
A.S.M. Golam Masum Akond
Division of Science and Mathematics, Mayville State University, ND, USA
Heath Crawford
Division of Science and Mathematics, Mayville State University, ND, USA
Janelle Berthold
Division of Science and Mathematics, Mayville State University, ND, USA
Zahirul I. Talukder
Division of Science and Mathematics, Mayville State University, ND, USA
Khwaja Hossain
Division of Science and Mathematics, Mayville State University, ND, USA
American Journal of Food Technology
Year: 2011 | Volume: 6 | Issue: 3 | Page No.: 235-243







ABSTRACT
In this study, the variation of zinc (Zn), iron (Fe), calcium (Ca) and magnesium (Mg) and the interference of phytic acid (PA) on their availability was investigated in 29 US grown and CIAT breeding genotypes of common bean. Fe levels showed the highest variation (8.9-112.9 mg kg-1) followed by Ca (58.67-122.98 mg kg-1) and Zn (30.90-64.60 mg kg-1) while variability of Mg concentration (6.47- 11.05 mg kg-1) is the least among the mineral components. PA showed a wide range of variability (12.52- 316.42 m kg-1) and inversely correlated with Fe, Ca and Mg concentrations. The results of the minerals and PA concentration can be interpreted in terms of expected bio-availability of minerals and the correlation study indicated that the presence of high concentration of PA inhibit the availability of most minerals under study in common beans. We suggest that the genotypes, MIB466, MIB465, MIB152 and JaloEEP 558 could be considered as sources of high Zn and Vista and NUA56-1770 for high seed Fe. We also identified G122 for high Ca and JaloEEP558 genotype for high Mg. We conclude that there is scope for the enhancement of mineral contents of common bean by selecting suitable genotype and bean products require processing for dephytinization for the improvement of mineral availability.
PDF Abstract XML References Citation
Received: February 22, 2010;
Accepted: May 24, 2010;
Published: July 05, 2010
How to cite this article
A.S.M. Golam Masum Akond, Heath Crawford, Janelle Berthold, Zahirul I. Talukder and Khwaja Hossain, 2011. Minerals (Zn, Fe, Ca and Mg) and Antinutrient (Phytic Acid) Constituents in Common Bean. American Journal of Food Technology, 6: 235-243.
DOI: 10.3923/ajft.2011.235.243
URL: https://scialert.net/abstract/?doi=ajft.2011.235.243
DOI: 10.3923/ajft.2011.235.243
URL: https://scialert.net/abstract/?doi=ajft.2011.235.243
INTRODUCTION
Common beans (Phaseolus vulgaris L.) are the most important grain legumes for direct human consumption in the world (Broughton et al., 2003). It has been considered as an important and inexpensive source of protein, dietary fiber, vitamins, minerals and bioactive compounds (Nyombaire et al., 2007). Compared with meat-based diets, plant-based diets are often limited in the content and bioavailability of essential minerals such as zinc (Zn), iron (Fe), calcium (Ca) and magnesium (Mg). A major constraint to the availability of minerals is the presence of toxic and antinutrient constituents like Phytic Acid (PA). Phytic acids chelate several mineral elements, especially Zn, Fe, Ca, Mg and Mo and interfere with their absorption and utilization (Ologhobo, 1980). The world population, particularly in Latin America, Sub-Saharan Africa, the Caribbean and Southeast Asia, is at risk for micronutrient intake (Brown and Peerson, 2001). Recent reports indicate that Fe deficiency is the most prevalent micronutrient problem in the world, affecting over 2 billion people globally, many of whom depend on beans as their staple food (Welch, 1999). An estimated 49% of the world population is at risk for low Zn intake (Brown and Peerson, 2001). Children diets high in cereals suffer from Zn deficiencies (Ranum, 1999) and nutritional Zn deficiency is common throughout the world, including the USA (Ganapathy and Volpe, 1999). Calcium content in rural diets in developing countries is not adequate (Rosado et al., 1992; Wyatt and Triana-Tejas, 1994) and dietary Ca deficiency has been epidemiologically linked to several chronic diseases, including osteoporosis. Without adequate Mg, energy production falters and protein production is insufficient for normal growth and development of infants, children, adolescents and pregnant women (Mangels and Havala, 1994). The American diet is rich in proteins, carbohydrates and fats but it is commonly poor in magnesium (Mg). The minerals in beans are readily available, which is important in reducing the risks of osteoporosis (Dawson-Hughes et al., 1990) and hypertension (Appel et al., 1997).
