
Manal A. Hamed
Department of Therapeutic Chemistry, National Research Center,Dokki, Cairo, Egypt
Asian Journal of Epidemiology
Year: 2010 | Volume: 3 | Issue: 3 | Page No.: 123-140







ABSTRACT
Schistosomiasis is a snail-borne trematode infection of humans, domestic and wild animals in different parts of Asia, Africa, the Middle East, South America and the Carribbean. Approximately 200 million people in 74 countries are affected; 120 million of these are symptomatic and 20 million have severe disease. Elimination of schistosomiasis has been mainly accomplished by control of the snail host. As measures of snail control, cement-lining of ditches and chemical mollusciciding were most effective in many countries. But the cost of this joint program is too expensive compared with health budget in almost developing countries. Due to persisting conditions of poor health infrastructure, lack of access to clean water and poverty, re-infections in humans still poses a challenge for the long-term control of schistosomiasis. It is hoped that vaccines and better diagnosis of human will help alleviate some of these challenges. However, until these become available, alternative strategies, including blocking parasite transmission in the snail host have been considered. Several studies have been conducted in recent years to begin to understand the molecular basis of the snail-parasite interaction and to identify genes that may be involved in rendering snails resistant to infection.
PDF Abstract XML References Citation
Received: January 28, 2010;
Accepted: March 13, 2010;
Published: June 05, 2010
How to cite this article
Manal A. Hamed, 2010. Strategic Control of Schistosome Intermediate Host. Asian Journal of Epidemiology, 3: 123-140.
DOI: 10.3923/aje.2010.123.140
URL: https://scialert.net/abstract/?doi=aje.2010.123.140
DOI: 10.3923/aje.2010.123.140
URL: https://scialert.net/abstract/?doi=aje.2010.123.140
INTRODUCTION
Schistosomiasis is considered the second most pre- valiant world wide parasitic disease ranked next to malaria. It has significant economic and public health consequences in many developing countries (Engels et al., 2002). Egypt is one of the most highly endemic areas in the world with infection rates exceeding 80% (5-6 million) in some localities in the Nile valley (El-Khoby et al., 1998).
Various human-pathogenic species of schistosomes are known of, which are dependent on various intermediate hosts and therefore occur in various regions of the world. S. haematobium causes urinary schistosomiasis and is the most prevalent and widespread species in Africa, Eastern Mediterranean and the Middle East. The other four species cause intestinal schistosomiasis; S. intercalatum occurs in 10 countries in the rainforest belt of Africa; S. mansoni is found in over 52 countries in Africa, Caribbean, Eastern Mediterranean, Latin America; S. japonicum and S. mekongi are prevalent in Africa and the Pacific region (Utzinger et al., 2001) (Table 1).
Biomphalaria alexandrina snail as specific intermediate host of Schistosoma mansoni are prevalent in both upper and lower Egypt, but during the last decade, it became the most dominant species in the Nile Delta forming a main threat for schistosomiasis transmission in the North of Egypt. This snail species invades the irrigation areas and drainage systems and also water sources in reclaimed areas leading to infection in previously uninfected populations which eventually leads to increase schistosomiasis transmission in Egypt (World Health Organization, 2002).
| Table 1: | Various human-pathogenic species of schistosomes |
 | |
Schistosome Life Cycle
The life cycle of Schistosoma mansoni provides an example for all species of schistosomes. After the eggs of the human-dwelling parasite are emitted in the feces into the water, the ripe miracidium hatches out of the egg. The miracidium searches for a suitable fresh water snail to act as an intermediate host and penetrates it. Following this, the parasite develops via a so-called mother-sporocyst and daughter-sporocyst generation to the cercaria. The purpose of the growth in the snail is the numerical multiplication of the parasite. From a single miracidium results of a few thousand cercaria, every one of which is capable of infecting man. The cercariae propel themselves in water with the aid of their bifurcated tail and actively seek out their final host. When the recognize human skin, they penetrate it within a very short time. Following a migration through the body within the bloodstream, if they meet a partner of the opposite sex, they develop into sexually mature adults, laying eggs and complete its life cycle (Ghandour, 1978) (Fig. 1).
