
A.S.M. Golam Masum Akond
Division of Science and Mathematics, Mayville State University, ND, USA
Laila Khandaker
Division of Science and Mathematics, Mayville State University, ND, USA
Janelle Berthold
Division of Science and Mathematics, Mayville State University, ND, USA
Lori Gates
Division of Science and Mathematics, Mayville State University, ND, USA
Katelyn Peters
Division of Science and Mathematics, Mayville State University, ND, USA
Hardy Delong
Division of Science and Mathematics, Mayville State University, ND, USA
Khwaja Hossain
Division of Science and Mathematics, Mayville State University, ND, USA
American Journal of Food Technology
Year: 2011 | Volume: 6 | Issue: 5 | Page No.: 385-394







ABSTRACT
The anthocyanin, total polyphenol and antioxidant activity of 29 common bean from diverse origins and seed coat color, was assessed. Among the bean genotypes, fourteen were developed by CIAT in various interests; thirteen were from the USA, representing several market classes and one each from Brazil and India. The seeds of included genotypes have shown distinction in shape, color and seed weight. The variations of seed color are white, cream, purple, red and black, with variations being striped, rhomboid spotted and circular mottled. Bean genotypes exhibited distinguishing differences in anthocyanin, total polyphenol and antioxidant activities. Anthocyanin content varied significantly among genotypes and market classes, ranging from 0.05 to 0.47 mg g-1. The bean genotypes with total polyphenol content ranging from 5.87 to 14.14 mg of gallic acid equiv/g and the sample also exhibited significant variation in antioxidant activity (17.09 to 36.96%). Considering the profile of bioactive compounds the genotypes T-39, XAN 176, BAT 93 and MIB 154 are promising. Generally bean genotypes with high anthocyanin and polyphenol content exhibit high antioxidant activity. The information of this study can be used for selecting superior bean genotypes for targeted food and feed purposes and also for a breeding program.
PDF Abstract XML References Citation
Received: April 09, 2010;
Accepted: July 13, 2010;
Published: September 28, 2010
How to cite this article
A.S.M. Golam Masum Akond, Laila Khandaker, Janelle Berthold, Lori Gates, Katelyn Peters, Hardy Delong and Khwaja Hossain, 2011. Anthocyanin, Total Polyphenols and Antioxidant Activity of Common Bean. American Journal of Food Technology, 6: 385-394.
DOI: 10.3923/ajft.2011.385.394
URL: https://scialert.net/abstract/?doi=ajft.2011.385.394
DOI: 10.3923/ajft.2011.385.394
URL: https://scialert.net/abstract/?doi=ajft.2011.385.394
INTRODUCTION
Common beans (Phaseolus vulgaris) are important crop in the USA include market class Pinto, Navy, Kidney, Black Beans and many others. Each market class is defined by a specific seed size, color, pattern and traits controlled by many genes (McClean et al., 2002). International Center for Tropical Agriculture (CIAT) has bred a series of Andean bean genotypes with improved micronutrient content and reduced anti-nutritional factors and are useful for production zones in the tropics and subtropics (Blair et al., 2010). These are also distinguished by biochemical (Gepts and Bliss, 1986; McClean et al., 2004) and morphological (Gepts and Debouck, 1991) traits. Consumption of beans has been linked to reduced risk of diabetes and obesity (Geil and Anderson, 1994; Venkateswaran et al., 2002), coronary heart disease (Anderson et al., 1984; Bazzano et al., 2001), colon cancer (Hughes et al., 1997; Hangen and Bennink, 2002) and gastrointestinal disorders (Bourdon et al., 2001). Epidemiological Studies (Correa, 1981; Kolonel et al., 2000) confirm the highly significant inverse correlation between bean intake and age adjusted mortality for colon, breast and prostate cancers. Legume consumption (excluding soy foods) may have a protective effect against prostate cancer in humans according to a recent multiethnic case control study (Kolonel et al., 2000). This protective effect could reflect the possibility of a ubiquitous bioactive constituent akin to polyphenols, since it was not related to dietary fiber (Jain et al., 1999). Polyphenols in bean include tannins, anthocyanins and flavonols (Aparicio-Fernandez et al., 2005; Beninger and Hosfield, 2003). Presence of anthocyanins has only been reported in black and blue-violet colored beans (Aparicio-Fernandez et al., 2005; Romani et al., 2004). Polyphenols are known to exhibit strong antioxidant, antimutagenic and antigenotoxic activities and particular components such as flavonoids in Vigna sinensis and anthocyanins from black beans also prevent genetic damage induced by chemical mutagens in animal models (Wong et al., 2003; Azevedo et al., 2003), thereby providing evidence of anticancer activity.