Phytic acid represents from 65 to 85% of the total seed P (Reddy et al., 1989). As a polyanion, PA is an effective chelator of positively charged molecules and has the potential to form stable insoluble complexes with minerals and proteins. These complexes confer PA with antinutritional properties, particularly in the humans and nonruminants lacking the hydrolytic enzyme phytase in their digestive tract (Cheryan, 1980). In some aspects, such as PA is considered beneficial source of nutrients during bean seed germination (Raboy, 2001), an antioxidant and anticarcinogen and may have beneficial role on human health, particularly for aged people (Zhou and Erdman, 1995). In order to improve availability of minerals such as Fe and Zn availability in foods, studies have been conducted to reduce the levels of PA in the seeds of various plants (Lucca et al., 2001). For instance, development of low PA grains of maize (Zea mays), barley (Hordeum vulgare) and rice (Oryza sativa) to improve availability of micronutrients (Larson et al., 1998, 2000; Raboy et al., 2001).
Common bean is considered as one of the most beneficial crop species and the knowledge of micronutrients and their availability in this species is essential for human health management.
The level, bioavailability of different micronutrients and their relationships with antinutrients, PA are crucial for pedigree selection and breeding common beans aiming to enhance micronutrients content (Welch and Graham, 2004). We conducted this study using 29 genotypes consist of US grown and CIAT (International Center for Tropical Agriculture) developed genotypes to understand the level of different minerals and the interference of PA on their availability.
MATERIALS AND METHODS
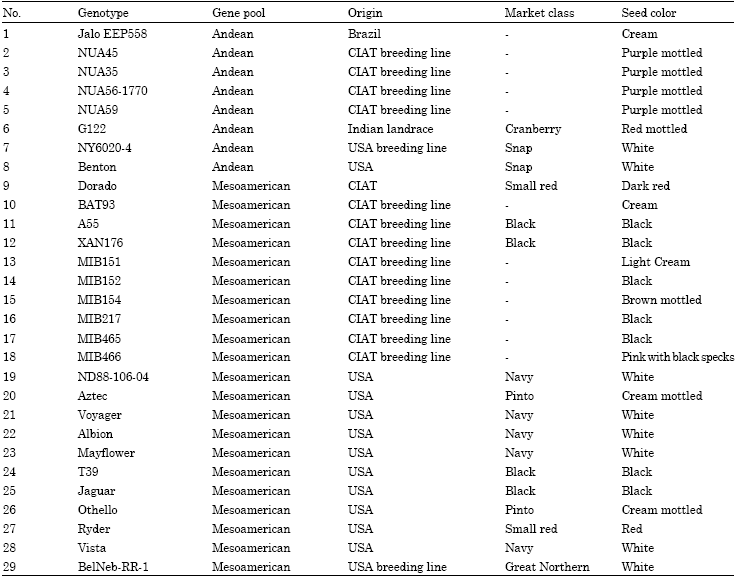
Plant material: The 29 common bean genotypes in this study consisted of 14 genotypes from CIAT, 13 from the USA and one each from Brazil and India. All common bean genotypes and their origin, are listed in Table 1. Common bean genotypes were grown in the greenhouse in 18x19 cm pots filled with Sunshine mix 1 (Sun Gro Horticulture Canada Ltd., formulated with Canadian Sphagnum peat moss, coarse grade perlite, gypsum and Dolomitic lime) as substrate. The seeds were planted on March 27, 2008 following a Randomized Complete Block Design (RCBD) with three replications. Two seeds for each genotype were placed in each pot for germination but one plant was allowed to grow until harvest of the seeds. Pots were watered periodically with tap water to the approximate field capacity to facilitate normal plant growth. No additional fertilizer or pesticide was applied during the period of experimentation.
