
B. Chutichudet
Department of Agricultural Technology, Faculty of Technology, Mahasarakham University, Mahasarakham 44000, Thailand
LiveDNA: 66.29804
P. Chutichudet
Department of Agricultural Technology, Faculty of Technology, Mahasarakham University, Mahasarakham 44000, Thailand
S. Kaewsit
Department of Agricultural Technology, Faculty of Technology, Mahasarakham University, Mahasarakham 44000, Thailand
American Journal of Food Technology
Year: 2011 | Volume: 6 | Issue: 3 | Page No.: 215-225







ABSTRACT
An analyzing study to evaluate the relationship between Polyphenol Oxidase (PPO) activity, internal qualities and degree of browning was conducted on lettuce var. Grand Rapids in order to understand the basis changes relating to the appearance of leaf browning and some quality characteristics. For PPO activity, Factorial experiment in completely randomized design was arranged with two factors: developmental stage (Factor A) at four levels (28, 42, 59 and 73 days after planting, DAP) with three different parts of plant (Factor B) (stem, leaf and root), while Completely Randomized Design (CRD) was applied for studying the internal qualities and browning appearance in leaves at 28, 42, 59 and 73 DAP. The experiment was carried out during May-July, 2009, with four replications, ten plants per replication. The results showed that each developmental stage and the various sections of the plant affected PPO activity. The leaf section of the plant particularly affected PPO, especially at 73 DAP. In addition, at the more mature stages of the leaf, more phenolic substance, ascorbic acid content and pH value were apparent, while quinone content decreased at advancing leaf age. In addition, as the plants reached an older stage, the leaf colour in terms of L* and b* decreased. Thus, the appropriate harvest for lettuce var. Grand Rapids should be at 59 DAP in order to face fewer incidence of browning and maintain good quality.
PDF Abstract XML References Citation
How to cite this article
B. Chutichudet, P. Chutichudet and S. Kaewsit, 2011. Influence of Developmental Stage on Activities of Polyphenol Oxidase, Internal Characteristics and Colour of Lettuce cv. Grand Rapids. American Journal of Food Technology, 6: 215-225.
DOI: 10.3923/ajft.2011.215.225
URL: https://scialert.net/abstract/?doi=ajft.2011.215.225
DOI: 10.3923/ajft.2011.215.225
URL: https://scialert.net/abstract/?doi=ajft.2011.215.225
INTRODUCTION
Lettuce (Lactuca sataiva), an important economic vegetable crop belonging to Compositae family (Asteraceae), is a popular vegetable and considered one of the most important all year round crops in Thailand. The lettuce planting area in all regions of the country is around 2, 119.2 ha with an estimated production of 15, 499.87 ton/year. Most of the lettuce is used for fresh consumption in fast foods and vegetable salads. Fresh lettuce is a significant source of dietary antioxidants. These include phenolics, ascorbic acid, carotenoids, tocopherols and glucosinolates, all of which have protective effects against various forms of cancer and cardiovascular and cerebrovascular diseases (Lister, 2003; Nicolle et al., 2004; Liorach et al., 2008). Moreover, lettuce is considered a low-acid food or health food (Dupont et al., 2000). Altunkaya and Gökmen (2008) reported that lettuce is highly susceptible to enzymatic browning. As such, leaf browning is a major problem that affects product losses resulting from rejection by consumers, shortened storage life and economic loss (Zhang et al., 2001). The appearance of this physiological disorders can be observed visually on leaf surfaces (Franck et al., 2007) during the preharvest period (Kays, 1999). Generally, browning in plants, was induced by several factors attributing to the activity of the PPO enzyme(Mayer, 1987). A loss of leaf colour is caused by enzymatic browning which is associated with the developmental stages of the plant. In plants, PPOs are localised in plastids, while plant phenolic substrates are mainly located in the vacuole. Therefore, the majority of enzymatic browning occurs when this sub-cellular compartmentation is lost (Rigal et al., 2000). Degl’Innocenti et al. (2007) also cited that the patterns of PPO activity could be related to their sensitive to browning incidence. However, there are few studies on the comparative browning level of lettuce during these growth stages. A further understanding of the browning appearance during the development stage is needed. The purposes of this study were threefold: quantify the distribution of PPO activity in the various organs of the lettuce plant, such as the stem, leaf or root; clarify internal substances related to enzyme-catalysed browning of fresh lettuce; compare the differences in leaf colour of Grand Rapids lettuces during its different maturity stages in order to determine the best quality. In addition, the quantification of the level of PPO activity and an understanding of the changes of other internal qualities at certain stages of development would be helpful for relating the occurrence of leaf browning to certain events and ultimately could provide a basis for predicting the optimal harvesting time in order to reduce quality loss.