Several findings on the biochemicals of bean have been reported. Some recent biochemical studies center on the profiles of seed coat anthocyanin glycosides (Choung et al., 2003) and polyphenols (Espinosa-Alonso et al., 2006). The work of Williams et al. (1995) focused on the flavonoid profiles in the leaves of 17 Phaseolus species (cultivated and wild) and in the flowers of nine species. Efforts have not yet been made to study the anthocyanin, total polyphenol and antioxidant profile of commonly grown USA and CIAT developed bean genotypes. This investigation describes determinants of naturally occurring variability in anthocyanin, polyphenols and antioxidant activity of 29 bean genotypes. Such information is critical in developing practical strategies to improve bean quality and enhance market opportunities for bean products in the functional food and nutraceutical industry.
MATERIALS AND METHODS
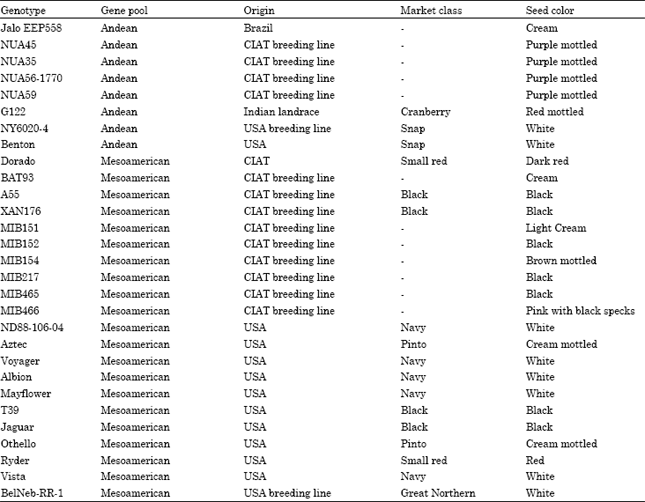
Plant material: Twenty-nine common bean genotypes were selected for the present study of which fourteen were from CIAT; thirteen were from USA and one each from Brazil and India. All common bean genotypes, along with their pedigree (if known), country of origin and market classes are listed in Table 1. Pot experiments were carried out at the greenhouse of the Mayville State University, ND, USA during the summer of 2008 in order to investigate the profile of anthocyanin, polyphenol and antioxidant activity.
Determination of total anthocyanin content: Total Anthocyanin was determined following the method described by Neff and Chory (1998) with minor modification. Three hundred microliter of Methanol with 1% HCl solution was added to 400 mg ground bean sample and mixed thoroughly; the extraction was then allowed to occur overnight in a dark refrigerator. Next day 200 mL Milli-Q H20 was added, followed by adding 500 μL of chloroform to each tube using a fume hood. Then the tubes were spun in a centrifuge set at the highest rpm for 2-5 min. The supernatant/aqueous (top) solution from the tube was transferred into a new 1.5 mL microfuge tube. The volume was brought up to 800 mL by adding 400 mL of a 60% Methanol 1% HCl solution. The absorbance of each tube was recorded using a spectrophotometer at 530 nm against a reagent blank with 480 μL Methanol 1% HCl and 320 mL Milli-Q H20 for a total solution of 800 μL. The optical density of the extracted solution was measured at 530 and 657 nm. Anthocyanin content was estimated by using the equation A530- (0.25).A657, which compensates for the contribution of Chlorophyll to the absorbance at 530 nm (Rabino and Mancinelli, 1986).
