron deficiency causes the death of 20% of young women during pregnancy or childbirth, lack of folic acid causes one death by heart disease in ten.
Cereal grains have phenolic compounds which are located mainly in the pericarp. Phenolics are compounds possessing one or more aromatic rings with one or more hydroxyl groups, also these compounds are known as antioxidants which have long been recognized to have protective function against oxidative damage and in diet they may provide health benefits associated with reduced risk of chronic disease (Abdel-Aal et al., 2006; Liu, 2007; Huet al., 2003).
Generally, cereal and cereal products are the main complementary food in most developing countries. Germination has a profound change on nutritional quality of the cereal (Chavan and Kadam, 1989). Protein bioavailability is increased.
Since, the consumption of the cereals based flour food is very popular in the world, there is necessary to study traditional method (soaking and germination) of improving the nutritive value of cereal-gruels.
In this study, we would like to evaluate the effect of germination on the energetic compounds (carbohydrates, fat and proteins), some minerals and vitamins, the antioxidant activity, phytase and amylase activity in the foxtail millet.
MATERIALS AND METHODS
Foxtail millet (Setaria italica or Panicum italicum L.) was obtained from the local market (Wuxi, China) in June 2009.
Sample preparation: The seeds of millet were cleaned manually to remove broken seeds, dust and other extraneous materials. One kilogram was soaked in distilled water in ratio of 1:3 (p/v) for 24 h and then the grains were spread on a white cotton fabric and kept wet by spraying of distilled water (Fig. 1). After germination, the seeds were dried separately at 45°C to constant weight and were grounded.
Chemical analysis: Total nitrogen was estimated using Kjedahl’s method and 5.83 was used as factor to obtain millet protein value; fat content was extracted with Buchi extraction systems B-811 using petroleum ether 30-60°C according to AOAC methods (AOAC, 1995) and the carbohydrate content of each sample was determined by difference (%carbohydrate = 100-(%Moisture + %Protein + %Fat + %Ash)). pH was determined as described by Vasconcelos et al. (1990). The total titratable acidity was determined as described by Kirk and Ronald (1991).
Minerals determination: Hydrochloric acid extractability of minerals: Minerals in the samples were extracted by Chauhan and Mahjan (1988). About 1 g sample was shaken with 10 mL of 0.03 M HCl for 3 h at 37°C and then filtered. The clear extract obtained was oven-dried at 100°C and then the sample was acid-digested with diacid mixture (HNO3:HClO4, 5:1, v/v). The amount of extractable minerals was determined using atomic absorption spectroscopy.
Extraction of soluble sugar
Extraction of soluble sugar in the millet flour: One gram of sample was homogenized with 10 mL of 80% ethanol solution. The mixture was centrifuged at 5000 g for 10 min. The supernatant was collected and 2 mL of lead acetate 10% were added. The insoluble residue was removed by centrifugation at 5000 g for 10 min. Two milliliter of oxalic acid 10% were added in the new supernatant and then recentrifuged under the same conditions as before. The supernatant was collected and dried under a stream of hot air and residue was resuspended in 5 mL of water.
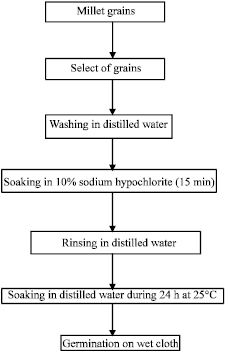 |
| Fig. 1: | Flow chart illustrating preparation of germinated millet |
Reduction sugars were estimated by the Dinitrosalisylic acid (DNS) method of (Bernfeld, 1955). Total sugars were carried out according phenol-sulfuric acid method (Dubois et al., 1956).
Analysis of vitamins by HPLC in millet flour
Vitamin E and A: Extraction of vitamin E and A in the millet flour: According to Mendoza et al. (2003), 1 g of sample was dissolved in 4 mL of ethanol. The mixture was stirred for 2 min and 400 μL of n-hexane was added. The homogenate was centrifuged at 2500 g for 5 min. The clear organic top layer was removed and filtered through a filter paper and then injection into HPLC.