MATERIALS AND METHODS
The research was carried out at the experimental field, Division of Agricultural Technology, Faculty of Technology, Mahasarakham University, in the Northeast of Thailand in the period between May to July, 2009. Lettuce cv. Grand Rapids was planted from seed. The seedlings were transplanted 25 DAP and grown singly in 2 L pot filled with a sandy loam soil : rice husk : manure ratio 1: 1 : 1 and placed under field conditions. A Factorial in Completely Randomized Design (CRD) was arranged and composed of two factors: developmental stage (Factor A) at four levels (28, 42, 59 and 73 DAP) with different parts of the plant (Factor B) at three levels (stem, leaf and root) for evaluation of PPO activities. Measurements for internal qualities and colour were conducted specifically on leaves at the same before mentioned developmental stages by using a CRD. Each treatment was carried out in four replicates, ten plants per replication. Plants were cut at each developmental stage and taken to the laboratory within a few minutes of cutting. The following were evaluated: (1) PPO activity was carried out according to the method reported by Jiang and Fu (1998). The attained enzyme extracts were measured by spectrophotometer model V-325-XS. One unit of PPO activity was defined as the amount of enzyme causing a change of 0.01 in absorbance (420 nm) per 60 sec. (2) Phenolic content was performed as described by Ribeiro et al. (2008). The content of phenolic was expressed as g/100 g fresh weight (FW) of leaf. (3) Quinone content was extracted as described by Pirie and Mullins (1976). Quinone content was expressed as absorbance at 437 nm per g fresh weight. (4) Ascorbic acid content was measured using juices squeezed from the flesh of the leaf with the use of distilled water at a ratio of 1:3. The measurements followed the method of AOAC (1990) and were expressed as mg ascorbic acid /100 mL juice (5) The measurement of pH values was carried out with the use of juices described earlier in number 4 and a pH meter ID 100D, from Singapore was used. (6) Leaf colour was measured on the leaf surface with a Hunter Lab Model No. 45/0-L, Serial No. 7092, USA. The CIE colour values L* (black = -100 and white = +100), a* (red) (- = green and + = yellow) and b* (yellowness) (- = blue and + = yellow) were measured to describe the colour of lettuce’s leaf. The collected data were statistically analyzed using the SPSS Version 6 (SPSS, 1999).
RESULTS
The recorded data received from measuring PPO activity, internal qualities and colour of ‘Grand Rapids’ lettuce at different ages (28, 42, 59 and 73 DAP) demonstrated the following results:
PPO activity: The results showed that PPO activity could be detected in various sections of the plant during the developmental stages. PPO activity increased sequentially when the plants entered their older stage. PPO activity increased quickly during plant growth and reached the maximal content at late harvesting of 73 DAP. In addition, changes in PPO activity were also influenced by different tissues. The results from Table 1 showed that PPO extracted from leaves had the highest activity, followed by the root and then the stem. For interaction of the developmental stage and various analyzed tissues, the results revealed that the highest PPO activity was observed in the leaf cut at late harvesting of 73 DAP (Table 1).
Phenolic: The phenolic content showed a surge of content during plant growth, suggesting the browning appearances of lettuce was affected by plant age. Significant changes in total phenolic content were found in lettuce leaves at different plant ages, although phenolic content in lettuce’s leaves featured the maximal content (2439.72 g/100 g FW) at the early developmental time (28 DAP). Afterwards, the levels of phenolics decreased dramatically to a value of 964.49 and 287.09 g /100 g FW at the ages of 42 and 59 DAP, respectively, then increasing markedly to a high level of 2425.54 g/100 g FW at the late-harvesting of 73 DAP (Table 2).
Quinone: With respect to quinone content, the results revealed a leaf from the early stage of 28 DAP showed the maximal quinone content of 0.1299 per g FW. Afterwards, quinone content from leaves of 42, 59 and 73 DAP showed insignificant quinone content of 0.0852, 0.0866 and 0.0935/g FW, respectively (Table 2).
