
Sathyanarayana N. Gummadi
Applied and Industrial Microbiology Laboratory, Department of Biotechnology, Indian Institute of Technology-Madras, Chennai 600 036, India
LiveDNA: 91.30544
Sawan Kumar
Applied and Industrial Microbiology Laboratory, Department of Biotechnology, Indian Institute of Technology-Madras, Chennai 600 036, India
American Journal of Food Technology
Year: 2008 | Volume: 3 | Issue: 6 | Page No.: 354-360







ABSTRACT
The effect of temperature and salts on the growth of halotolerant yeast, Debaryomyces nepalensis was studied by growing cells in shake flask on rotary shaker and cell growth was measured. Specific growth rate (μ) of D. nepalensis increased with increase in temperature from 15 to 35 °C and then decreased with increase in temperature beyond 35 °C when grown in presence and absence of different salts. At 40 °C, both NaCl and KCl at 1.0 M concentration enhanced the specific growth rate. D. nepalensis showed synergistic effect on thermal and salt stress when grown at 35-40 °C (0.5 M NaCl) and 30-40 °C (0.5 and 1.0 M KCl). The organism was able to revert its specific growth rate when temperature was shifted from 20 °C to its optimum temperature for growth (30 °C) only in the presence of salts, which was not observed in the absence of salts. However, the same phenomenon was not observed when the temperature was shifted from 40 to 30 °C. Decrease in activation energy was observed for growth at salt concentration beyond 0.5 M of NaCl and KCl.
PDF Abstract XML References Citation
How to cite this article
Sathyanarayana N. Gummadi and Sawan Kumar, 2008. Effects of Temperature and Salts on Growth of Halotolerant Debaryomyces nepalensis NCYC 3413. American Journal of Food Technology, 3: 354-360.
DOI: 10.3923/ajft.2008.354.360
URL: https://scialert.net/abstract/?doi=ajft.2008.354.360
DOI: 10.3923/ajft.2008.354.360
URL: https://scialert.net/abstract/?doi=ajft.2008.354.360
INTRODUCTION
High salt concentration is tolerated by a diversity of microorganisms. Halotolerant yeasts constitute a heterogeneous group of yeast belonging to different genera viz., Candida versatilis (Silva-Graça and Lucas, 2003), Rhodotorula mucilaginosa (Lahav et al., 2004), Saccharomyces rouxii and Debaryomyces hansenii (Norkrans, 1966; Onishi, 1963) and Pichia farinose (Suzuki, 1999). Among them, D. hansenii, isolated from marine water has been extensively studied for its halotolerant attributes to NaCl and KCl and the molecular and physiological basis for its salt tolerance has also been well elucidated (Prista et al., 1997). Studies have shown that NaCl and KCl have stimulatory effect on D. hansenii at 34 °C (Almagro et al., 2000). However, growth of this strain was inhibited in the absence of salts at 34 °C, thus establishing its halophilic nature.
Previously, we have isolated a yeast strain, Debaryomyces nepalensis from rotten apple, which is capable of utilizing pectin as the sole carbon source (Gummadi and Kumar, 2006). This strain was also tolerant to high concentrations of NaCl (up to 2.0 M) and KCl (up to 3.0 M), with the specific growth rate being higher than that of D. hansenii (Gummadi et al., 2007). Apart from that, D. nepalensis showed considerable growth in the presence of 0.5 M LiCl, which has been reported to be toxic to D. hansenii (Prista et al., 1997). Usually the combination of temperature, salt and pH are the important factors affecting the food spoilage and stability of food products (Papouskova and Sychrova, 2007). A detailed study and understanding about synergistic or antagonistic effects among these factors are required for economical food preservation. Hence, in this study, we examined the effect of different temperatures on the growth of D. nepalensis under different salt stress conditions.
MATERIALS AND METHODS
This study was conducted at Indian Institute of Technology-Madras during January-June, 2007.
Media
Debaryomyces nepalensis NCYC 3413 was maintained at 30 °C on YEPD (Yeast extract-Peptone-Dextrose) plates, which had the following composition (g L-1): Yeast extract, 10; peptone, 20; dextrose, 20 and agar 20. The pH of media was adjusted to 7.0 with NH4OH (100 mM) before sterilization.