| Table 1: | Common bean genotypes, gene pool group, origin, seed color |
 | |
Chemical analysis: After harvesting, seeds from each pod of individual plant were mixed thoroughlyand randomly selected 100 seed were weighed. For chemical analysis, 10 randomly selected seeds were washed with deionized water containing Joy detergent (Proctor and Gamble, Cincinnati, OH) and later rinsed with deionized water only. Samples were oven-dried at 70°C for 48 h, weighed and ground in an agate mortar with an agate pestle (Brinkmann Instruments Co., Westbury, NY). A 300 mg aliquot of the ground material was processed for concentrated nitric acid digestion, followed by 30% hydrogen peroxide. Fe and Zn concentrations were determined following the method described by Moraghan and Grafton (2001). Magnesium and Ca were determined from the samples by dry ashing method that described by Chapman and Pratt (1961). Concentrations of Zn, Fe, Mg and Ca were converted and expressed in mg kg-1 from the absorbance using Atomic Absorption Spectroscopy.
Phytic acid was determined using the ferric precipitation method as described by Raboy et al. (1984). Briefly, samples were extracted in 0.4 M HCl: 10% (w/v) sodium sulfate. Following centrifugation, supernatant PA was precipitated as a ferric salt. Ferric phytates were washed, wet ashed and digest phytic acid phosphorus content determined colorimetrically using the method as described (Chen et al., 1956). Spectrophotometric reading was converted to PA by multiplying with the conversion factor 3.5484 and expressed as mg kg-1.
Statistical analysis: Each determination was carried out on three separate replications and analyzed in triplicate and values were then averaged. Data was assessed by the Analysis Of Variance (ANOVA) following Tukey's multiple range tests and significance was accepted at p<0.05 (Tukey, 1953). The PC software Excel Statistics (Version 5.0, Esumi Co. Ltd., Japan) was used for the calculations.
RESULTS AND DISCUSSION
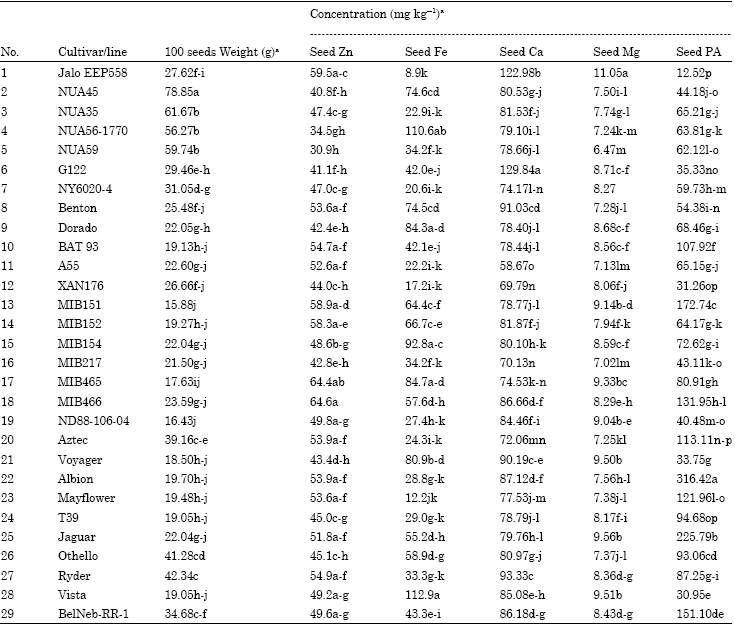
Seed mineral concentrations: The four mineral constituents (Zn, Fe, Ca and Mg) and the antinutrient component PA of US and CIAT common bean genotypes were determined (Table 2). Seed weight (100 seeds) ranged from 15.88 to 78.85 g with a mean of 29.97 g. The highest seed weight was recorded for Andean genotype NUA45 (78.85 g) and lowest for Mesoamerican genotype MIB152 (19.27 g), both of these genotypes were collected from the CIAT breeding program. Analysis of variance reveals that the seeds of 29 genotypes are significantly (p<0.05) differed in Zn, Fe, Ca and PA concentration except Mg. Seed Zn concentration ranged from 30.90 to 64.60 mg kg-1 and seven bean genotypes namely, MIB466 (64.6 mg kg-1), MIB465 (64.4 mg kg-1), JaloEEP558 (59.5 mg kg-1), MIB151 (58.9 mg kg-1), MIB152 (58.3 mg kg-1), Ryder (54.9 mg kg-1) and BAT93 (54.7 mg kg-1) had consistently higher seed Zn than others. Most of the MIB genotypes from CIAT have considerably high seed Zn than the US genotypes. The MIB genotypes are from CIAT breeding program and are selected for nutritional quality (Matthew Blair, personal communication), NUA59 also from CIAT but found lowest in Zn concentration. The Brazil originated Andean genotype JaloEEP558 and the US genotype Ryder also posses considerably high seed Zn concentration.