Ascorbic acid content: Content of ascorbic acid showed an increasing trend with plant age. At 28 DAP, the lettuce leaf showed a significantly lower ascorbic acid level of 12.72 mg ascorbic acid/100 mL juice. Afterwards, there was a similarly high amount of ascorbic acid of 14.00-14.05 mg ascorbic acid/100 mL juice as growth progressed from day 42 to 73 DAP (Table 3).
pH: In a comparison of the pH value of lettuce’s leaves during plant growth, the data indicated that plant age had a highly significant effect on the pH value of leaves. The mean values of pH of leaf age at the early stage of 28 DAP showed the highest pH value of 7.20. Afterwards, as plant age reached 42 DAP and 59 DAP, the samples decreased the pH value remarkably to 5.52 and 5.98, respectively, then increased again to 6.81 as growth progressed to the late-harvest time at 73 DAP (Table 3).
Leaf colour: The results showed that leaf colour, measured by monitoring in terms of L*, a* and b* values, changed significantly during the different plant developmental stages. From Table 4, there were highly significant differences in the leaf colour in parameters of L* and b* between the four plant ages.
| Table 1: | Activities of PPO in different parts of lettuce during developing stage |
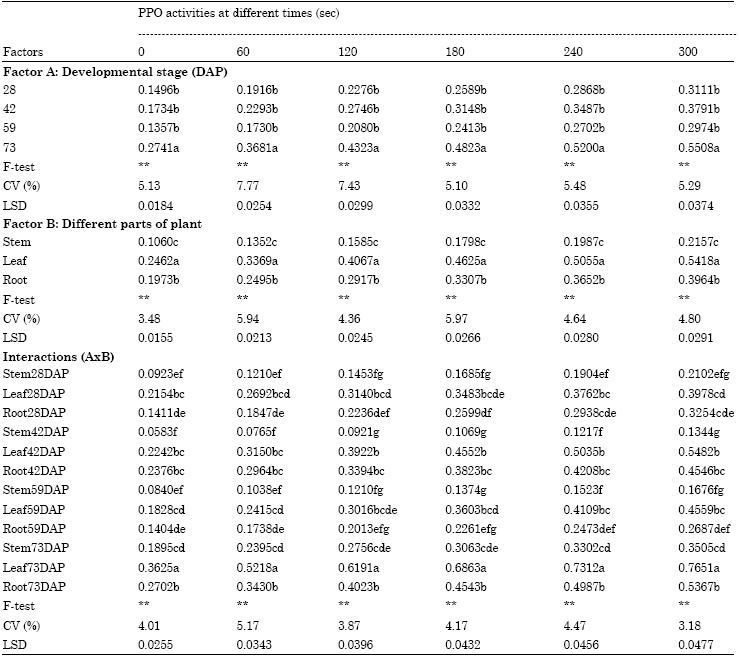 | |
| Letters within columns indicate Least Significant Differences (LSD) at **p = 0.01 | |
| Table 2: | Content of phenolic and quinone in leaf of lettuce during developing stage |
 | |
| Letters within columns indicate Least Significant Differences (LSD) at **p = 0.01 | |
| Table 3: | Ascorbic acid content and pH value of lettuce’s leaf during developing stage |
 | |
| Letters within columns indicate least significant differences (LSD) at **p = 0.01 | |
| Table 4: | Colour of lettuce during developing stage |
 | |
| Letters within columns indicate Least Significant Differences (LSD) at **p = 0.01, NS: Non significant | |
The results showed that at the early stage of 28 DAP, lettuce leaves showed the significantly highest L* and b* of 54.09 and 31.64, respectively. As the plant advanced through its developing stages, less colouring of the leaf in terms of L* and b* became apparent, while a* value among treatments showed no significant difference.
DISCUSSION
This study can provide an important basis for understanding leaf browning in lettuce. In this research, fresh ‘Grand Rapids’ lettuce, harvested at the different growth stages, was compared and analyzed for PPO activity, chemical characteristics and leaf color.