Disease Complications
The ova reaching the liver initiating Schistosoma granulomata in variable numbers as well as pathological changes in the liver ranging from early to advanced stage of fibrosis, depending on the degree of ova deposition (Aly and Hamed, 2006; Hamed, 2006). Worm embolism in the intrahepatic portal veins has a role in hepatic fibrosis, portal vein obstruction occurs with continuing infection and fibrogranulomatus reaction due to ova deposition, lead to massive schistosomal fibrosis and blockage of venules by numerous ova either with or without thrombosis (Silva et al., 2003) (Fig. 2).
So, schistosomiasis might pass from the isolated granulomata to hepatomegaly due to wide-spread granulomatous cellular infiltration of the portal tracts where ascites and oesophagogastric varices could be demonstrated in advanced cases of Schistosoma fibrosis (Njenga et al., 1998). Patients with both schistosomal and hepatitis C virus induced highly significant degree of fibrosis due to the addition of schistosomal hepatic periportal fibrosis (Amin et al., 1999).
Control of Schistosomiasis
Several ways have been practiced in order to bring the disease under an adequate control through the breakage of the life cycle of the parasite (Fig. 1).
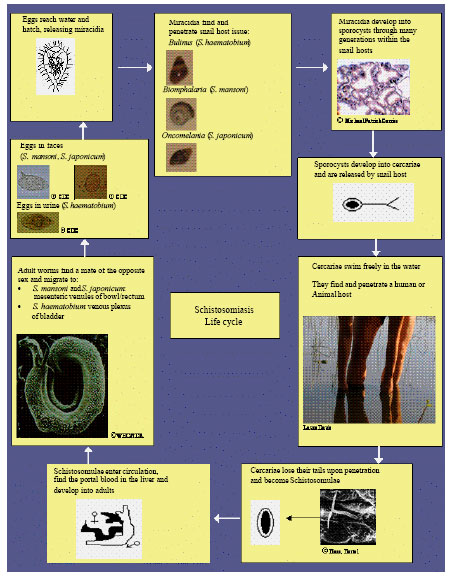 | |
| Fig. 1: | Schistosomiasis life cycle |
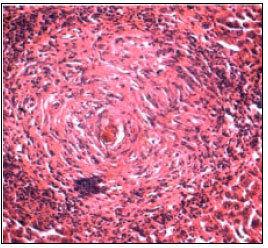 | |
| Fig. 2: | Haematoxylin and eosin liver stained section of S. mansoni infected mice liver showing large granulomata size with high infiltration area (200X) |
CONTROL THROUGH THE MAIN HOST
Health Education
Key to long term control of schistosomiasis are improvements in hygiene and sanitation. By eliminating human waste in fresh water bodies, part of the complex life cycle of the schistosome can be eliminated (Brinkmann and Steingruber, 1986). High rates of reinfection demonstrated the need for health education through programs established by the government.
Immunological Control
Generating immunity through the use of vaccines is complex. In the presence of high prevalence, vaccine would not be given to naïve patients. Rather, those receiving the vaccine can be expected to have already been exposed and to experience repeated exposure to schistosomiasis after getting the vaccine. It is precisely the host immune response that gives rise to the granulomas responsible for the morbidity of schistosomiasis. Potentially, by triggering the production of immunity to various schistosomiasis antigens, the vaccine could promote the production of granuloma formation. In fact, however, progress is being made in phase 1 and 2 clinical trials of different vaccines. Even without eradication of schistosomes from the environment, the vaccine appears to reduce susceptibility to re-infection. It is postulated that the vaccine’s artificially-induced immunity is boosted by re-exposure to the not-yet-eradicated schistosomes. This suggests that immunogenicity may need to be assessed if and when schistosomes are eliminated (World Health Organization, 2002). Many trials of vaccinations are based on homologous or heterologous antigens. Bashtar et al. (2006) found that schistosomal worm and egg antigen had a potency role in protection against schistomiasis, while Hamed (2006) postulated the immunization against schistosomiasis by using the excretory-secretory product of Fasciola hepatica worms.
Chemotherapy
Praziquantel (PZQ), a pyrazinoisoquinoline derivative, is the mainstay of treatment and a critical part of community-based Schistosomiasis Control Programs. Recent reports on praziquantel elucidate its fail to stop reinfection as a result of development of drug resistant Schistosoma strain (Silva et al., 2003), beside it induces hemorrhages in the lung tissue of the host (Flisser and McLaren, 1989) as well as abdominal pain and diarrhea by long term application of the drug (Kabatereine et al., 2003).