Wide range of variability of seed Fe concentration was found among the genotypes (8.9 to 112.9 mg kg-1). The higher level of seed Fe concentration was observed in the US navy bean genotype Vista (112.9 mg kg-1) and CIAT genotype NUA56-1770 (106.2 mg kg-1). Among all genotypes Brazil originated Andean genotype JaloEEP558 (8.9 mg kg-1) had the lowest seed Fe concentration. Seven bean genotypes, Vista (112.9 mg kg-1), NUA56-1770 (110.6 mg kg-1), MIB154 (92.8 mg kg-1), MIB465 (84.7 mg kg-1), Dorado (84.3 mg kg-1), Voyger (80.9 mg kg-1) and NUA45 (74.6 mg kg-1) had higher seed Fe concentration than those of others. NUA genotypes identified as high iron content developed by CIAT however, in our study, we did not find similar iron accumulation efficiency for all genotypes. The CIAT genotypes NUA35 and NUA59 have relatively lower level of Fe concentrations, while the genotype NUA45 has considerable high Fe concentration.
In an analysis of 2000 accessions at CIAT, a range of 34 to 89 mg kg-1 (average = 55 mg kg-1) for Fe and 21 to 54 mg kg-1 (average = 35 mg kg-1) for Zn were reported (Beebe et al., 2000). Some bean accessions from Peru were also found to contain high levels of Fe, averaging >100 mg kg-1 (Beebe et al., 2000; Islam et al., 2002). In this study we found some genotypes with higher level of Fe and Zn than previously reported by Beebe et al. (2000) and Islam et al. (2002). Beebe et al. (2000) also suggested that the seed Fe content in the Andean gene pool tended to present higher values than those from the Mesoamerican pool. In our study we found the Mesoamerican US bean genotype, Vista has the higher iron concentration than all of Andean genotypes. Cichy et al. (2005) reported high Zn concentration in Albion than Voyager, which is reverse in our findings. Recently, common bean genotypes, NUA35 and NUA56 have been registered for high seed mineral content at the CIAT (Blair et al., 2010). To our knowledge, these are the first registered genotypes specifically for nutritional quality in common beans. In our study we found high Fe concentration in NUA56 but the level of Zn concentration lower than some of the genotypes.
| Table 2: | Zinc (Zn), iron (Fe), calcium (Ca), magnesium (Mg) and phytic acid (PA) concentrations in the seeds of 29 common bean genotypes |
 | |
| aMeans having common letter do not differ significantly at p<0.05 | |
The discrepancy of our finding in mineral concentration might be due to difference in growing environment. We conducted our study in greenhouse condition which is reflected mostly the efficiency of accumulation of mineral by common bean genotypes. Our findings also indicate that seed Fe and Zn concentration was simple inherited and highly heritable traits across environments and soil types. Calcium concentrations were significantly (p<0.05) higher in Brazilian Andean genotype JaloEEP558 (122.98 mg kg-1) and Indian Andean genotype G122 (129.84 mg kg-1), where as the Mesoamerican CIAT genotypes XAN176 (69.79 mg kg-1) and MIB217 (70.13 mg kg-1) has lower concentration of Ca. On average, US bean genotypes namely Ryder, Voyager, BelNeb-RR-1 and Vista have high level of Ca concentration. Magnesium concentration ranged from 6.47 to 11.05 mg kg-1. The Brazilian Andean genotype JaloEEP558 (11.05 mg kg-1) has the highest seed Mg concentration, followed by US bean genotypes Vista (9.51mg kg-1) and Voyager (9.50 mg kg-1). The CIAT breeding lines NUA59 (6.47 mg kg-1), MIB217 (7.02 mg kg-1) and A55 (7.13 mg kg-1) has the lower level of seed Mg concentration.