Vitamins B1, B2 and B6: Vitamins contents were determined according to the methods described by Bergaentzle et al. (2000) with slight modification. Five grams of the sample were weighted; 50 mL of 0.05 M sodium acetate pH 4.5 were added to the sample in 250 mL conical flask, followed by enzyme mixture (10 mg of α-amylase, 100 mg of papain and 20 mg of phosphatase acid) was added. The solution was incubated for 18 h in a water batch at 37°C, then diluted to 100 mL with distilled water in a volumetric flask and filtered through a filter paper. The clear filtrate obtained was used for the chromatographic determination.
Vitamin C: Vitamin C was assessed According to Fontannaz et al. (2006), 10 g of sample were weighted into 100 mL volumetric flask, then 40 mL of TCEP-HCl (Tris 2-carboxyethl-phosphine hydrochloride) solution at 250 μg mL-1 and 10 mg of α-amylase were added and the suspension was incubated at 42°C for 30 min before filling the volumetric flask to the mark with TCA (Trichloroacetic acid) solution 1%. The resulting solution was shaking for about 1 min and filtered through a filter paper. Aliquots of the solution were diluted with the mobile phase prior the injection on to HPLC system.
Enzymatic extract
Preparation of enzyme extract: Ten gram of seeds millet were degermed and grounded with 20 mL of sodium chloride 0.9% at 4°C. The homogenate was centrifuged at 5000 g for 30 min at 4°C and the supernatant containing enzyme was used for assays.
The amylolytic activity was determined by the following way:
The amylolytic activity of crude enzyme extracts was measured by colorimetric after a dosage of reducing sugars released by action on the soluble starch (1%).
Three hundred seventy five microliter of acetate buffer (100 mM, pH 5) into test tube, 25 μL of enzymatic extract was added and followed of 100 μL of starch flour (1%). The mixture was incubated in a water bath at 37°C for 30 min. Reducing sugars released were assayed by the method of Bernfeld (1955) using the DNS. The reaction was stopped by adding 300 μL of DNS. The mixture was heated to the boiling water bath for 5 min and then cooled for 10 min at room temperature. Two millilitres of distilled water are added. The absorbance was read at 540 nm against a control containing no enzyme extract. This value was then converted into mg of reducing sugars through a calibration curve obtained under the same conditions from a solution of maltose to 2 mg mL-1. The specific activity was expressed in mg of reducing sugar/min/mg of protein.
The phytase activity was determined by following way:
Sodium phytate (5 mM) was the substrat used. One hundred fifty microliter of acetate buffer (100 mM, pH 5) was added to 25 μL of enzymatic extract into test tube and then 75 μL of sodium phytate (5 mM) was added. The mixture was incubated in a water bath for 4 h at 37°C. Inorganic phosphate (Pi) released was determined by the method of Makower (1970). The reaction was stopped by adding 1 mL of 2.5% TCA (w/v). We added 3 mL of molybdate vanadate and the test tube was left to stand at room temperature for 10 min. The absorbance was read at 410 nm against the control containing no enzyme extract. The value obtained was converted into mg of inorganic phosphate through a calibration curve obtained from a solution of KH2PO4 (5 mM). The specific activity of phytase is expressed as mg Pi/h/mg protein.
Assessment of antioxidant activity
Extraction: Two grams of each sample was extracted with 25 mL of 60% methanol containing 1% HCl for 4 h at room temperature by shaking vigorously. The mixture was centrifuged at 4000 g for 15 min at 4°C and the supernatants filtered through filter paper and the filtrate stored at 4°C still analysis.
Total phenolic content determination: Total Phenolic Content (TPC) was determined by Folin-Ciocalteu colorimetric method with slight modification (Bao et al., 2005; Cai et al., 2004). The 0.5 mL of the sample extract with equal volume of methanolic solution and 0.5 mL of Folin-Ciocalteu reagent were mixed and then 1 mL of 75 g L-1 sodium carbonate was added. The mixture was incubated at room temperature for 2 h and the absorbance was measured at 760 nm using a spectrophotometer. A calibration curve was prepared using gallic acid solution. TPC were expressed as milligrams of gallic acid equivalent (GAE) per g of dry weight (mg GAE/g of dw).