The PPO extracted from the lettuce plant at different stages of growth exhibited different levels of activity as shown in Table 1. It was obvious that PPO activity was present throughout the developmental stages. After 28 to 59 DAP, PPO enzymes maintained low activity. Activity then increased and reached maximum as the plants entered their advancing maturity stage, especially the later maturity stages. These results imply that lettuce has a potential for browning during the preharvest stage and especially at late-harvesting of 73 DAP. Generally, enzymatic browning of plant product was characterized by PPO activities (Peiser et al., 1998). Therefore, browning incidence in lettuce may be a development-associated activity of PPO based on the sensitivity of enzyme activity. This is in agreement with the results of Thipyapong et al. (1997) and Núñez-Delicado et al. (2007) who reported that PPO activity was considerably influenced by stages of development, such as tomato leaves and grapes. While Jiang et al. (2004) indicated that young tissues had significantly lower PPO activity than older tissues, this may be due to membrane integrity being maintained at the young stage. In addition, Lichanporn et al. (2009) cited that as the plant reached senescence and it led to a breakdown of the compartmentalization of enzymes, finally causing a rigorous symptom of enzymatic browning, especially harvest at a delayed maturity stage (Benjawan et al., 2008). Furthermore, these results were in agreement with Segovia-Bravo et al. (2009) who reported that in older plants the sub-cellular compartmentation was lost leading to intense leakage of PPO from the cells which in turn caused the plant to become sensitive to the browning appearance.
For the activity of the PPO analyzed from various tissues of lettuce, the results showed that the extent of PPO activity in ‘Grand Rapids’ lettuce was ubiquitous in various tissues. The total PPO activity of the leaf extracts was superior to that of the root and stem extracts. These results imply that the lettuce leaves were the most susceptible to browning compared with the root and stem, respectively. Yingsanga et al. (2008) cited that the higher activities of PPO in the leaf could be explained by higher rates of oxygen transmission into tissues due to their higher surface area and stomata density compared to the root and stem (Yingsanga et al., 2006). While, Collier and Wurr (1981); Collier and Huntington (1983) indicated the reason leaves of lettuce plants were the most rigorous in developing browning incidence because the leaves are often larger and more succulent due to their rapid expansion and higher water content than the other sections.
A compared interaction of different plant ages and various plant organs also affected different enzymatic activities of PPO in Grand Rapids lettuce. The comparison revealed that leaves harvested from a plant aged 73 DAP had the maximum PPO activity. The delayed harvested lettuce tended to exhibit a progressive increase in the susceptibility of enzymatic browning due to a loss of cellular compartmentation and membrane disintegration (Franck et al., 2007). Thus, the appropriate harvesting stage of lettuce should be done at 59 DAP in order to decrease the problem of leaf browning occurrence. Unfortunately, the identity of the different locations of PPO activity causing browning incidence in Grand Rapids lettuce is still unclear.
For the results on phenolic content, the results obtained from this study indicate that significant levels of phenolic components have been detected in lettuce and it was fluctuated according to plant age. Lettuce leaves at the early age of 28 DAP and the late stage of 73 DAP showed the highest phenolic content, while leaves from plant age of 59 DAP composed of only traces of phenolic substance. Thus, the concentrations of phenolic acids in lettuce were sensitive to the maturity stage. These results corresponded with King and Young (1999), who cited that the maturity stage was an important factor affecting the content of phenolic compounds. These results are also supported by the findings of Spanos and Wrolstad (1990), who claim that the phenolic content of pear depends primarily on level of maturity. Nevertheless, in lettuce leaves, as development progressed, the increase in PPO activity paralleled the increase in total phenol content. Thus, lettuces were susceptible to browning incidence during late-harvest due to an increase of both PPO activity and phenolic content and these facilitated the appearance of the browning disorder. This may be due to PPO, normally bound to membranes or walls, becoming active when released under altering plant metabolism and increasing phenolic synthesis when the plants reaches the advancing mature stage (Ruiz et al., 1999). This is consistent with the observation of Muñoz-Muñoz et al. (2009), who found that some phenolic compounds have been shown to be good substrates involved in the oxidation of polyphenols by PPO enzymes (Ke and Saltveit, 1989; Nicoli et al., 1991). These results were in agreement with the previous findings of Leoni et al. (1990) who reported that the phenolic compounds were the best substrate for artichoke PPO, while Vela et al. (2002) found the highest levels of PPO and phenolics occurred together in Algerie loquat at harvest. This coincidence of high PPO and phenolics enabled the loquat fruits to become more susceptible to enzymatic browning. Thus, both phenolic contents and PPO activity were found to be closely correlated to the degree of browning (Lee et al., 1990) because phenolic compounds were the major browning substrates present in plant tissues during late-harvesting (Sun et al., 2006). Furthermore, it may be possible that phenolic content might be largely regulated by PPO activity (Sun et al., 2009). While the opposite results were reported by Caldwell (2003) and Liorach et al. (2004) who cited that phenolic compounds acted as the main antioxidants in leafy vegetables. A general decrease of phenolic content was noticed in correspondence of maximum values of PPO activity (Chisari et al., 2010). Nevertheless, the major role of PPO in catalyzing the oxidation of phenolic substances and its involvement in the browning of lettuce leaves, was not fully understood and requires further investigation.