Flask Culture Experiments
Flask culture experiments were performed in YEPD broth to study the effect of temperature on growth of halotolerant yeast, D. nepalensis in presence or absence of different salts viz., NaCl, KCl and LiCl. A loopful of the strain from YEPD plates was transferred to 50 mL sterile YEPD medium and incubated on rotary shaker (Techlab Incubator shaker, Techlab Instruments Co., Chennai) at 180 rpm for 12 h at 30 °C. When OD 600 reached ~ 1.0, 5% (v/v) of the culture was transferred to 50 mL of YEPD medium containing different concentration of NaCl (0, 0.5, 1.0 and 2.0 M), KCl (0, 0.5, 1.0 and 2.0 M) and LiCl (0, 0.5 and 1.0 M) and incubated on rotary shaker at 180 rpm at temperatures ranging between 15 to 40 °C. Samples were collected at regular intervals of time and analyzed for cell growth as a function of temperature at different concentration of NaCl, KCl and LiCl. The effect of temperature shifts (whether the cells can revert their growth when growth temperature was changed) on growth of D. nepalensis in medium without salt or with 1.0 M NaCl or 2.0 M KCl was studied. To accomplish this, cells were first grown at stress condition of low temperature (20 °C) or at high temperature (40 °C) and were subsequently shifted to 30 °C (optimum temperature for the growth) at the mid-log phase. Specific growth rates were calculated before and after temperature shift.
Calculation of Specific Growth Rate and Activation Energy
In order to obtain the unknown cell dry weight directly from the optical density readings, the calibration graph between optical density and the corresponding cell dry weight was established (OD600 of 1.0 corresponds to 0.33 g cell dry weight L-1). A plot of log (cell concentration at any time/initial cell concentration) versus time was plotted during the exponential growth phase. The specific growth rate was then calculated from the slope of the linear plot obtained. Activation energy for temperature dependent growth of D. nepalensis under salt stress condition was calculated using Arrhenius equation.
where, μ is the specific growth rate, Ea is the activation energy, R is the universal gas constant (8.314x10-3 kJ mol-1 K-1), T is the absolute temperature and A is a non-zero constant called Arrhenius constant. A graph of ln (specific growth rate) versus absolute temperature inverse was plotted. Slope of the plot gave the value of Ea/R from which the value of Ea was calculated. Similar plots were constructed for various salts concentrations to get the values of Ea for D. nepalensis when grown at different salt concentrations.
Statistical Analysis
All the experiments were performed twice in quadruplicates and values reported are mean of eight independent values with p<0.05 from F-test.
RESULTS AND DISCUSSION
A characteristic feature of D. nepalensis is its ability to grow in high salt concentration. Since, this yeast has been isolated from rotten apple, it has been considered to be involved in fruit spoilage. Hence, it is particularly important to understand the behavior of this yeast when a high salt concentration is present together with other stress conditions like extremes of temperature. Preliminary experiments revealed that minimum temperature for the growth of the organism was 8-10 °C and maximum was 40- 42 °C. Complete inhibition of growth was observed at 8 and 45 °C. Hence, the effect of temperature was studied from 15 to 40 °C under different salt concentrations for D. nepalensis. Specific growth rate (μ) of D. nepalensis increased with increase in temperature from 15 to 35 °C and then decreased with increase in temperature when grown in presence and absence of different salts (Fig. 1A-C). Maximum biomass growth was obtained when grown at 35 °C. In the case of NaCl, μ decreased with increase in salt concentration in the temperature range 15-35 °C. The growth rate was reduced by ~60% at 25, 30 and 35 °C as compared to control and was completely inhibited at 15 and 20 °C in medium supplemented with 2.0 M NaCl. At 35 °C, μ of D. nepalensis when grown at 0.5 M NaCl and in the absence of salt was found to be same (Fig. 1A). Further increase in salt concentration at 35 °C decreased the specific growth rate. Even though μ was lower at 40 °C than at 35 °C, μ was same for control (no salt) and at all concentrations of NaCl (Fig. 1A). But this phenomenon was not observed at 30 °C. This clearly showed that synergistic effect of thermal and salt stress exists in D. nepalensis between 35-40 °C and at 0.5 M NaCl.