DPPH free radical scavenging activity: Free radical scavenging activity was determined according to the method of Gaulejac et al. (1998) with slight changes. Briefly, 0.1 mL aliquot of sample extract was mixed with 2.9 mL of 6.10-5 mol L-1 methanolic solution of DPPH and stirred vigorously. The absorbance at 517 nm was measured after the solution had been allowed to stand in the dark for 60 min. Lower absorbance of the reaction mixture indicates higher free radical scavenging activity. The final results were expressed as micromoles of Trolox Equivalents (TE) per gram of dry weight (μmol TE g-1 of dw).
Data analysis: All analysis were carried out in triplicate and the data were reported as Means±SD. Analysis of variance and significant differences among means were tested by one-way ANOVA using SAS 2003.
RESULTS AND DISCUSSION
The fat and protein content of foxtail millet during germination are presented in Table 1.
The total protein content of foxtail millet is 11.9%, however the result is included in the interval given by Zheng-li et al. (2006) and they found that the crude protein content of all foxtail millet varieties in China is in the range of 9.3-12.71%. During germination, the protein content was not significance at 5% level. There isn’t real change in the amount of protein because of the germination system.
Fat content in raw foxtail millet was 4.095% (Table 1). Germination reduced significantly (p<0.05) fat content of the foxtail millet with time. The earlier days (1 and 2 day), the fat content did not be different greatly.
The decrease of fat during germination may be due to the action of lipolytic enzymes which used the fat present. This could be owing to fat catabolism for energy production contributing to seed growth. This reduce brings an increase in palatability of millet food products. It reduced the possibility of development of free fatty acids, which occur mainly due to the action of lipase, causing bitterness and can make millet meals unacceptable within a few days. The same study was reported by Nnam (2000), Inyang and Idoko (2006) and Inyang and Zakari (2008).
During germination of foxtail millet, several significant (p<0.05) changes occurred in the carbohydrates profile. A significant increase (p<0.05) in total soluble sugar was observed, the amount of total soluble sugar increased drastically 0.995% (raw millet) to 12.995% (6 days), after these days the amount of total soluble sugar was insignificantly change (Table 2). In germination, the reducing sugar increased significantly (p<0.05) to reach its maximum on the third day and thereafter a slight decrease was observed (Table 2). The total carbohydrate decreased also considerably in the foxtail millet during germination. These changes may be caused by degradation of starch content. The major enzyme involved in starch degradation is amylase. So, the total amylase activity changes significantly (p<0.05) during germination (Table 3). The results show a rise in amylase activity in foxtail millet and the level of amylase activity achieved its maximum (0.0512 μg reducing sugar/min/mg protein) on the third day and started to decrease until at the end of process as reducing sugar. The increase in soluble sugar (reducing and total soluble sugars) and amylase action during germination of foxtail millet might be due to carbohydrate metabolism, it may be due to enhanced amylase action which results in the degradation of starch into simpler and more absorbable compounds which were utilized by embryo for its future development.