For quinone content, the results showed that the peak level for quinone content of lettuce leaf was observed only at early plant growth (28 DAP). Afterwards, quinone content decreased significantly and remained at stable levels as growth progressed during 42 to 73 DAP. Generally, the oxidation of phenolic compounds by the enzyme PPO produces o-diphenols. Then the o-diphenols mix together to produce o-quinones. These o-quinones were quite unstable and spontaneous polymerized to form brown, red or black pigments (Gawlik-Dziki et al., 2007; Ayaz et al., 2008). A gradual decrease in quinone content was responsible for browning in lettuce (Ke and Saltveit, 1986, 1989). Nevertheless, it is not clear whether the changed level of quinone was beneficial or detrimental to the browning incidence.
With ascorbic acid, the results showed that at the early stages of growth (28 DAP), the ascorbic acid content of lettuce’s leaf appeared to be at the lowest level of 12.72 mg ascorbic acid /100 mL juice. The level then increased at 42 DAP and maintained a similar level throughout the late growth period (73 DAP). Jeffery et al. (2003) found that a variation in the ascorbic acid contents of Brassica vegetables was caused by many factors including maturity at harvest. These results corresponded to Deepa et al. (2007) who found that ascorbic acid in fresh sweet pepper showed an increasing trend with advancing maturity. In addition, the increase of ascorbic acid in lettuce could have been attributed to synergism of this compound with a phenolic substance (Altunkaya and Gökmen, 2008). Núñez-Delicado et al. (2007) reported that ascorbic acid had a great capability for the prevention of the degradation of phenolic compounds in fresh-cut lettuce (Altunkaya and Gökmen, 2009). Therefore, the results from this study showed that PPO activity in lettuce has been shown to be positively associated with phenolic content and ascorbic acid content. In contrast, Sapers (1993) cited that ascorbic acid may act as an antioxidant agent to inhibit the catalytic action of PPO. However, the exact mechanism of ascorbic acid on the mechanism of browning occurrence in lettuce is not yet clear. Further work is required to better understand the biochemical mechanism of ascorbic acid related to the browning appearance in Grand Rapids lettuces.
For pH value, the results showed that different developmental stages exhibited different pH values. The pH value of lettuce’s leaves had the highest value of 7.20 at an early developmental stage (28 DAP). Afterwards, the pH levels were markedly reduced at 42 and 59 DAP and then they significantly increased again to the neutral level of 6.81 at late harvesting (73 DAP). Thus, the developmental stages of the plant affected pH alteration in the lettuce leaves. These results were in agreement with previous findings (Fang et al., 2007). In addition, Sun et al. (2009) found that the severity of PPO activity was pH-dependent. Generally the maximum PPO activity in most plants is at or near neutral pH values of 7.0 (Gawlik-Dziki et al., 2008). Thus, the activity of PPO was found to promote rapidly under the condition of higher pH of 6.81 at 73 DAP, while a lower pH at 42 and 59 DAP effected a decrease of PPO activity as a result of inappropriate pH (Altunkaya and Gökmen, 2008).
For the results on leaf colour, it was found that leaf colour in terms of L* (lightness) and b* (blue-yellow) tended to decrease their values with advancing developmental time, while the a* value (green-red) showed no significant difference irrespective o f plant ages. At early plant growth of 28 DAP, both L* and b* exhibited their highest level, then these values tended to decrease at 42 DAP through the 73 DAP. These results implied that as plant age advances, leaf colour changed toward a less bright green colour. This could be explained by, with the extension of development, increased PPO activity was found to be correlated with the susceptibility to browning in fresh lettuce measured by leaf colour (Zhang et al., 2000; De Castro et al., 2008).