The PA concentration of dry beans differed significantly (p<0.05) among genotypes and market classes (Table 2). PA concentration ranged from 12.52 to 316.42 mg kg-1 for the Brazilian Andean genotype JaloEEP558 and the US bean genotype Albion, respectively. Those two genotypes representing 25 folds difference in PA concentration between these two genotypes. Colored beans, particularly from the black and red market classes (Canadian navy bean cultivars), had significantly lower PA content than those of white bean (Oomah et al., 2008) but we did not find this trend in our study. Cichy et al. (2005) did not find any difference for PA concentration of seed between genotypes Albion and Voyager but we found significant differences between the two genotypes. Results (Tabe 2) showed that genotypes with low PA concentration have high contents Fe (e.g., Vista) and Ca (e.g., JaloEEP558), moderate level of Zn concentration (e.g., JaloEEP558). Again, in some cases genotypes with higher concentration showed low concentration of minerals. Research has shown that PA significantly inhibits the absorption minerals in the PA-laden whole grains and fresh legumes. A study on Fe absorption in cereal porridges reported, in some cases, a twelve-fold increase in the absorption of Fe when the PA was removed from the food (Hurrell et al., 2003). In several studies, it has been reported that human body can absorb about 30% of Mg and Zn without PA, however with PA, human body absorbed only 13 and 23%, respectively (Egli et al., 2004; Bohn et al., 2004). These results clearly indicate a low PA food product can provide a greater availability of minerals. In our study we also find similar trends. We identified few low PA contained genotypes namely, JaloEEP558, Vista, Xan176, Albion, Voyger and G122 with cumulatively high level of minerals concentration.
Correlations among phytic acid and other minerals: Non significant or weak correlations were found between 100 seeds weight and the concentration of seed Zn (r = 0.11; p<0.05), Fe (r = 0.02), Ca (r = 0.46) and Mg.(r = 0.27) concentration, where as PA inversely correlated with 100 seeds weight (r = -0.26; (Table 3). Marschner (1997) reported a significant correlation between seed weight and seed mineral concentration. However, Hacisalihoglu et al. (2004) indicated that largest seeded genotypes have high amount of total seed mineral content but not concentration. In our study, we found seed minerals, Zn, Fe, Mg and Ca were non significantly correlated with each other except Fe and Zn, which was inversely correlated (r = -0.11; p<0.05). It has been reported that the accumulation of seed Fe and Zn is significantly correlated and the genetic factors for increasing Fe are cosegregating with genetic factors for increasing Zn (Gregorio, 2002; Hacisalihoglu et al., 2005). Inverse correlation between Zn and Fe in our result did not support their findings which may be attributed with the wider variation in genomic constituents of the genotypes under study. Our results suggest the accumulation and enhancement of one mineral do not influence on the concentration of others and they are independently inherited in bean genome which is in agreement with Welch and Graham (2004).
| Table 3: | Correlation coefficients (r; p<0.05) for 100 seeds weight (SW), PA, Zn, Fe, Ca and Mg concentrations of beans |
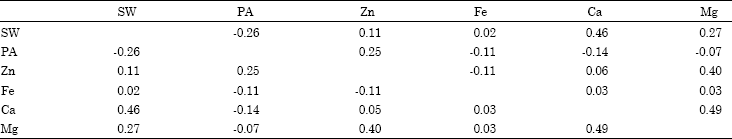 | |
Phytic acid concentration was inversely related to seed Fe (r = -0.11; p<0.05), Ca (r = -0.14) and Mg (r = -0.07) concentration. Among 29 genotypes, many genotypes contain higher concentration of phytic acid and low concentration of Fe, Ca and Mg (Table 2). Lower concentrations of Fe, Ca and Mg may be due to presence high concentration of PA. Because PA or phytate is a chelating agent, which is involved in binding minerals (such as K+, Ca2+, Zn2+, Fe2+, etc.) and making them unavailable for dietary absorption (Hirschi, 2009). It has been reported that cereals and legumes are rich in minerals but the bioavailability of these minerals is usually low due to the presence of antinutrient factor such as PA (Ann-Sofie 2002). Some genotypes, BelNeb-RR-1, Aztect or MIB151, we identified with high content of PA and correspondingly low content of most of the minerals (Table 2). Therefore, in order to evaluate mineral availability, PA should be considered as a major factor for common bean. Weak but non significant correlation (r = 0.25) between phytic acid and Zn concentration was observed in the genotypes tested, which suggest that breeding for increasing seed Zn may increase seed PA. We recommend screening PA levels in bean with high seed Zn concentration to ensure that increased levels of PA do not negate the value of gains in Zn in the diet.