Mirazid, the oleo-resin extract from Myrrh of Commiphora molmol tree, was established in Egypt as a new drug against schistosomiasis and other parasitic diseases (Hamed and Hetta, 2005; Aly and Aly, 2006).
Hamed et al. (2004) mentioned that the methanolic extracts of Pulicaria crispa and Citherexylum quadrangular Jacq. were used as prophylactic treatments in S. mansoni infected mice through improvement of certain liver enzymes representing different metabolic pathways.
CONTROL THROUGH THE INTERMEDIATE HOST
Snail control could be regarded as a rapid and efficient of reducing or eliminating transmission and remains among the methods of choice for schistosomiasis control. The importance of snail control should be overlooked, for despite greatly improved chemotherapy by single dose oral treatment with praziquantel, there are logistical problems in mass treatment that threat of reinfection (Wilkins, 1989) and absence of a completely and safe schistosomiasis vaccine support the use of snail control as an important means in control programs (Butterworth, 1992).
Snail control can be divided into three categories; environmental, molluscicides and biological control.
Environmental Control
Environmental factors which influence snail distribution and which might be manipulated to be achieved were suggested by Thomas and Tait (1984). These factors are classified as follows:
Water Chemistry
Water chemistry includes calcium concentration, total dissolved chemical content and oxygen. The densities of all snail species were very low in salt water, high in medium water and somewhat lower in hard water, where low calcium concentration appears to be the direct or indirect cause of the poor snail fauna (Utzinger et al., 1997).
The effect of NaCl concentration and other aspects of salinity on freshwater pulmonate snails were reviewed by Madsen (1990). It appears that high salinity will rarely prevent the establishment of intermediate hosts for schistosomes in water, where the free living stages of the parasite are likely to survive.
The level of oxygenation may be an important influence on distribution within habitats. There is a zone of high concentration of oxygen available to snails immediately beneath the floating leaves of, however dense floating vegetation preventing snail from reaching the surface where there is a shortage of dissolved oxygen (Brown, 1994).
Temperature
The power of temperature to limit distribution was simply demonstrated by Pitchford (1981) who observed that Biomphalaria snails were killed during the winter when night temperature fell gradually below freezing.
Coelho and Bezerra (2006) studied the influence of temperature on the development of Schistosoma mansoni infections in Biomphalaria glabrata. The results show a direct relationship between temperature and infection rate, i.e., the lower temperature the lowest is the infection level of B. glabrata with S. mansoni. The snails were infected at 15, 20 and 30°C and the cercarial release was recorded after 30 and 60 days post-infection.
Yang et al. (2007) found a positive relationship between the development of S. japonicum within Oncomelania hupensis and temperature. In snails kept at 15.3°C, S. japonicum arrested their development, while the fastest development occurred at 30°C. The temperature at which half of the snails were in hibernation was 6.4°C.
Current Speed
Some snails are adapted to fast-flowing water and have the shell modified to resist dislodgement, while in small rivers and streams that are slowly flowing or stagnant for most of the year the sudden spates following heavy rainfall sweep away many snails and cause major fluctuations in population density. So, the water current may be an important means for the dispersal of snails. The adverse effect of strong current on snail populations was attributed also to sweeping away of food and stress caused to the snails (Marti, 1986).
Light and Shade, Circadian Rhythms
Snail hosts for schistosomes showed a regular pattern of activity in relation to the 24 h cycle of day and night (circadian rhythms) (El-Emam and Madsen, 1982).
In assessing the effect of solar radiation on snails in the field it is difficult to separate the effects of light and temperature. Shaded sites are unfavorable for B. pfeifferi and shading by trees is suggested as a mean for controlling this snail. Dense shade beneath mats of floating vegetation is generally unsuitable for snails. The adverse effect of shade is thought to be indirect due to depression of the growth of sub-aquatic vegetation that provides snails with food and oxygenates the water. Yet some species such as Lymnaea libycus and Physa waterloti may found most frequently in high shaded places (Ndifon and Ukoli, 1989).
Snail-Plant Association
The greatest species diversities of freshwater snails are usually associated with aquatic or sub-aquatic (emergent) leafy plants (Macrophytes). There may be a symbiotic relationship between snails and aquatic macrophytes, evolved over a long period. Plants provide snails with shelter from solar radiation and the water current, sources of food and egg-laying sites (Thomas, 1987).