In conclusion, PPO were found in various tissues of lettuce plants, especially in the leaf of plants aged at 73 DAP when the highest PPO activity was measured. As the plants reached an older stage, the contents of total phenol, ascorbic acid and pH increased, while quinone content and leaf colour in terms of L* and b* decreased. These findings show that as the lettuce plant reached an older stage, the plants were more sensitive to browning appearance and lower quality.
ACKNOWLEDGMENTS
This research was funded by the Mahasarakham University under project No. 5201089/2552. The authors wish to express their sincere thanks to the Financial Office for financial assistance. Authors gratefully acknowledge Mr. Paul Dulfer for revising the manuscript.
REFERENCES
- Altunkaya, A. and V. Gokmen, 2008. Effect of various inhibitors on enzymatic browning, antioxidant activity and total phenol content of fresh lettuce (Lactuca sativa). Food Chem., 107: 1173-1179.
CrossRefDirect Link - Altunkaya, A. and V. Gokmen, 2009. Effect of various anti-browning agents on phenolic compounds profile of fresh lettuce (L. sativa). Food Chem., 117: 122-126.
CrossRef - Ayaz, F.A., O. Demir, H. Torun, Y. Kolcuoglu and A. Colak, 2008. Characterization of polyphenoloxidase (PPO) and total phenolic contents in medlar (Mespilus germanica L.) fruit during ripening and over ripening. Food Chem., 106: 291-298.
CrossRef - Caldwell, C.R., 2003. Alkylperoxyl radical scavenging activity of red leaf lettuce (Lactuca sativa L.) phenolics. J. Agric. Food Chem., 51: 4589-4595.
CrossRefPubMedDirect Link - Chisari, M., A. Todaro, R.N. Barbagallo and G. Spagna, 2010. Salinity effects on enzymatic browning and antioxidant capacity of fresh-cut baby Romaine lettuce (Lactuca sativa L. cv. Duende). Food Chem., 119: 1502-1506.
CrossRef - Benjawan, C., P.T. Chutichude and S. Khumkratok, 2008. Effect of harvesting index on browning reaction and changes of tissue structure in santol fruits. Pak. J. Biol. Sci., 11: 1212-1219.
CrossRefPubMedDirect Link - Collier, G.F. and V.C. Huntington, 1983. The relationship between leaf growth, calcium accumulation and distribution and tipburn development in field-grown butterhead lettuce. Scientia Hortic., 21: 123-128.
CrossRef - Collier, G.F. and D.C.E. Wurr, 1981. The relationship of tipburn incidence in head lettuce to evaporative water loss and leaf dimensions. J. Hortic. Sci. Biotechnol., 56: 9-14.
Direct Link - De Castro, E., D.M. Barrett, J. Jobling and E.J. Mitcham, 2008. Biochemical factors associated with a CO2-induced flesh browning disorder of Pink Lady apples. Posthar. Biol. Technol., 48: 182-191.
CrossRefDirect Link - Deepa, N., C. Kaur, B. George, B. Singh and H.C. Kapoor, 2007. Antioxidant constituents in some sweet pepper (Capsicum annuum L.) genotypes during maturity. LWT-Food Sci. Technol., 40: 121-129.
CrossRef - Degl`Innocenti, E., A. Pardossi, F. Tognoni and L. Guidi, 2007. Physiological basis of sensitivity to enzymatic browning in lettuce, escarole and rocket salad when stored as fresh-cut products. Food Chem., 104: 209-215.
CrossRef - Dupont, S., Z. Mondi, G. Willamson and K. Price, 2000. Effect of variety, processing and storage on the flavonoid glycoside and composition of lettuce and chicory. J. Agric. Food Chem., 48: 3957-3964.
CrossRefDirect Link - Fang, Z., M. Zhang, S. Yunfei and J. Sun, 2007. Polyphenol oxidase from bayberry (Myrica rubra Sieb. et Zucc.) and its role in anthocyanin degradation. Food Chem., 103: 268-273.
CrossRefDirect Link - Franck, C., J. Lammertyn, Q.T. Ho, P. Verboven, B. Verlinden and B.M. Nicolai, 2007. Browning disorders in pear fruit. Postharvest Biol. Technol., 43: 1-13.
CrossRef - Gawlik-Dziki, U., U. Szymanowska and B. Baraniak, 2007. Characterization of polyphenol oxidase from broccoli (Brassica oleracea var. botrytis italica) florets. Food Chem., 105: 1047-1053.