CONCLUSION
We have identified a set of common bean genotypes contained high concentrations of seed minerals. Our data represent a comprehensive report on the genetic variation for several human health related nutrient concentrations in US grown and CIAT breeding genotypes representing both the Andean and Mesoamerican gene pools of common beans. We identified MIB466, MIB465, MIB152 and JaloEEP558 could be considered as sources of high seed Zn; Vista and NUA56-1770 for high seed Fe; G122 for high Ca and JaloEEP558 for high seed Mg genotypes. Our study also suggested that high concentration of PA inhibit the availability of most minerals in common beans. Thus, selection of low PA common bean may be important particularly for nutritional point of view.
ACKNOWLEDGMENTS
The authors thank Dr.s Phillip N. Miklas, USDA-ARS, Prosser, WA; Matthew Blair, CIAT, Columbia; and Juan Osorno, NDSU, ND for providing seeds for common bean genotypes. This research was partially supported by NIH grant P20 RR016741 from the NCRR and ND Department of Commerce grant through SUNRISE Byproducts Center of Excellences.
REFERENCES
- Sandberg, A.S., 2002. Bioavailability of minerals in legumes. Br. J. Nutr., 88: 281-285.
CrossRefDirect Link - Appel, L.J., T.J. Moore, E. Obarzanek, W.M. Vollmer and L.P. Svetkey et al., 1997. A clinical trial of the effects of dietary patterns on blood pressure. DASH collaborative research group. N. Engl. J. Med., 336: 1117-1124.
CrossRefPubMedDirect Link - Blair, M.W., F. Monserrate, S.E. Beebe, J. Restrepo and J.O. Flores, 2010. Registration of high mineral common bean germplasm lines NUA35 and NUA56 from the red-mottled seed class. J. Plant Reg., 4: 55-59.
Direct Link - Bohn, T., L. Davidsson, T. Walczyk and R.F. Hurrell, 2004. Phytic acid added to white-wheat bread inhibits fractional apparent magnesium absorption in humans. Am. J. Clin. Nutr., 79: 418-423.
Direct Link - Broughton, W.J., G. Hernandez, M. Blair, S. Beebe, P. Gepts and J. Vanderleyden, 2003. Beans (Phaseolus spp.)-model food legumes. Plant Soil, 252: 55-128.
CrossRefDirect Link - Brown, K.H., S.E. Wuehler and J.M. Peerson, 2001. The importance of zinc in human nutrition and estimation of the global prevalence of zinc deficiency. Food Nutr. Bull., 22: 113-125.
CrossRefDirect Link - Beebe, S., P.W. Skroch, J. Tohme, M.C. Duque, F. Pedraza and J. Nienhuis, 2000. Structure of genetic diversity among common bean landraces of mesoamerican origin based on correspondence analysis of RAPD. Crop Sci., 40: 264-273.
Direct Link - Cichy, K.A., S. Forster, K.F. Grafton and G.L. Hosfield, 2005. Inheritance of seed zinc accumulation in navy bean. Crop Sci., 45: 864-870.
Direct Link - Chen, P.S., T.Y. Toribara and H. Warner, 1956. Microdeterminations of phosphorous. Anal. Chem., 28: 1756-1758.
Direct Link - Dawson-Hughes, B., G.E. Dallal, E.A. Krall, L. Sadowski, N. Sahyoun and S. Tannenbaum, 1990. A controlled trial of the effect of calcium supplementation on bone density in postmenopausal women. N. Engl. J. Med., 323: 878-883.