| Table 1: | Changes in fat and protein during germination of foxtail millet |
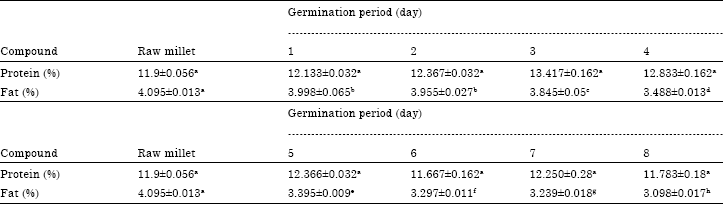 | |
Values represent means of triplicate. Values with the same letter are not significantly different (p<0.05) | |
| Table 2: | Changes in carbohydrate profile during germination of foxtail millet |
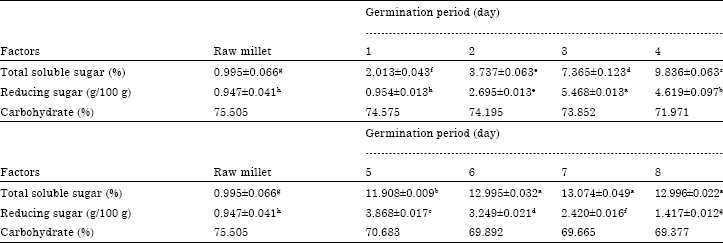 | |
Values represent means of triplicate. Values with the same letter are not significantly different (p<0.05) | |
During germination there is a reduction in total carbohydrate content of the foxtail millet and increase in the percentage of the total soluble sugar (Table 2). This can be attributed to the fact that some of the endosperm starch is consumed during germination to provide energy. These results are consistent with the finding of Prado et al. (2000), Demuyakor and Ohta (1992) and Lasekan (1996). The increase in total soluble sugar might be due to the synthesis of new polysaccharides for the walls. Similar results had been found by Jood et al. (1998) in chickpea grain.
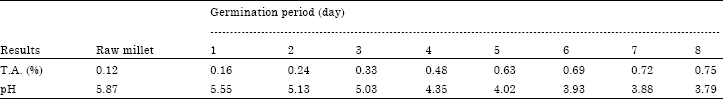
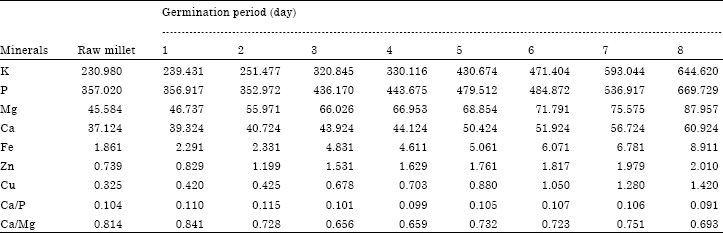
Table 4 presents pH and titratable acidity in the foxtail millet during germination. The pH in raw foxtail millet is 5.87 and 0.12% for titratable acidity. During germination titratable acidity increased. This could be due to the production of organic acids such as lactic acid, citric acid and acetic acid during the degradation of carbohydrate to provide energy for the growth of seedlings. So, the decrease in pH of foxtail millet flour may be due to higher total titratable acidity. The pH values from first to fourth day of germination are within the range of optimum pH of amylase, which could explain the increase of amylolytic activity from first to third day of germination. Nirmala and Muralikrishna (2003) have shown that, amylases extracted from finger millet were completely inactive at pH below 4 and their optimum pH was between 4.5 and 5.5. Table 5 presents the HCl extractability of minerals during germination of foxtail millet. The K, P, Mg, Ca, Fe, Zn and Cu were respectively in the foxtail millet 230.98, 357.02, 45.584, 37.124, 1.861, 0.739, 0.325 mg/100 g. The HCl extractability for all minerals increased, germination improve the minerals content of foxtail millet. The result was consistent with (Nnam, 2000). The phosphorus content decreased at the beginning of germination and started to increase until at the end of process. The decrease in level of phosphorus during the two firsts days of germination may be attributed to soaking. From the second day of germination we observed an increase in phosphorus. This may be due to hydrolysis of phytic acid by phytase. Parallel to HCL extractability of phosphorus, Table 6 presents the change in phytase activity during germination of foxtail millet.