CrossRef - Gawlik-Dziki, U., U. Zlotek and M. Swieca, 2008. Characterization of polyphenol oxidase from butter lettuce (Lactuca sativa var. capitata L.). Food Chem., 107: 129-135.
CrossRef - Jeffery, E.H., A.F. Brown, A.C. Kurilich, A.S. Keck, N. Matusheski, B.P. Klein and J.A. Juvik, 2003. Variation in content of bioactive components in broccoli. J. Food Compos. Anal., 16: 323-330.
CrossRefDirect Link - Jiang, Y. and J. Fu, 1998. Inhibition of polyphenol oxidase and the browning control of litchi fruit by glutathione and citric acid. Food Chem., 62: 49-52.
CrossRefDirect Link - Jiang, Y., X. Duan, D. Joyce, Z. Zhang and J. Li, 2004. Advances in understanding of enzymatic browning in harvested litchi fruit. Food Chem., 88: 443-446.
CrossRefDirect Link - Kays, S.J., 1999. Preharvest factors affecting appearance. Postharvest Biol. Technol., 15: 233-247.
CrossRefDirect Link - Ke, D. and M.E. Saltveit, 1986. Effects of calcium and auxin on russet spotting and phenylalanine ammonia-lyase activity in iceberg lettuce. Hort. Sci., 21: 1169-1171.
Direct Link - Ke, D. and M.E. Saltveit Jr., 1989. Wound-induced ethylene production, phenolic metabolism and susceptibility to russet spotting in Iceberg lettuce. Physiol. Planta., 76: 412-418.
CrossRefDirect Link - King, A. and G. Young, 1999. Characteristics and occurrence of phenolic phytochemicals. J. Am. Dietetic Assoc., 99: 213-218.
CrossRefPubMedDirect Link - Lee, C.Y., V. Kagan, A.W. Jaworski and S.K. Brown, 1990. Enzymatic browning in relation to phenolic compounds and polypheoloxidase activity among various peach cultivars. J. Agric. Food Chem., 38: 99-191.
CrossRefDirect Link - Leoni, O., S. Palmeri, V.L. Christiaan and F.V. Sumere, 1990. Polyphenol oxidase from architoke (Cynara scolymus L.). Food Chem., 38: 27-39.
CrossRef - Lichanporn, I., V. Srilaong, C. Wongs-Aree and S. Kanlayanarat, 2009. Postharvest physiology and browning of longkong (Aglaia dookkoo Griff.) fruit under ambient conditions. Postharvest Biol. Technol., 52: 294-299.
CrossRef - Llorach, R., A. Martínez-Sánchez, F.A. Tomás-Barberán, M.I. Gil and F. Ferreres, 2008. Characterisation of polyphenols and antioxidant properties of five lettuce varieties and escarole. Food Chem., 108: 1028-1038.
CrossRefDirect Link - Liorach, R., F.A. Tomas-Barberan and F. Farreras, 2004. Lettuce and chicory byproducts as a source of antioxidant phenolic extracts. J. Agric. Food Chem., 52: 5109-5116.
CrossRefDirect Link - Mayer, A.M., 1987. Polyphenoloxidase in plants-recent progress. Phytochemistry, 26: 11-20.
Direct Link - Munoz-Munoz, J.L., F. Garcia-Molina, P.A. Garcia-Ruiz, E. Arribas, J. Tudela, F. Garcia-Canovas and J.N. Rodriguez-Lopez, 2009. Enzymatic and chemical oxidation of trihydroxylated phenols. Food Chem., 113: 435-444.
CrossRef - Nicoli, M.C., B.E. Elizalbe, A. Piotti and C.R. Lerici, 1991. Effects of sugars and maillard reaction products on polyphenol oxidase activity in food. J. Food Biochem., 15: 169-184.
CrossRefDirect Link - Nicolle, C., N. Cardinault, E. Gueux, L. Jaffrelo and E. Rock et al., 2004. Health effect of vegetable-based diet: Lettuce consumption improves cholesterol metabolism and antioxidant status in the rat. Clin. Nutr., 23: 605-614.
CrossRefPubMedDirect Link - Nunez-Delicado, E., M. Serrano-Megias, A.J. Perez-Lopez and J.M. Lopez-Nicolas, 2007. Characterization of polyphenol oxidase from Napoleon grape. Food Chem., 100: 108-114.