Direct Link - Egli, I., L. Davidsson, C. Zeder, T. Walczyk and R. Hurrell, 2004. Dephytinization of a complementary food based on wheat and soy increases zinc, but not copper, apparent absorption in adults. J. Nutr., 134: 1077-1080.
Direct Link - Gregorio, G.B., 2002. Progress in breeding for trace minerals in staple crops. J. Nutr., 132: 500S-502S.
Direct Link - Ganapathy, S. and S.L. Volpe, 1999. Zinc, exercise and thyroid hormone function. Crit. Rev. Food Sci. Nutr., 39: 369-390.
CrossRefDirect Link - Hirschi, K.D., 2009. Nutrient biofortification of food crops. Annu. Rev. Nutr., 29: 401-421.
CrossRefDirect Link - Hacisalihoglu, G., J.J. Hart, C.E. Vallejos and L.V. Kochian, 2004. The role of shoot-localized processes in the mechanism of Zn efficiency in common bean. Planta, 218: 704-711.
Direct Link - Hurrell, R.F., M.B. Reddy, M.A. Juillerat and J.D. Cook, 2003. Degradation of phytic acid in cereal porridges improves iron absorption by human subjects. Am. J. Clin. Nutr., 77: 1213-1219.
Direct Link - Islam, F.M.A., K.E. Basford, C. Jara, R.J. Redden and S.E. Beebe, 2002. Seed compositional and disease resistance differences among gene pools in cultivated common bean. Gen. Resour. Crop Evol., 49: 285-293.
CrossRefDirect Link - Lucca, P., R. Hurrell and I. Potrykus, 2001. Approaches to improving the bioavailability and level of iron in rice seeds. J. Sci. Food Agric., 81: 828-834.
CrossRefDirect Link - Larson, S.R., J.N. Rutger, K.A. Young and V. Raboy, 2000. Isolation and genetic mapping of a non-lethal rice (Oryza sativa L.) low phytic acid mutation. Crop Sci., 40: 1397-1405.
Direct Link - Larson, S.R., K.A. Young, A. Cook, T.K. Blake and V. Raboy, 1998. Linkage mapping of two mutations that reduce phytic acid content of barley grain. Theor. Applied Genet., 97: 141-146.
CrossRefDirect Link - Moraghan, J.T. and K. Grafton, 2001. Genetic diversity and mineral composition of common bean seed. J. Sci. Food Agric., 81: 404-408.
CrossRefDirect Link - Mangels, A.R. and S. Havala, 1994. Vegan diets for women, infants and children. J. Agric. Environ. Ethics., 7: 111-122.
CrossRefDirect Link - Oomah, B.D., C. Blanchard and P. Balasubramanian, 2008. Phytic acid, phytase, minerals and antioxidant activity in canadian dry bean (Phaseolus vulgaris L.) cultivars. J. Agric. Food Chem., 56: 11312-11319.
CrossRefDirect Link - Raboy, V., 2001. Seeds for a better future: Low phytate grains help to overcome malnutrition and reduce pollution. Trends Plant Sci., 6: 458-462.
CrossRefDirect Link - Rosado, J.L., P. Lopez, M. Morales, E. Munoz and L.H. Allen, 1992. Bioavailability of energy, nitrogen, fat, zinc, iron and calcium from rural and urban Mexican diets. Br. J. Nutr., 68: 45-58.
CrossRefDirect Link - Raboy, V., P.F. Gerbasi, K.A. Young, S.D. Stoneberg and S.G. Pickett et al., 1984. Variation in seed total P, phytic acid, zinc, calcium, magnesium and protein among lines of Glycine max and G. sojae. Crop Sci., 24: 431-434.
Direct Link - Welch, R.M. and R.D. Graham, 2004. Breeding for micronutrients in staple food crops from a human nutrition perspective. J. Exp. Bot., 55: 353-364.
CrossRefDirect Link - Wyatt, C. and A. Triana-Tejas, 1994. Soluble and insoluble Fe, Zn, Ca and phytates in foods commonly consumed in northern Mexico. J. Agric. Food Chem., 42: 2204-2209.
Direct Link - Zhou, R.J. and J.W. Jr. Erdman 1995. Phytic acid in health and disease. Crit. Rev. Food Sci. Nutr., 35: 495-508.
CrossRefDirect Link