Determination of total polyphenol contents: Total phenolic content was determined using the method described by Khandaker et al. (2008) with Folin-Ciocalteu reagent according to the method of Slinkard and Singleton (1977) using gallic acid as a standard phenolic compound. Two milliliter of a 1:10 diluted sample (1 mg extract in 10 mL 70% ethanol) was placed in a test tube and mixed with 10 mL of Folin-Ciocalteu reagent (2 N, Sigma Chemical Co., St. Louis, MO, USA) previously diluted 1:10 with deionized water.
| Table 1: | Common bean genotypes, gene pool group, origin, seed color |
 | |
Between 1 min and 8 min, 8 mL of a sodium carbonate solution, prepared by dissolving 75 g in 1 L of deionized water (Sigma Chemical Co., St. Louis, MO, USA), was added to the test tube and mixed thoroughly with a Vortex mixer (Genie 2, Fisher Scientific, Bohemia, NY, USA) for about 5 sec. Then the test tubes with the mixtures were allowed to stand for 1 h in the dark. Absorbances of the resulting solutions were read at 760 nm using a spectrophotometer (Model 8451A, Diode Array Spectrophotometer, Hewlett Packard, CA, USA). Quantification of total phenolics was based on a gallic acid standard curve generated by preparing 0, 5, 10, 15, 20, 30 mg L-1 of gallic acid (Sigma Chemical Co., St. Louis, MO, USA) in deionized water. Total phenolics were expressed as mg gallic acid equivalent (GAE) per gram of bean seed using the following formula: Gallic acid equivalent (mg g-1 GAE) = ("x" Coefficient from the GA standard curve x Absorbance at 760 nm + Slope of the GA standard curve).x 10 (Dilution factor).
Antioxidant activity assay: Antioxidant activity was measured by the diphenylpicrylhydrazyl (DPPH) radical degradation method (Burits and Bucar, 2000) described by Khandaker et al. (2008). Briefly, 10 μL of leaf extract solution (three replicates) was introduced into test tubes and 4 mL distilled water and 1 mL of 250 μM DPPH solution was added. The tubes were mixed and allowed to stand for 30 min in the dark. Absorbance was read against a blank at 517 nm using a spectrophotometer. Antioxidant activity was calculated as the percent of inhibition relative to the control using the following equation: Antioxidant activity (%) = (Ablank-Asample/Ablank) x 100, where Ablank is the absorbance of the control reaction (control consisted of 10 μL, methanol instead of a sample extract) and Asample is the absorbance of the test compound.
Statistical analysis: Each determination was carried out on three separate replications, analyzed in triplicate and then the figures were averaged. Data was assessed by the Analysis Of Variance (ANOVA) following Tukey's multiple range tests and significance was accepted at p<0.05 (Tukey, 1953). The PC software ‘Excel Statistics’ (Version 5.0, Esumi Co. Ltd., Japan) was used for the calculations.
RESULTS AND DISCUSSION
Bean seeds of selected genotypes have shown distinction in shape, color and seed weight. The variations of seed color are white, cream, purple, red and black. Patterns of seed color vary between striped, rhomboid spotted and circular mottled. The weight of 100 beans ranges between 15.88-78.85 g (p<0.05) with a mean of 29.97 g (Table 2). The highest seed weight was recorded for Andean genotype NUA45 (78.85 g) and the lowest seed weight was recorded for Mesoamerican genotype MIB152 (19.27 g), both of these genotypes were developed by CIAT breeding program.
Significant differences were found in anthocyanin content among 29 bean genotypes and market classes (Table 2). The black colored Mesoamerican genotype T-39 contained the highest anthocyanin content (0.47 mg g-1), followed by the black colored CIAT genotype XAN (0.45 mg g-1). Cream mottled Pinto bean Orthello (0.05 mg g-1) has the lowest content followed by cream colored Brazilian genotype JaloEEP 558 (0.06 mg g-1), Great Northern white bean BelNEB (0.06 mg g-1) and cream colored CIAT genotype BAT 93 (0.06 mg g-1). Purple mottled four NUA series genotypes developed by CIAT contained medium range (0.20-0.33 mg g-1) of anthocyanin. Seven bean genotypes, T-39 (4.60 mg kg-1), XAN (4.56 mg kg-1), MIB154 (4.59 mg kg-1), G 122 (4.18 mg kg-1), BAT 93 (3.65 mg kg-1), DOR 364 (3.32 mg kg-1) and MIB 217 (3.16 mg kg-1) had higher seed anthocyanin than those of others. The anthocyanin content decreased in the following order based on average values of market classes in the USA grown bean genotype: Black > Small red > Snap > Navy > Pinto > Great Northern. Bean genotypes with dark colored (black, red, or purple) flesh have higher anthocyanin content than the white or yellow genotypes. Low concentrations of anthocyanins were found in the seeds of the white/yellow flesh genotypes because anthocyanins are a group of well-known water-soluble pigments, which contribute significantly to the red-blue coloration of plant materials. The included genotypes contained at the higher end of the anthocyanin spectrum compared to the studied red and black bean spectrum reported by other scientists (Horbowicz et al., 2008; Wu et al., 2006). Although, we did not determine the specific anthocyanin group in the 29 bean genotypes, several researchers found different anthocyanin groups in colored beans. For example, found in black beans, delphinidin-3-O-β-D-glucoside, petunidin-O-β-D-glucoside and malvidin-3-O-β-D-glucoside accounted for 56, 26 and 18%, respectively, of the anthocyanins and were responsible for the black seed color (Tsuda et al., 1994). The delphinidin-3-glucoside, petunidin-glucoside, malvidin-3, 5-diglucoside, malvidin-3-galactoside and malvidin-3-glucoside contents of 2.2, 0.8, 0.5, 0.03 and 0.4 mg g-1 of black beans were reported by Xu et al. (2007).