Most aquatic habitats contain rich microflora which, together with decaying vegetable matter, provide the principal food of snails, although there may be no preference for any particular species of microflora (Madsen, 1992).
Control by Molluscicides
Chemical Molluscicides
Molluscicides consider as an effective mean for control of snail populations and it has played an important role in schistosomiasis control. The use of molluscicides as one of the strategies to control schistosomiasis began by using niclosamide, an ethanolamine salt of 2', 5-dichloro-4'-nitrosalicylanilide, manufactured under the trade name Bayluscide, whose efficacy had previously been established. Application of this product caused biocidal action on non-target plants and animals, besides causing genotoxicity and carcinogenic effects. The high cost, the possibility of recolonization of breeding grounds and the ecological toxicity of this product were limitations on its use as an official molluscicide program of schistosomiasis control (Mello-Silva et al., 2006).
Contradictory, Takougang et al. (2006) studied the effect of lower Bayluscide doses on snail host and non-target fish, frog and tadpole. Bayluscide concentration of 0.50 g m-3 applied to ponds under investigation resulted high snail mortality and show low lethality to fish, frogs and tadpoles.
Zhu et al. (1998) and Lima et al. (2002) postulated the effect of bromoacetamide and potassium salts of isolapachol and lapachol as safe and effective molluscicides against adult snails of Biomphalaria glabrata and Oncomelania species as well as snail egg masses.
Some N-p-substituted phenyl uracil-5-sulphonamide derivatives have been synthesized to be evaluated as molluscicides against Biomphalanaria alexandrina snails and also its role as immunostimulatory effect on S. mansoni infected mice. The selection of the concentration based on the predetermined dose which caused mortality of less than 50% of snails/24 h. Therefore, treatment of hemolymph obtained from pre-treated snails with these derivatives can stimulate specific immune response and induce protective effects against S. mansoni infection by 44-50% reduction in worm burden (Fathalla et al., 2003).
Thymol, Linalool and Eugenol (monoterpenes) showed considerable molluscicidal effect against Biomphalaria alexandrina, Bulinus truncatus and Lymnneae natalensis, where a significant decrease in succinate dehydrogenase together with a concomitant increase in glucose-6-phosphate dehydrogenase, acid phosphatase and alkaline phosphatase activity levels were investigated. Treated snails also showed an elevation in the hemolymph glucose content, while the tissue glycogen content was reduced. The infection of B. alexandrina with S. mansoni miracidia was greatly reduced by thymol LC10. The infection rate reduction was 43.1%. The treated snails’ prepatent period was prolonged (34.2±3.3 days) compared to control (28.4±1.2 days) and a highly significant reduction of total cercarial production per snail was also observed (El-Din, 2006).
Fenitrothion and anilofos (aniloguard) were tested as molluscicides against Lymnaea natalensis and Biomplhalaria alexandrina. The results obtained showed that sublethal concentrations of fenitrothion caused reduction in growth rate of B. alexandrina and reduction in the hatchibility of snails eggs. The mortality rates of miracidia and cercariae were elevated by increasing both the concentrations of fenitrothion and the time of exposure. Also, total protein, alkaline phosphatase, alanine and aspartate aminotransferases enzyme activities were markedly disturbed in treated snails (Tantawy, 2006).
Saad and Sayed (2000) postulated that caprylic acid, margaric acid and lenoleic acid, are three fatty acids which could be chemoattractive of Schistosoma miracidium and could be used as safe control compounds, which needs further research.
Mostafa (2006) tested three oils for their molluscicidal activity, Caple-2, Kemasol and Super-max. Super-max had the strongest toxic effect on B. alexandrina and other non target snail species, where the hatchability of snails’ eggs was stopped completely and 100% mortality of miracidia. The infection rate of B. alexandrina with S. mansoni miracidia was greatly reduced, a highly significant reduction of total cercarial production per snail, decrease of total protein content and increase of alkaline phosphatase, alanine and aspartate aminotransferases enzyme activities were also demonstrated.