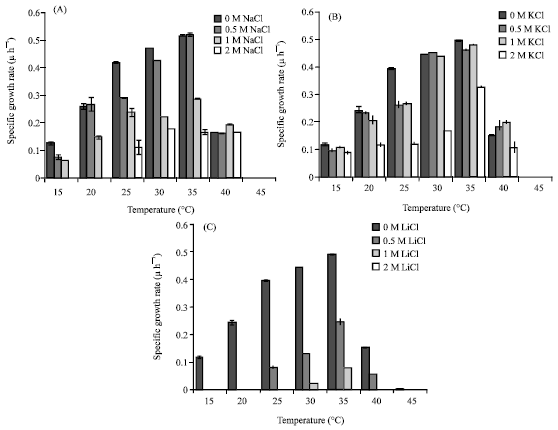 |
| Fig. 1: | Effect of temperature on specific growth rate of D. nepalensis under different concentration of salt: (A) for NaCl (B) for KCl and (C) for LiCl. The strain was grown at temperature ranging between 15 to 45 °C in medium containing required concentration of NaCl, KCl or LiCl and medium without salt was used as control. The data reported are mean of eight data points within ±5% standard error |
In the case of KCl, μ increased with increase in temperature for all the concentrations of KCl (Fig. 1B). Growth of isolate in the presence of KCl was higher than NaCl at all temperatures considered in this study. At 30 °C and 35 °C, specific growth rate was same for control and up to 1.0 M KCl but decreased at 2.0 M KCl (Fig. 1B). Similar observation was found for NaCl at 35 °C and up to 0.5 M NaCl (Fig. 1A). Interestingly at 40 °C, specific growth rate was enhanced in the presence of 0.5 and 1.0 M KCl than control (Fig. 1B). Similar to NaCl, the synergistic effect of thermal and salt stress in D. nepalensis exists between 35-40 °C and at 0.5 and 1.0 M KCl. At 15 and 20 °C, 2.0 M KCl was not inhibitory to growth as observed in case of 2.0 M NaCl. In the presence of 0.5 M LiCl, the isolate was unable to grow up to 20 °C and μ increased with increase in temperature from 25 to 35 °C (Fig. 1C). Similarly, μ increased with increase in temperature from 15-35 °C for control (in the absence of salt). In contrast to NaCl and KCl, synergistic effect of thermal and salt stress was not observed for LiCl in the concentration range considered in this study.
The growth of D. nepalensis at high cultivation temperatures is not only improved by presence of NaCl, but also by KCl. Thus, growth improvement in cells under high temperature stress is connected to the increased osmotic pressure of the media. The growth improvement in the presence of two non-related stress conditions (high temperature and increased osmotic pressure of the media due to high salt concentration) can be a manifestation of a phenomenon similar to stress cross protection. Stress cross protection describes the protection of yeast cells against a stress condition upon exposure to another, seemingly unrelated stress (Ruis, 1997). These observations have been explained by the environmental stress response in S. cerevisiae (Causton et al., 2001; Gasch et al., 2000). In C. albicans, the stress-activated protein kinase hog1 is known to play an important role in the environmental stress response (Enjalbert et al., 2006; Smith et al., 1995). This enzyme has also been found in D. hansenii (Bansal and Mondal, 2000; Sharma et al., 2005) hence making a positive contribution to stress response in D. nepalensis. Both the salts inhibited the growth of D. nepalensis at lower temperature. Presumably, low temperature and the presence of high salt concentration are antagonistic as for their effect on synthesis of appropriate lipids for adaptation to each of these situations. It has been shown that lipid saturation is important for salt tolerance in Zygosaccharomyces rouxii (Yoshikawa et al., 1995) while adaptation to low temperature results in an increase of degree of lipid unsaturation in several yeast species (Arthur and Watson, 1976). On the contrary, K+ being non-toxic enhanced the growth at high temperature up to 1.0 M concentration.
When temperature shift was made from 20 to 30 °C, it has been found that specific growth rate of D. nepalensis reduced from 0.25 (at 20 °C) to 0.16 (shifted to 30 °C) in the absence of salt. However, in the presence of 1.0 M NaCl, μ increased from 0.14 (at 20 °C) to 0.24 (which is greater than 0.18 when grown at only 30 °C) when temperature was shifted to 30 °C. Similar observation was found when temperature shift was made for D. nepalensis in the presence of 2.0 M KCl (Table 1). These results clearly showed that in the presence of salts, the strain can overcome the thermal stress or thermal shock. Increase in specific growth rate was observed when temperature shift was made from 20 to 30 °C in the presence of 1.0 M NaCl and 2.0 M KCl but not in the absence of any salts (Table 1). Growth at low temperature can cause derangement of regulatory processes.