| Table 2: | Anthocyanin, Total Polyphenol (TP) and antioxidant activity in common bean genotypes |
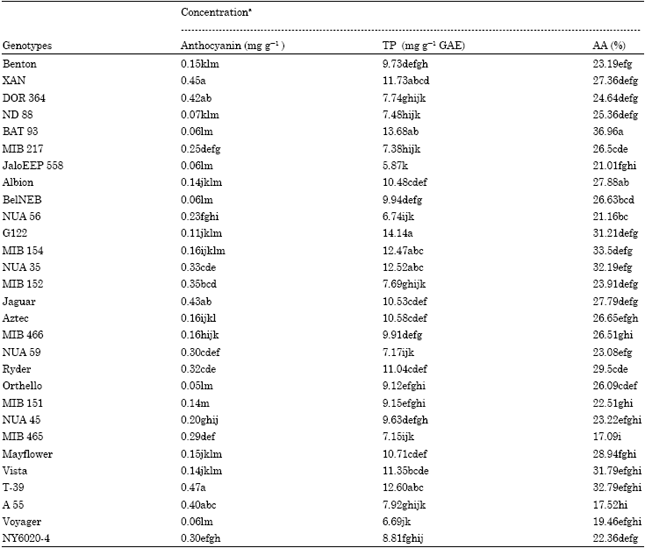 | |
aMeans in a column with different letters are significantly different (p<0.05) (n = 3). Concentrations of anthocyanin, total polyphenols are expressed as mg (+) cyanidin-3-glucoside, gallic acid, equivalents g-1 sample for anthocyanin and total polyphenol, respectively. Antioxidant activity is expressed as a percentage | |
Pelargonidin 3-glucoside was the major anthocyanin in red kidney beans (Choung et al., 2003) and is responsible for the red pigmentation of this type of kidney bean. Compared to the few data on anthocyanin in cereals and pseudocereals available in the literature, our anthocyanin values were higher than jasmine rice, Amaranthus hybridus, soybean (Gorinstein et al., 2007) and black sorghum (Awika et al., 2004). Quoted total anthocyanin values however cannot be compared directly with the results of other research group because of the inherent variability of botanical issues, sampling preparation and extraction procedures. Also content of anthocyanin may differ among different plant parts in same cultivars as the total anthocyanin content varies considerably; affected by genes, light, temperature and agronomic factors (Horbowicz et al., 2008).