| Table 3: | Total amylase (μg Reducing sugar/min/mg protein) and Phytase activity (μg Pi/h/mg protein) during germination of foxtail millet |
 | |
Values represent means of triplicate. Values with the same letter are not significantly different (p<0.05) | |
| Table 4: | Changes in pH and titratable acidity (T.A) during germination of foxtail millet |
 | |
| Table 5: | Hydrochloric acid extractability (mg/100 g) of minerals in foxtail millet during germination |
 | |
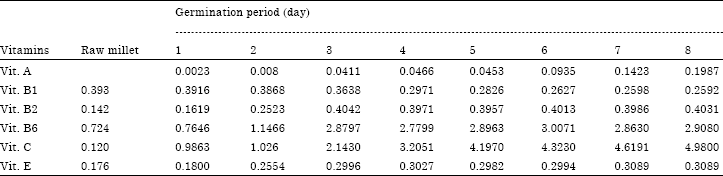
| Table 6: | Changes in some vitamins during germination of foxtail millet (mg/100 g) |
 | |
Phytase activity increased significantly (p<0.05) with increase in germination period. The level of phytase activity reached a maximum at 7 days of germination, this maximum value of phytase activity is 70.023 μg Pi/h/mg of proteins. Larsson and Sandberg (1992) observed phytase activity during germination of wheat, barley, rye and oats; they noted that, this enzyme hydrolysing phytic acid to phosphate and myo-inositol phosphates. The increase in HCI extractable minerals may be attributed to reduction in phytate and presence of enhancers such as organic acids and ascorbic acid, similar result have been mentioned by Indumadhavi and Agte (1992). During germination, the Ca/P and Ca/Mg value are between 0.091 to 0.115 mg/100 g and 0.656 to 0.841 mg/100 g, respectively. The recommended value of Ca/P and Ca/Mg are 1.0 and 2.2 mg/100 g, respectively (NRC, 1989). Ca/P and Ca/Mg value during germination are very low compared with the recommended value; this could be due to the low value of calcium comparatively with phosphorus and magnesium. These three minerals are important in teeth and bone formation and the control of calcium in the blood. From this analysis it appears to supplement products made from millet with calcium could prevent deficiency diseases.
Table 6 shows some vitamins content in the foxtail millet during germination. In the raw foxtail millet we note the absence of vitamin A, 0.393, 0.142, 0.724, 0.120 and 0.176 mg/100 g are respectively vitamins A, B1, B2, B6, C and E content. During germination, the slight decrease was observed for vitamin B1, for other vitamins we observed an increase. Bibi et al. (2008) also observed that water-soluble vitamins such as B complex and vitamin C are synthesized. The increase in vitamin A could be explained by the conversion of β-carotene into vitamin A during germination. The similar results were observed by Chavan (1989) who had reported the decrease in thiamine (vitamin B1) content and the increase in riboflavin (vitamin B2), niacin and vitamin C content in malted cereals. Increasing the quantity of vitamins could be due to the fact that during germination, seeds synthesized vitamins for its development.
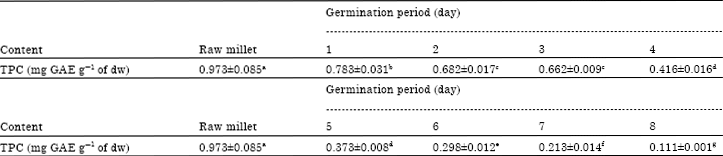
| Table 7: | Total phenolic content (mg of gallic acid equivalent/g dry weight) during germination of foxtail millet |
 | |
Values represent means of triplicate. Values with the same letter are not significantly different (p<0.05) | |
| Table 8: | DPPH radical scavenging activity (μmol of trolox equivalents/g dry weight) during germination of foxtail millet |
 | |
Values represent means of triplicate. Values with the same letter are not significantly different (p<0.05) | |
Table 7 presents the evolution of Total Phenolic Content (TPC) in the foxtail millet during germination. In the raw seed, TPC is 0.973 mg GAE/g of dry weight and then, we observed the decrease of TPC from first day to end of germination process. The results showed that germination had significantly (p<0.05) reduced total phenolic content. To germinate, the seed in addition to starch, amino acids and fat, also uses its coat; this could explain the decrease of TPC. According to the germination process the seeds have been soaked for 24 h, so the leaching during soaking could promote also the decrease of total phenolic content. During germination the decrease of TPC could be due to hydrolysis of total phenolic compounds, similar results had been found by Maillard et al. (1996). This may also be due to rupture of ester bonds in the phenolic compounds by esterase that act on phenolic acid esters linked either to arabinoxylans or other non-starch polysaccharides. To eliminate the mousy odour in the foxtail millet meal the decrease of total phenolic content is necessary.