CrossRef - Peiser, G., G. Lopez-Galvez, M. Cantwell and M.E. Saltveit, 1998. Phenylalanine ammonia lyase inhibitors control browning of cut lettuce. Postharvest Biol. Technol., 14: 171-177.
CrossRefDirect Link - Pirie, A. and M.G. Mullins, 1976. Changes in anthocyanin and phenolic content of grapevine leaf and fruit tissue treated with sucrose, nitrate and abscisic acid. Plant Physiol., 58: 468-472.
PubMedDirect Link - Ribeiro, S.M.R., L.C.A. Barbosa, J.H. Queiroz, M. Knodler and A. Schieber, 2008. Phenolic compounds and antioxidant capacity of Brazilian mango (Mangifera indica L.) varieties. Food Chem., 110: 620-626.
CrossRefDirect Link - De Rigal, D., F. Gauillard and F. Richard-Forget, 2000. Changes in the carotenoid content of apricot (Prunus armeniaca, var Bergeron) during enzymatic browning: Beta-carotene inhibition of chlorogenic acid degradation. J. Sci. Food Agric., 80: 763-768.
CrossRefDirect Link - Ruiz, J.M., P.C. Garcia, R.M. Rivero and J. Romero, 1999. Response of phenolic metabolism to the application of carbendazim plus boron in tobacco. Physiol. Plant., 106: 151-157.
CrossRefDirect Link - Sapers, G.M., 1993. Browning of foods: Control by sulfites, antioxidants and other means. Food Technol., 47: 75-84.
Direct Link - Segovia-Bravo, K.A., M. Jaren-Galan, P. Garc�a-Garc�a and A. Garrido-Fernandez, 2009. Browning reactions in olives: Mechanism and polyphenols involved. Food Chem., 114: 1380-1385.
CrossRef - Spanos, G.A. and R.E. Wrolstad, 1990. Influence of variety, maturity, processing and storage on the phenol composition of pear juice. J. Agric. Food Chem., 38: 817-824.
CrossRefDirect Link - Sun, J., Y. Jiang, X. Wei, J. Shi and Y. You et al., 2006. Identification of (-)-epicatechin as the direct substrate for polyphenol oxidase isolated from litchi pericarp. Food Res. Int., 39: 864-870.
CrossRef - Sun, J., X. Xiang, C. Yu, J. Shi and H. Peng et al., 2009. Variations in contents of browning substrates and activities of some related enzymes during litchi fruit development. Scientia Hortic., 120: 555-559.
CrossRef - Thipyapong, P., D.M. Joel and J.C. Steffens, 1997. Differential expression and turnover of the tomato polyphenol oxidase gene family during vegetative and reproductive development. Plant Physiol., 113: 707-718.
Direct Link - Vela, J.C., S.S. Marchart, I.G. Lucas and R.B. Martinez, 2002. Evolution of Phenolics and Polyphenoloxidase Isoenzymes in Relation to Physical-Chemical Parameters During Loquat (Eriobotrya japonica cv. Algerie) Fruit Development and Ripening. In: First International Symposium on Loquat, Lacer, G. and M.L. Badenes (Eds.). Zaragoza: CIHEAM-IAMZ, Valencia, Spain, pp: 161-164.
Direct Link - Yingsanga, P., V. Srilaong, S. Kanlayanarat, S. Noichinda and W.B. McGlasson, 2008. Relationship between browning and related enzymes (PAL, PPO and POD) in rambutan fruit (Nephelium lappaceum Linn.) cvs. Rongrien and See-chompoo. Postharvest Biol. Technol., 50: 164-168.
CrossRef - Yingsanga, P., V. Srilaong, W.B. McGlasson, E. Kabanoff, S. Kanlayanarat and S. Noichinda, 2006. Morphological differences associated with water loss in Rambutan fruit cv. Rongrien and See-chompoo. Acta Hortic., 712: 453-460.
Direct Link - Zhang, D., P.C. Quantick and J.M. Grigor, 2000. Changes in phenolic compounds in litchi (Litchi chinensis Sonn.) fruit during postharvest storage. Postharvest Biol. Technol., 19: 165-172.
CrossRefDirect Link - Zhang, Z., X. Pang, Z. Ji and Y. Jiang, 2001. Role of anthocyanin degradation in litchi pericarp browning. Food Chem., 75: 217-221.
CrossRefDirect Link