Variation in total polyphenol content ranging from 5.87 (JaloEEP) to 14.14 (G122) mg g-1 GAE of sample was significant (p<0.05) among genotypes and market classes (Table 2). Seven bean genotypes, G 122 (14.14 mg kg-1), BAT 93 (13.68 mg kg-1), T-39 (12.60 mg kg-1), NUA 35 (12.52 mg kg-1), MIB 154 (12.47 mg kg-1), XAN (11.73 mg kg-1) and Vista (11.35 mg kg-1) had higher seed total polyphenol than those of others. Among the CIAT developed, genotype BAT 93 contained the highest (13.68 mg g-1 GAE), which is second highest among all. The CIAT developed MIB bean series contained a good amount of polyphenols except MIB 465 (7.15 mg g-1 GAE). The polyphenol content decreased in the following order based on average values of bean market classes in the USA grown bean genotype: Black > Small Red > Great Northern > Pinto > Navy > Snap, the same order was not observed previously in a study of 39 Canadian dry bean cultivars grown at four locations in Manitoba in 2003 (Balasubramanian et al., 2004). In general White Beans, Navy and Great Northern, contained significantly less polyphenol than colored beans in accordance with previous studies (Laparra et al., 2008). Recently Xu and Chang (2009) reported the phenolic acids in Pinto and Black beans varied depending on bean type and Pinto beans contained higher total phenolic acids than Black beans which were not consistent with our results. Aside from bean, other food commodities, such as sweet potatoes and sorghum, show a similar relationship between seed color and polyphenols. Brown and black sorghum had the highest levels of freely extractable polyphenols (Awika, 2003). Because of the differences in the methods of extraction and determination and in the ways of expressing results between various authors, it is difficult to compare our data with those from literature. For example, Cardador-Martínez et al. (2002) has found the total phenolic content of bean cv. Flor de Mayo to be 2.09 mg of catechin equivalents per gram of seeds. The same set of authors in another publication have reported the concentrations of phenolic compounds in six bean cultivars in the range 3.28-16.61 mg of catechin equivalents per gram of seeds (Oomah et al., 2005). The levels established by Vinson et al. (1998) using the Folin-Ciocalteu method has been 35.9 μmol and 31.9 μmol of catechin per gram of seeds for Kidney bean and Pinto bean, respectively. Wu et al. (2006) employing the latter method has determined from 2.23 to 12.47 mg of phenolics (GAE) per gram of seeds in various bean cultivars. Yet another author, (Marzo et al. 2002), who has determined the total phenolic content of bean cv. Pinto by a method based on the Folin-Denis reagent, has obtained the value of 0.44 mg of phenolics per gram of seeds. The phenolic contents of twelve Italian cultivars of bean investigated by Heimler et al. (2005) have ranged between 1.17 and 4.40 mg GAE per gram of seeds. As mentioned before, besides the determination method itself, also the way of phenolic compound extraction from the study material is of great importance. However, the results of the present studies are close to those reported by Wu et al. (2006).
The antioxidant activity of bean genotypes, measured by the DPPH procedure, showed significant variations among genotypes (Table 2). The antioxidant activity of common bean genotypes varied in a wide range, cream colored CIAT genotype BAT 93 had the highest (36.96%) and black colored CIAT genotype MIB 465 had the lowest (17.09%) levels. Among the CIAT developed genotype, BAT 93 showed the highest antioxidant activity (36.93%) which is also the highest among all. Seven bean genotypes, BAT 93 (36.96%), MIB 154 (33.5%), T-39 (32.79%), MIB465 (17.09%), NUA 59 (23.08%), Ryder (29.05%) and Mayflower (28.94%) had higher seed antioxidant activity than those of others. The antioxidant activity decreased in the following order based on average values of bean market classes in the USA grown bean genotype: Small Red > Pinto > Navy > Great Northern > Black > Snap. Likely total polyphenol, the CIAT developed MIB bean series showed a high level of antioxidant activity except MIB 465 (17.09%). Tsuda et al. (1994) assessed the antioxidants of white, red and black bean seeds (Phaseolus vulgaris L.) and found that the seed coat and germ of the white varieties had no antioxidant activity. In contrast, the red and black seed coats had good antioxidant activity. This observation was later confirmed by Chou et al. (2003) who reported that a 50% ethanol extract of red beans had very good antioxidant activity and Wu et al. (2004) who found that red beans had very good in vivo antioxidant activity. Oomah et al. (2005) reported that dark red Kidney beans had the highest antioxidant activity while Navy beans had the lowest. However, the antioxidant activity of the Kidney beans was low compared to other beans. Beans such as Black, Cranberry and Pinto had good antioxidant and antiradical activity (Oomah et al., 2005). These observations suggest that the colored beans have greater antioxidant and antiradical properties than less colored beans. These values are different for Navy, Pinto, Black and Red beans previously reported (Wu et al., 2006) due to different extraction method, but generally higher than recent values (Wu et al., 2004).
In the present study (Fig. 1a-c) anthocyanin shows a significant positive correlation with the total polyphenol (r = 0.71; p<0.05) and antioxidant activity (r = 0.68; p<0.05). The anthocyanin rich vegetables, like black carrot and beetroots, showed high phenolic content and correspondingly high antioxidant activity. The results thus confirm that anthocyanin rich beans possess strong antioxidant activity (Wang et al., 1997; Velioglu et al., 1998). A strong correlation between total polyphenol content and antioxidant activity was observed (r = 0.86; p<0.05) and this finding suggests that total polyphenol content is a good predictor of in vitro antioxidant activity. Numerous studies have demonstrated the antioxidant activity of phenolic compounds.