The antioxidant capacity during the germination of foxtail millet was measured using the 1, 1-diphenyl-2 picrylhydrazyl (DPPH) assay and the results are presented in the Table 8.
The free radical scavenging activity for the raw millet is 15,780 μmol TE g-1 of dry weight. DPPH radical scavenging activity decreased significantly (p<0.05) from first day to third day of germination, after the third day the increase of the DPPH radical scavenging was not significant. The decrease of DPPH radical scavenging could be due to soaking. According to the DPPH assay, the germinated millet for three days has a high free radical scavenging activity.
CONCLUSION
We observed that the minerals and vitamins content increased during germination; a strong enzymatic activity (amylase and phytase activity) and mobilisation of soluble sugar were also observed. The protein content in the foxtail millet did not change during germination; however the fat content decreased with germination period. Further, the results showed that the supplementation of millet flour with other sources of protein could overcome the lack of protein in cereal food and decreasing fat content allowed to increase the shelf life of millet flour by reducing the rancidity level. The technique of germination simple and requires few tools. The cost is negligible and food is fresh produced lacking chemical agents. Germination is simple process of transformation enlivens the food by releasing all enzymatic and nutritional power. Thus, germination increases tenfold potential nutritious. Vitamins, enzymes and minerals increase. It removed by soaking and rinsing, the enzyme inhibitors, chemical and natural pesticides, thus producing food easier to digest, more easily assimilated by the body, while requiring less work for our body.
ACKNOWLEDGMENTS
The germination process and the nutritional evaluation was done at the State Key Laboratory of Food Science and Technology in Southern Yangtze University. Financial support was provided by Key Laboratory of Food Science and Technology, Ministry of Education, Wuxi, China.
REFERENCES
- Abdel-Aal, E.M., J.C. Young and I. Rabalski, 2006. Anthocyanin composition in black, blue, pink, purple and red cereal grains. J. Agric. Food Chem., 54: 4696-4704.
CrossRefPubMedDirect Link - AOAC., 1995. Official Methods of Analysis. 16th Edn., Association of Official Analytical Chemists, Washington, DC., USA.
Direct Link - Bao, J., Y. Cai, M. Sun, G. Wang and H. Corke, 2005. Anthocyanins, flavonols, and free radical scavenging activity of Chinese bayberry (Myrica rubra) extracts and their color properties and stability. J. Agric. Food Chem., 53: 2327-2332.
CrossRefDirect Link - Bibi, N., Z. Aurang, B.K. Amal and S.K. Mohammad, 2008. Effect of germination time and type of illumination on proximate composition of chickpea seed (Cicer arietinum L.). Am. J. Food Technol., 3: 24-32.
CrossRefDirect Link - Cai, Y., Q. Luo, M. Sun and H. Corke, 2004. Antioxidant activity and phenolic compounds of 112 traditional Chinese medicinal plants associated with anticancer. Life Sci., 74: 2157-2184.
CrossRefPubMedDirect Link - Chavan, J.K., S.S. Kadam and L.R. Beuchat, 1989. Nutritional improvement of cereals by sprouting. Crit. Rev. Food Sci. Nutr., 28: 401-437.
CrossRefPubMedDirect Link - Chauhan, B.M. and H. Mahjan, 1988. Effect of natural fermentation on the extractability of minerals from pearl millet flour. J. Food Sci., 53: 1576-1577.
CrossRefDirect Link - DuBois, M., K.A. Gilles, J.K. Hamilton, P.A. Rebers and F. Smith, 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem., 28: 350-356.
CrossRefDirect Link - Demuyakor, B. and Y. Ohta, 1992. Malt characteristics of sorghum vulgares varietes from Ghana. J. Sci. Food Agric., 59: 457-462.