Plant Molluscicides
The high cost of synthetic molluscicides used in the control of the intermediate snail hosts of schistosomiasis, along with increasing concern over the possible built up of snail resistance of these molluscicides and their toxicity to non-target organisms, has drawn much attention during recent years in renewed interest in the use of plant molluscicides (El-Ansary et al., 2001a; Mantawy and Mahmoud, 2002). These plant molluscicides may provide cheap, locally produced, biodegradable and effective control agents in rural areas of developing countries where schistosomiasis is endemic (Clark et al., 1997). The same authors suggested that plant molluscicides could be used in low doses at transmission foci to reduce schistosome in infected snails and recommended the use of sublethal concentrations. El-Ansary et al. (2000a, b, 2001a, b) recorded that sublethal concentrations of selected plant molluscicides were effective in reducing the compatibility of B. alexandrina snails to S. mansoni infection as seen through reduction in cercarial shedding and elongation of the prepatent periods.
Molluscicidal potency of many plants as Ambrosia maritime, Solanum nigrum, Thymelaea hirsuta, Callistemon lanceolatus and Peganum harmala were previously studied by Ahmed and Ramzy (1997), El-Ansary et al. ( 2000a, b, 2001a, b). They attributed the molluscicidal effect of these plants to the disturbance occurs in glycolytic pathways. They reported that reduction of snail compatibility for the developing parasite was due to the disturbance of hexokinase, glucose phosphate isomerase and pyruvate kinase as three glycolytic enzymes. Moreover, they declared that glycolysis is the most important metabolic pathway for infected snails which should be targeted by synthetic or plant molluscicides.
Rug and Ruppel (2000) studied the toxic activity of methanolic extract of Jatropha curcas L. (Euphorbiaceae) against snails transmitting Schistosoma mansoni and S. haematobium. It showed the highest toxicity with LC100-values of 25 ppm for Biomphalaria glabrata and 1 ppm for Bulinus truncatus and B. natalensis. Dodonaea viscosa and Haplophyllum tuberculatum herbs also showed molluscicide potency through marked alteration in AMP, ADP, ATP and adenylate energy charge of B.alexandrina snails (El-Ansary et al., 2001a).
Mantawy and Mahmoud (2002) added that, Allium cepa (onion) and Allium sativum (garlic) have molluscicidal effect through disturbance in the protein profile, glucose and glycogen content of B.alexandrina snails.
Dry powder of Capparis spinosa and Acacia arabica plant leaves seem to have a molluscicides potency against Biomphalaria alexandrina snails through disturbance in glycolytic and gluconeogenic pathways as well as protein, glucose and glycogen content (Aly et al., 2004; Mantawy et al., 2004).
Truiti et al. (2005) stated that the aerial parts of Melochia arenosa plant was 100% lethal to Biomphalaria glabrata snails at 200 μg mL-1 and showed LD50 of 143 μg mL-1.
El-Sayed (2006) stated that treatment of Biomphalaria alexandrina snails with the dry powder of the plant aerial part; Cupressus macro-carpa (Cupressacea) was significantly reduced pyruvate kinase, lactate dehydrogenase, hexokinase and phosphoenol pyruvate carboxykinase which are very important in metabolism of the protein and carbohydrate in both haemolymph and tissue of Biomphalaria alexandrina snail.
Mello-Silva et al. (2006) considered that the latex of Euphorbia splendens var. hislopii is the most promising plant molluscicides because it meets the recommendations of the World Health Organization (WHO, 2002). The researchers found that 0.6 mg L-1 of the latex of Euphorbia splendens var. hislopii causes a sharp reduction in the reserves of glycogen in the digestive gland and elevation of the protein content in the hemolymph of B. glabrata.
Commiphora molmol (Myrrh) has molluscicidal effect on Biomphalaria snails, where the number of dead-snails increased with increasing the time of exposure. One day-old egg masses were more susceptible to the ovicidal effect of C. molmol than the five-day old ones, hence the embryogenesis began to stop and the fecundity decreased. Based on safety to man and animals, C. molmol is recommended as a safe molluscidide (Massoud et al., 2004; Al-Mathal and Fouad, 2006).
Dos Santos et al. (2007) evaluated the latex of Euphorbia conspicua (Euphorbiaceae) for its molluscicidal and cercaricidal activities. It exhibited high activities against adult snails with LC90 values of 4.87 μg mL-1 and showed a lethal effect to the cercaria of Schistosoma mansoni at concentrations of 100 μg mL-1.
Biological Control
Snails and Different Organisms' Relationship
Biological methods for the control of fresh water snails were reviewed by Madsen (1992), who concluded that more emphasis should be put on searching for pathogens or microparasites as agents for control that can affect directly or indirectly on the intermediate snail hosts.