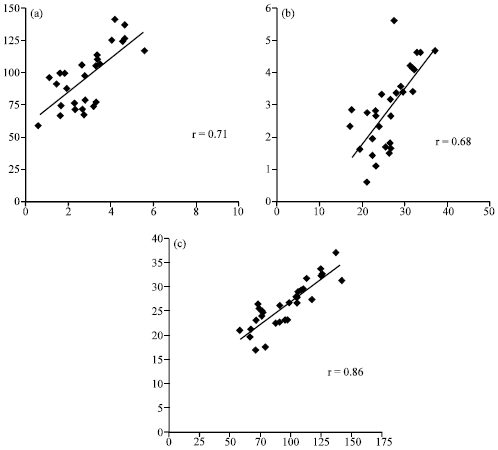 |
| Fig. 1: | Correlation among Anthocyanin, Total Polyphenol (TP) and Antioxidant Activity in 29 Common Bean Genotypes. Correlation between (a) anthocyanin and total polyphenol. (b) anthocyanin and antioxidant activity and (c) total polyphenol and antioxidant activity |
Oomah et al. (2005) reported that total phenolic content was the best indicator of the antioxidant activity of bean phenolics. They calculated that 40-71% of the antiradical activity could be explained by total phenolics and that flavonols were responsible for 20-39% of the antioxidant activity depending on the bean type. Phenolics enhance antioxidant activity due to their redox properties, which allow them to act as reducing agents, hydrogen donors and singlet oxygen quenchers (Rice-Evans et al., 1995). Thus, the antioxidant activity of a bean is due primarily to its phenolic content, which can be used as an indicator for assessing the antioxidant activity of the bean. Bean genotypes exhibited distinguishing differences in anthocyanin, total polyphenol and antioxidant activities. Generally bean genotypes with high anthocyanin and polyphenol content exhibit high antioxidant activity. Therefore, content and bioavailability of these health beneficiary components should not be overlooked in the selection of cultivars for phytochemical improvements and consumption for human foods. Considering the profile of bioactive compounds the genotypes T-39, XAN, BAT 93 and MIB 154 are promising. The information of this study can be used for selecting superior bean genotypes for targeted food and feed purposes and also for breeding programs. Such information can be used to closely scrutinize the strategy of breeding programs for selecting superior dry bean cultivars for targeted food and feed purposes.
REFERENCES
- Anderson, J.W., L. Story, B. Sieling, W.J. Chen, M.S. Petro and J. Story, 1984. Hypocholesterolemic effects of oat-bran or bean intake for hypercholesterolemic men. Am. J. Clin. Nutr., 40: 1146-1155.
Direct Link - Aparicio-Fernandez, X., L. Manzo-Bonilla and G. Loarca-Pina, 2005. Comparison of antimutagenic activity of phenolic compounds in newly harvested and stored common beans Phaseolus vulgaris against aflatoxin B1. J. Food Sci., 70: 73-78.
CrossRef - Awika, J.M., L.W. Rooney and R.D. Waniska, 2005. Anthocyanins from black sorghum and their antioxidant properties. Food Chem., 90: 293-301.
CrossRefDirect Link - Azevedo, L., J.C. Gomes, P.C. Stringheta, A.M.M.C. Gontijo, C.R. Padovani, L.R. Ribeiro and D.M.F. Salvadori, 2003. Black bean (Phaseolus vulgaris L.) as a protective agent against DNA damage in mice. Food Chem. Toxicol., 41: 1671-1676.
PubMed - Bazzano, L., J. He, L.G. Ogden, C. Loria, S. Vupputuri, L. Myers and P.K. Whelton, 2001. Legume consumption and risk of coronary heart disease in US men and women: NHANES i epidemiologic follow-up study. Arch. Int. Med., 161: 2573-2578.
Direct Link - Beninger, C.W. and G.L. Hosfield, 2003. Antioxidant activity of extracts, condensed tannin fractions and pure flavonoids from Phaseolus vulgaris L. seed coat color genotypes. J. Agric. Food Chem., 51: 7879-7883.
CrossRefPubMedDirect Link - Blair, M.W., F. Monserrate, S.E. Beebe, J. Restrepo and J.O. Flores, 2010. Registration of high mineral common bean germplasm lines NUA35 and NUA56 from the red-mottled seed class. J. Plant Reg., 4: 55-59.