| Table 1: | Effect of temperature shift (20 to 30 °C and 40 to 30 °C) on growth of D. nepalensis on YEPD medium with no salt or with 1 M NaCl or 2.0 M KCl |
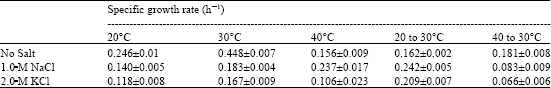 | |
Therefore, certain enzyme systems are repressed at low temperatures, thus resulting at lower enzyme level or activity at low temperature. As temperature increased from 20 to 30 °C, the enzymes involved in cellular metabolism is activated more, this in turn enhanced the specific growth rate (Daniel et al., 2008). Since the specific growth rates are higher than values obtained only at 30 °C in the presence of salts suggested that the temperature shock or temperature stress was overcome by salt stress.
Similarly, temperature shift from 40 to 30 °C was studied for cells grown in medium with and without salts. It has been found that specific growth rate of 0.18 h-1 was achieved after the temperature shift was made and this was intermediate between specific growth rates obtained at 30 and 40 °C (Table 1). A different situation for the temperature shift was noted when the medium was supplemented with salts. In medium supplemented with 1.0 M NaCl or 2.0 M KCl, decrease in specific growth rate was noticed for the strain when it was shifted from 40 to 30 °C, which is in contrast to shift in temperature from 20 °C to 30 °C (Table 1). Under this condition, probably enzymes involved in cellular metabolism are deactivated at 40 °C and they are not capable of regaining their normal activity when temperature was shifted to 30 °C. When cells growing at higher temperature, intuition would predict that higher the operating temperature, the faster the catalyzed reaction, but also the enzyme is less stable (Eisenthal et al., 2006). Hence, D. nepalensis are not capable of regaining their normal activity when temperature was shifted from 40 to 30 °C. These results clearly suggest that D. nepalensis exhibits different response to temperature shift at lower and higher temperatures.
Table 2 shows the activation energy (Ea) profile of D. nepalensis grown on medium with or without salt (NaCl and KCl) at different temperature ranging between 15-35 °C. The value of Ea was observed to be almost identical in absence or presence of salt (up to 0.5 M of NaCl and KCl). The average value was 58.5 kJ mol-1 and values ranged from 56.5 to 60.4 kJ mol-1. However, drastic decrease in activation energy (p<0.05) was observed for growth at salt concentration beyond 1.0 M of NaCl and KCl. Activation energy profile of growth of D. nepalensis in medium with and without salt at different temperature indicated that Na+ and K+ were non-toxic to its growth up to 0.5 M of KCl and NaCl. Above 0.5 M, activation energy decreased drastically, which in turn decreased growth rate values. This also suggest that growth of D. nepalensis is not much temperature sensitive up to 0.5 M concentration of NaCl and KCl, whereas growth is highly temperature sensitive at higher concentrations of salt. Similar results were interpreted for the microbial growth in packaged refrigerated beef at different temperatures (Giannuzzi et al., 1998). From these results, we conclude that synergistic effect of thermal and salt stress exists in D. nepalensis when grown in NaCl (0.5 M) and KCl (0.5 and 1.0 M) at 35-40 °C and 30-40 °C, respectively. In addition, D. nepalensis exhibits different response to thermal shock at lower and higher temperatures in the presence and absence of salts. The food industry is aware of the importance of the crossed effects of stress conditions in food preservation procedures. Present research describes the synergistic and antagonistic behavior of fruit spoilage yeast, D. nepalensis under condition when salt stress is present together with temperature stress. This will help establish fruit preservation procedures taking into account the resistance parameters of this yeast. Further studies on the molecular and physiological aspects such as morphology change and membrane lipid composition are being carried out.
| Table 2: | Activation energy (Ea) profile of D. nepalensis on medium with and without salt at different temperatures |
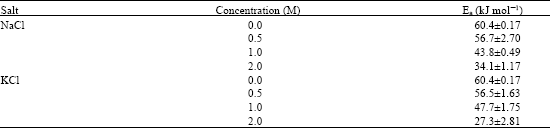 | |
ACKNOWLEDGMENTS
This research was supported by a grant from Department of Biotechnology, Government of India. Authors acknowledge Swati for critical reading of the manuscript.
REFERENCES
- Almagro, A., C. Prista, S. Castro, C. Quintas, A. Madeira, J. Ramos and M.C. Loureiro-Dias, 2000. Effects of salts on Debaryomyces hansenii and Saccharomyces cerevisiae under stress conditions. Int. J. Food Microbiol., 56: 191-197.