Fishes as Trematocranus placodon are employed in the biological control of schistosome intermediate host, where snails are consider as its preferred food (Evers et al., 2006). In addition, Kloos et al. (2004) recommended the possible use of tilapia fish for biological control of Biomphalaria in fishponds as well as modeling of S. mansoni transmission and re-infection.
Muschovy ducks (Cairina maschata) seem also to be predators of snail populations (Ndlela and Chimbari, 2000).
Host Parasite Relationship
The interaction of Schistosoma mansoni and the intermediate host has been the object of several studies. These studies show that the innate defense system is basically composed of phagocitary cells named hemocytes and their soluble products. Parasite recognition and hemocyte activation are mainly mediated by lectins. Besides lectins, the hemocytes in snails also produce some proteins, which are similar to mammal cytokines, such as TNF-α, that is depleted in S. mansoni infections and IL-1 "like", that was also found in snails and is associated to the activation and cellular proliferation, as well as to the increase of phagocitary activity of hemocytes and to the production of super-oxydes. This defense system play a major role in the resistance of different snails to infection, however there are no strain completely resistant to S. mansoni infection, the snail strain which was reported as resistant to S. mansoni, posteriorly proved to be susceptible to this parasite, when juvenile specimens were used (Coelho et al., 2004).
Grassi et al. (2001) attributed the resistance of B. straminea to S. mansoni miracidia to an efficient defense system that destroys miracidia once they have penetrated, where 94% of the penetrating miracidia appeared encapsulated by the B. straminea defense system.
Smit et al. (2004) found a novel internal defense peptide of the snail Lymnaea stagnalis which increases upon infection with the avian schistosome Trichobilharzia ocellata. This protein, named granularin, is secreted by granular cells. The protein is unique because it comprises only a single Von Willebrand factor type C domain that is normally found in large transmembrane and secreted extracellular matrix proteins. The granularin gene is twice up-regulated during parasitation. Hence, purified granularin stimulates phagocytosis of foreign particles by blood hemocytes which indicate that granularin represents a novel protein that acts as an opsonin in the molluscan internal defense response.
Khayath et al. (2006) demonstrated the possible central role of glutamine in mollusc-schistosome interactions, where it is massively expressed in larval forms as compared to adult parasites, suggested that glutamine could also be used for glucose or glycerol production, hence several hypotheses can be proposed concerning the importance of glycerol for the adaptation of this helminth to its host osmotic and energetic environment.
Blair and Webster (2007) characterized the impact of dose-dependent schistosome exposure and/or infection establishment on intermediate host survival and reproduction of Biomphalaria glabrata snails exposed to increasing doses of Schistosoma mansoni parasites. Increased mortality was observed amongst both snails infected and also those snails exposed to the parasite but within which infection did not establish. Snails also facultatively altered their reproductive output in response to parasite exposure, where the egg mass production decreased with increasing parasite dose in patently infected snails. These results uniquely suggest an exposure-dose-dependent post-patent fecundity compensation occurring in relation to the risk of future parasite-associated mortality.
Lehr et al. (2007) postulated the concept that the parasite synthesizes a wide array of glycoconjugates, exhibiting, in part, unique carbohydrate structures and expresses definitive host-like sugar epitopes, such as Lewisxdeterminants, supporting the concept of carbohydrate-mediated molecular mimicry as an invasion and survival strategy. The results demonstrated the presence of common carbohydrate epitopes at the level of S. mansoni and its intermediate host Biomphalaria glabrata.
Genetic Control
The concept of snail control on genetic basis has gained a considerable interest, to bring this hazardous disease under an adequate control, since snail control is one of the most rapid and effective means available for reducing transmission of schistosomiasis. The objective was to change high susceptible strains to non-susceptible state through the release of resistance snails into natural habitats as reported by Joubert et al. (1991). This approach however, requires a more thorough understanding of the complex interrelationship between parasites and snails (Spada et al., 2002; Coelho et al., 2004; Da Silva et al., 2004).
Since, the development of the Schistosoma parasite in the intermediate host snail is influenced by a number of parasite and snail genes as reported by Rollinson et al. (1998), genetic control of the snails plays an important role in schistosomiasis control. A growing interest revolves around identifying the products of the snail and parasite genes influencing these associations. Previous studies have demonstrated the great variability in the suitability of different snail genera and species to act as carriers for S. mansoni species (Da Silva et al., 2004; Lotfy et al., 2005).