Direct Link - Bourdon, I., B. Olson, R. Backus, B.D. Richter, P.A. Davis and B.O. Schneeman, 2001. Beans, as a source of dietary fiber, increase cholecystokinin and apolipoprotein b48 response to testmeals in men. J. Nutr., 131: 1485-1490.
Direct Link - Burits, M. and F. Bucar, 2000. Antioxidant activity of Nigella sativa essential oil. Phytother. Res., 14: 323-328.
CrossRefPubMedDirect Link - Cardador-Martinez, A., G. Loarca-Pina and B.D. Oomah, 2002. Antioxidant activity in common beans (Phaseolus vulgaris L.). J. Agric. Food Chem., 50: 6975-6980.
CrossRefDirect Link - Chou, S.T., W.W. Chao and Y.C. Chung, 2003. Antioxidative activity and safety of 50% ethanolic red bean extract (Phaseolus radiatus L. var. Aurea). J. Food Sci., 68: 21-25.
CrossRef - Choung, M., B. Choi, Y. An, Y. Chu and Y. Cho, 2003. Anthocyanin profile of korean cultivated kidney bean (Phaseolus vulgaris L.). J. Agric. Food Chem., 51: 7040-7043.
PubMed - Correa, P., 1981. Epidemiological correlations between diet and cancer frequency. Cancer Res., 41: 3685-3690.
Direct Link - Espinosa-Alonso, L.G., A. Lygin, J.M. Widholm, M.E. Valverde and O. Paredes-Lopez, 2006. Polyphenols in wild and weedy mexican common beans (Phaseolus vulgaris L.). J. Agric. Food Chem., 54: 4436-4444.
CrossRefPubMedDirect Link - Geil, P.B. and J.W. Anderson, 1994. Nutrition and health implications of dry beans: A review. J. Am. College Nutr., 13: 549-558.
CrossRefDirect Link - Gepts, P. and F.A. Bliss, 1986. Phaseolin variability among wild and cultivated common beans (Phaseolus vulgaris) from Colombia. Econ. Botony, 40: 467-478.
CrossRef - Gorinstein, S., O.J.M. Vargas, N.O. Jaramillo, I.A. Salas and A.L.M. Ayala et al., 2007. The total polyphenols and the antioxidant potentials of some selected cereals and pseudocereals. Eur. Food Res. Technol., 225: 321-328.
CrossRefDirect Link - Hangen, L. and M.R. Bennink, 2002. Consumption of black beans and navy beans (Phaseolus vulgaris) reduced azoxymethane-induced colon cancer in rats. Nutr. Cancer, 40: 60-65.
CrossRefPubMedDirect Link - Heimler, D., P. Vignolini, M.G. Dini and A. Romani, 2005. Rapid tests to assess the antioxidant activity of Phaseolus vulgaris L. dry beans. J. Agric. Food Chem., 53: 3053-3056.
CrossRefPubMedDirect Link - Horbowicz, M., R. Kosson, A. Grzesiuk and H. Dębski, 2008. Anthocyanins of fruits and vegetables - their occurrence, analysis and role in human nutrition. Vegetable Crops Res. Bull., 68: 5-22.
CrossRefDirect Link - Hughes, J.S., C. Ganthavorn and S. Wilson-Sanders, 1997. Dry beans inhibit azoxymethane-induced colon carcinogenesis in F344 rats. J. Nutr., 127: 2328-2333.
PubMed - Jain, M., G.T. Hislop, G.R. Howe and P. Ghadirian, 1999. Plant foods, antioxidants and prostate cancer risk: Findings from casecontrol studies in Canada. Nutr. Cancer, 34: 173-184.
PubMed - Khandaker, L., M.B. Ali and S. Oba, 2008. Total polyphenol and antioxidant activity of red amaranth (Amaranthus tricolor L.) as affected by different sunlight level. J. Jpn. Soc. Hortic. Sci., 77: 395-401.
Direct Link - Kolonel, L.N., J.H. Hankin, A.S. Whittemore, A.H. Wu and R.P. Gallagher et al., 2000. Vegetables, fruits, legumes and prostate cancer: A multiethnic case-control study. Cancer Epidemiol. Biomark Prev., 9: 795-804.