CrossRefDirect Link - Prado, F.E., C. Boero, M. Gallardo and J.A. Gonzalez, 2000. Effect of NaCl on germination, growth and soluble sugar content in Chenopodium quinoa willd. seeds. Bot. Bull. Acad. Sin., 41: 27-34.
Direct Link - Gibson, R.S., E.L. Fergurson and J. Lehrfeld, 1998. Complemntary foods for infant feeding in developing countries: Their nutrient adequacy and improvement. Eur. J. Clin. Nutr., 52: 764-770.
PubMedDirect Link - De Gaulejac, N.S.C., C. Provost and N. Vivas, 1999. Comparative study of polyphenol scavenging activities assessed by different methods. J. Agric. Food Chem., 47: 425-431.
CrossRefPubMedDirect Link - Hu, C., J. Zawistowski, W. Ling and D.D. Kitts, 2003. Black rice (Oryza sativa L. indica) pigmented fraction suppresses both reactive oxygen species and nitric oxide in chemical and biological model systems. J. Agric. Food Chem, 51: 5271-5277.
CrossRefPubMedDirect Link - Hatzack, F., F. Hubel, W. Zhang, P.E. Hansen and S.K. Rasmussen, 2001. Inositol phosphates from barley low-phytate grain mutants analysed by metal-dye detection HPLC and NMR. Biochem. J., 354: 473-480.
Direct Link - Indumadhavi, M. and V. Agte, 1992. Effect of fermentation on ionizable iron in cereal-pulse combinations. Int. J. Food.Sci. Technol., 27: 221-228.
CrossRef - Inyang, C.U. and C.A. Idoko, 2006. Assessment of the quality of ogi made from malted millet. Afr. J. Biotechnol., 5: 2334-2337.
Direct Link - Inyang, C.U. and U.M. Zakari, 2008. Effect of germination and fermentation of pearl millet on proximate, chemical and sensory properties of instant Fura: A Nigerian cereal food. Pak. J. Nutr., 7: 9-12.
CrossRefDirect Link - Jood, S., B.M. Chauhan and A.C. Kapoor, 1998. Contents and dgestibility of carbohydrates of chickpea and black gram as affected by domestic processing and cooking. Food Chem, 30: 113-127.
Direct Link - Larsson, M. and A.S. Sandberg, 1992. Phytate reduction in oats during malting. J. Food Sci., 57: 994-997.
CrossRefDirect Link - Zheng-li, L., S. Shi-xian, C. Ru-hong, H. Wen-sheng and L. Jun-xin et al., 2006. Development of a new foxtail millet germplasm with super early maturity and high iron content. Agric. Sci. China, 5: 558-562.
CrossRef - Makower, R.U., 1970. Extraction and determination of phytic acid in beans (Phaseolus vulgaris). Cereal Chem., 47: 288-295.
Direct Link - Maillard, M.N., M.H. Soum, P. Biovin and C. Berset, 1996. Antioxidant activity of barley and malt: Relationship with phenolic content. LWT-Food Sci. Technol., 29: 238-244.
CrossRefDirect Link - Nirmala, M. and G. Muralikrishna, 2003. In vitro digestibility studies of cereal flours and starches using purified finger millet (Eleusine coracana, ragi, Indaf-15) amylases. Carbohydrate Polym., 52: 275-283.
CrossRef - Fontannaz, P., T. Kilinc and O. Heudi, 2006. HPLC-UV determination of total vitamin C in a wide range of fortified food products. Food Chem., 94: 626-631.
CrossRef - Bergaentzle, S.N.D.M., D. Aoude-Werner and C. Hasselmann, 2000. Extraction procedures for the liquid chromatographic determination of thiamin, riboflavin and vitamin B6 in food stuffs. Food Chem., 71: 129-138.
CrossRef - Liu, R.H., 2007. Whole grain phytochemicals and health. J. Cereal Sci., 46: 207-219.
CrossRefDirect Link