CrossRefPubMedDirect Link - Arthur, H. and K. Watson, 1976. Thermal adaptation in yeast: Growth temperatures, membrane lipid and cytochrome composition of psychrophilic, mesophilic and thermophilic yeasts. J. Bacteriol., 128: 56-68.
PubMed - Bansal, P.K. and A.K. Mondal, 2000. Isolation and sequence of the HOG1 homologue from Debaryomyces hansenii by complementation of the hog1 strain of Saccharomyces cerevisiae. Yeast, 16: 81-88.
CrossRefPubMedDirect Link - Causton, H.C., B. Ren, S.S. Koh, C.T. Harbison and E. Kanin et al., 2001. Remodeling of yeast genome expression in response to environmental changes. Mol. Biol. Cell, 12: 323-337.
PubMed - Eisenthal, R., M.E. Peterson, R.M. Daniel and M.J. Danson, 2006. The thermal behaviour of enzymes: Implications for biotechnology. Trends. Biotechnol., 24: 289-292.
CrossRefPubMedDirect Link - Enjalbert, B., D.A. Smith, M.J. Cornell, I. Alam, S. Nicholls, A.J. Brown and J. Quinn, 2006. Role of the Hog1 stress-activated protein kinase in the global transcriptional response to stress in the fungal pathogen Candida albicans. Mol. Biol. Cell, 17: 1018-1032.
PubMedDirect Link - Gasch, A.P., P.T. Spellman, C.M. Kao, O. Carmel-Harel and M.B. Eisen et al., 2000. Genomic expression programs in the response of yeast cells to environmental changes. Mol. Biol. Cell, 11: 4241-4257.
PubMedDirect Link - Giannuzzi, L., A. Pinotti and N. Zaritzky, 1998. Mathematical modeling of microbial growth in packaged refrigerated beef stored at different temperatures. Int. J. Food Microbiol., 39: 101-110.
CrossRefPubMedDirect Link - Gummadi, S.N. and D.S. Kumar, 2006. Pectin lyase and pectate lyase from Debaryomyces nepalensis isolated from apple. Res. J. Microbiol., 1: 152-159.
CrossRefDirect Link - Gummadi, S.N., S. Kumar and C.N.A. Aneesh, 2007. Effect of salts on growth and pectinase production by halotolerant yeast, Debaryomyces nepalensisNCYC 3413. Curr. Microbiol., 54: 472-476.
CrossRef - Lahav, R., A. Nejidat and A. Abeliovich, 2004. Alterations in protein synthesis and levels of heat shock 70 proteins in response to salt stress of the halotolerant yeast Rhodotorula mucilaginosa. Antonie Leeuwenhoek, 85: 259-269.
CrossRefPubMedDirect Link - Norkrans, B., 1966. Studies on marine occurring yeasts: Growth related to pH, NaCl concentration and temperature. Arch. Microbiol., 54: 374-392.
CrossRef - Papouskova, K. and H. Sychrova, 2007. The co-action of osmotic and high temperature stresses results in a growth improvement of Debaryomyces hansenii cells. Int. J. Food Microbiol., 118: 1-7.
PubMedDirect Link - Prista, C., A. Almagro, M.C. Loureiro-Dias and J. Ramos, 1997. Physiological basis for the high salt tolerance of Debaryomyces hansenii. Appl. Environ. Microbiol., 63: 4005-4009.
PubMed - Sharma, P., N. Meena, M. Aggarwal and A.K. Mondal, 2005. Debaryomyces hansenii, a highly osmo-tolerant and halo-tolerant yeast, maintains activated Dhog1p in the cytoplasm during its growth under severe osmotic stress. Curr. Genet., 48: 162-170.
CrossRefPubMedDirect Link - Silva-Graça, M. and C. Lucas, 2003. Physiological studies on long-term adaptation to salt stress in the extremely halotolerant yeast Candida versatilis CBS 4019. FEMS. Yeast Res., 3: 247-260.
CrossRefPubMedDirect Link - Smith, B.A., D.H. Oday and G.A. Proteau, 1995. Lithium in the matting response and cell-cycle of Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun., 206: 401-407.
CrossRef - Yoshikawa, S., N. Mitsui, K.I. Chikara, H. Hashimoto, M. Shimosaka and M. Okazaki, 1995. Effect of salt stress on plasma membrane permeability and lipid saturation in the salt-tolerant yeast Zygosaccharomyces rouxii. J. Ferment. Bioeng., 80: 131-135.
CrossRef