PubMedDirect Link - Marzo, F., R. Alonso, E. Urdaneta, F.J. Arricibita and F. Ibanez, 2002. Nutritional quality of extruded kidney bean (Phaseolus vulgaris L. var. pinto) and its effects on growth and skeletal muscle nitrogen fractions in rats. J. Anim. Sci., 80: 875-879.
Direct Link - McClean, P.E., R.K. Lee and P.N. Miklas, 2004. Sequence diversity analysis of dihydroflavanol reductase intron 1 in common bean. Genome, 47: 266-280.
PubMed - McClean, P.E., R.K. Lee, C. Otto, P. Gepts and M.J. Bassett, 2002. Molecular and phenotypic mapping of genes controlling seed coat pattern and color in common bean (Phaseolus vulgaris L.). J. Heredity, 93: 148-152.
Direct Link - Neff, M.M. and J. Chory, 1998. Genetic interactions between phytochrome A, phytochrome B and cryptochrome 1 during Arabidopsis development. Plant Physiol., 118: 27-35.
PubMed - Oomah, B.D., A. Cardador-Martinez and G. Loarca-Pina, 2005. Phenolics and antioxidative activities in common beans (Phaseolus vulgaris L.). J. Sci. Food Agric., 85: 935-942.
CrossRef - Rabino, I. and A.L. Mancinelli, 1986. Light, temperature and anthocyanin production. Plant Physiol., 81: 922-924.
Direct Link - Rice-Evans, C.A., N.J. Miller, P.G. Bolwell, P.M. Bramley and J.B. Pridham, 1995. The relative antioxidant activities of plant-derived polyphenolic flavonoids. Free Radic. Res., 22: 375-383.
CrossRefPubMedDirect Link - Romani, A., P. Vignolini, C. Galardi, N. Mulinacci, S. Benedettelli and D. Heimler, 2004. Germplasm characterization of zolfino landraces (Phaseolus Vulgaris L.) by flavonoid content. J. Agric. Food Chem., 52: 3838-3842.
CrossRef - Slinkard, K. and V.L. Singleton, 1977. Total phenol analysis: Automation and comparison with manual methods. Am. J. Enol. Vitic, 28: 49-55.
Direct Link - Venkateswaran, S., L. Pari and G. Saravanan, 2002. Effect of Phaseolus vulgaris on circulatory antioxidants and lipids in rats with streptozotocin-induced diabetes. J. Med. Food, 5: 97-103.
Direct Link - Vinson, J.A., Y. Hao, X. Su and L. Zubik, 1998. Phenol antioxidant quantity and quality in foods: Vegetables. J. Agric. Food Chem., 46: 3630-3634.
CrossRefDirect Link - Velioglu, Y.S., G. Mazza, L. Gao and B.D. Oomah, 1998. Antioxidant activity and total phenolics in selected fruits, vegetables and grain products. J. Agric. Food Chem., 46: 4113-4117.
CrossRefDirect Link - Wang, H., G. Cao and R.L. Prior, 1997. Oxygen radical absorbing capacity of anthocyanins. J. Agric. Food Chem., 45: 304-309.
CrossRef - Wu, X., G.R. Beecher, J.M. Holden, D.B. Haytowitz, S.E. Gebhardt and R.L. Prior, 2006. Concentrations of anthocyanins in common foods in the United States and estimation of normal consumption. J. Agric. Food Chem., 54: 4069-4075.
CrossRefDirect Link - Wu, X., G.R. Beecher, J.M. Holden, D.B. Haytowitz, S.E. Gebhardt and R.L. Prior, 2004. Lipophilic and hydrophilic antioxidant capacities of common foods in the United States. Agric. Food Chem., 52: 4026-4037.
CrossRefPubMedDirect Link - Williams, C.A., J.C. Onyilagha and J.B. Harborne, 1995. Flavonoid profilesin leaves, flowers and stems of forty-nine members of the phaseolinae. Biochem. Syst. Ecol., 23: 655-667.
CrossRef - Xu, B. and S. Chang, 2009. Total phenolic, phenolic acid, anthocyanin, flavan-3-ol and flavonol profiles and antioxidant properties of pinto and black beans (Phaseolus vulgaris L.) as affected by thermal processing. J. Agric. Food Chem., 57: 4754-4764.
PubMed - Xu, B.J., S.H. Yuan and S.K.C. Chang, 2007. Comparative analyses of phenolic composition, antioxidant capacity and color of cool season legumes and other selected food legumes. J. Food Sci., 72: S167-S177.
CrossRefDirect Link