The detection of specific DNA sequences by Polymerase Chain Reaction (PCR) has proved extremely valuable for the analysis of genetic disorders and the diagnosis of a variety of infectious disease pathogens especially that the recommendation of the World Health Organization is to focus the researches of schistosomiasis on the development and evaluation of new strategies and tools for control of the disease, hence recent studies describe sensitive and specific PCR systems to detect S. mansoni, indicating possible applications in the detection of snail infection, monitoring of transmission sites and diagnosis of human infection (Lardans and Dissous, 1998; Abath et al., 2006).
Due to difficulties of morphological identification of Biomphalaria species, five Biomphalaria populations from the Colombian Amazon region were identified and characterized by polymerase chain reaction-restriction fragment length polymorphism directed at the internal transcribed spacer region of the rRNA gene. These species known as; B. straminea, B. peregrina, B. kuhniana, B. intermedia and B. amazonica (Velasquez et al., 2002).
Caldeira et al. (2004) detected DNA from traces of organic material from inside shells, which found empty in the filed, in order to identify molluscs through polymerase chain reaction and restriction fragment length polymorphism and to detect S. mansoni into these snails as well as the infection rate in B. tenagophila, B. stramina and B. glabrata.
Lotfy et al. (2005) used PCR technique for survey of Biomphalaria glabrata presence in Egypt. The authors found no evidence for B. glabrata in Egypt, so if it is present, it is uncommon, while Biomphalaria alexandrina remain common and no evidence for hybridization with B. glabrata was found.
A simple and single-step technique based on multiplex PCR (multiplex polymerase chain reaction) has been developed for simultaneous identification of Brazilian Biomphalaria species, the intermediate hosts of Schistosoma mansoni and their diagnosis of infection by the trematode. The species-specific primers directed both to the internal transcribed spacer 2 of ribosomal DNA from 3 of the S. mansoni host species and to the mitochondrial DNA from the trematode, revealing the presence of specific bands efficient for identification of Biomphalaria species and diagnosis of snails infected by S. mansoni during prepatent periods (Jannotti-Passos et al., 2006).
Chen et al. (2006) established a sensitive and specific PCR assay for detecting Schistosoma japonicum-infected Oncomelania hupensis, based on 18S-rRNA gene of S. japonicum. They found the location of PCR product of detecting Oncomelania snails infected with S. japonicum was similar to the target DNA, with a length of 469 base pair and the same sequence as the target DNA.
Polymerase Chain Reaction (PCR) technique was also established to determine the state of susceptibility and resistance of snails to infection. Lockyer et al. (2004) studied the changes in gene expression in response to parasite infection in a susceptible and a resistant strain. Ten transcripts were initially identified, present only in the profiles derived from snails of the resistant strain when exposed to infection. The differential expression of five of these genes, including several novel transcripts with one containing at least two globin-like domains, has been confirmed by semi-quantitative RT-PCR.
Theron and Coustau (2005) found that the susceptibility and resistance of Biomphalaria glabrata snails to S. mansoni infection does not depend on the snail susceptibility/resistance status, but on the matched or mismatched status of the host and parasite phenotypes.
Rosa et al. (2005) observed that the Taim population of B. tenagophila is dominant resistant and presents a molecular marker of 350 base pair, a missed band from juvenile of B. tenagophila which is highly susceptible. Barbosa et al. (2006) added that transplantation of the haematopoietic organ from Biomphalaria tenagophila (Taim strain), resistant to Schistosoma mansoni, to a highly susceptible strain (Cabo Frio strain) of the same species, showed in the recipient snails resistance against the trematode, when a successful transplant occurred. The success of transplantation could be confirmed by a typical molecular marker of the Taim strain in haemocytes of the recipients of 350 base pair.
Abdel-Hamid et al. (2006) postulated that there is a genetic variations between susceptible and resistant strains to Schistosoma infection within B. alexandrina snails using random amplified polymorphic DNA analysis technique, where in the resistant genotype snails, OPA-02 primer produced a major low molecular weight marker of 430 base pair.
Lockyer et al. (2007) identified transcripts involved in Biomphalaria glabrata snail-schistosome interactions. Subtractive cDNA libraries were prepared, using suppression subtractive hybridization between a parasite-exposed schistosome-resistant and a susceptible strain of B. glabrata and also between schistosome-exposed and unexposed snails from the resistant snail line. Eight genes differentially expressed between the haemocytes of resistant and susceptible snail strains were identified and confirmed with reverse transcriptase PCR, including two transcripts expected to be involved in the stress response mechanism for regulating the damaging oxidative burst pathways involved in cytotoxic killing of the parasite. These regulatory machineries include; the iron storage- immunoregulatory molecule; ferritin and a serine protease involved in the cellular stress response. Transcripts with elevated levels in the resistant strain had the same expression patterns; higher levels in exposed resistant snails compared to susceptible ones and down-regulated in exposed compared with unexposed resistant snails.
Several studies have been done for determining the susceptibility and resistance of Schistosoma snails to infection according to the metabolic disorders occur in gastropods using PCR technique. This is in accordance to Hahn et al. (2001), who demonstrated the defensive role of reactive nitrogen species in interactions between hemocytes derived from the resistant 13-16-R1 strain of B. glabrata and Schistosoma mansoni parasite. The researchers suggested that NO and H2O2 are both involved in hemocyte-mediated toxicity of B. glabrata against S. mansoni sporocysts.
Bender et al. (2005) studied the production level of reactive oxygen species by hemocytes from the gastropod Biomphalaria glabrata and their ability to kill the trematode parasite Schistosoma mansoni. The findings suggest that the capacity to produce hydrogen peroxide in resistant snails is more than susceptible one and could be critical in determining susceptibility or resistance to S. mansoni.
Lockyer et al. (2005) postulated the role of cytochrome p450s as a family of structurally related proteins, with diverse functions, including steroid synthesis and breakdown of toxins. Hence, isolation and amplification of cytochrome p450 gene was carried out using PCR in both totally resistant or susceptible snail lines when exposed to infection which, suggests ubiquitous expression. Therefore, this protein did not determine resistance or susceptibility of snail, but seems as metabolically important protein.
Goodall et al. (2006) identified alleles of the gene coding for cytosolic copper/zinc superoxide dismutase (SOD1), where resistance to the parasite was found to be significantly associated with one allele of the SOD1 gene while a separate SOD1 allele was significantly associated with susceptibility of B. glabrata to infection. Contradictory, the anti-oxidant enzyme manganese superoxide dismutase (MnSOD) did not determine resistance or susceptibility for parasites, but expression of MnSOD is consistent with its involvement in a stress response of B. glabrata to infection (Jung et al., 2005).
Humphries and Yoshino (2006) proposed that p38 mitogen-activated protein kinase play a role in B. glabrata immune signaling, which are known to be associated with stress and inflammatory signaling. In a comparative study, activated haemocyte p38 mitogen-activated protein kinase could also be detected using the anti-phosphorylated p38 antibody following cell treatment with anisomycin. The results suggesting fundamental differences in the role of p38 mitogen-activated protein kinase in signal transduction pathways between haemocytes and B. glabrata embryonic cells. Similar comparative studies, based on proteomics of either snail or parasite protein extracts are also beginning to reveal key molecules (such as mucin-like proteins from both parasite and snail) that may play a role in snail/schistosome compatibility (Ittiprasert et al., 2010). A stress response, manifested by the modulation of genes encoding the stress response protein such as heat shock protein 70 may also underlie the snail-host/parasite encounter. Ittiprasert et al. (2009, 2010) founded that stress-related genes, heat shock protein 70 and reverse transcriptase were dramatically induced early in susceptible snails, but not in resistant/non-susceptible ones.
Most studies aimed towards deciphering differences in gene regulation between resistant and susceptible snails during the snail/schistosome encounter have focused mainly on this relationship in adult, but not juvenile snails. Age dependent variability in B. glabrata susceptibility to S. mansoni has been well documented with results showing that juvenile snails (even within the same stock) are, in general, more vulnerable than their adult counterparts to infection (Ittiprasert et al., 2010).
In summary, the concept of snail control has gained a considerable interest being easier, cheaper, safer and more promising, since; there is a high degree of specificity of schistosomes, as well as of other trematodes, to their intermediate snail hosts. Despite abundant emerging molecular information, very little is known about which snail genes to specifically target to develop transmission-blocking strategies for the eventual goal of disease control. For more permanent control method of schistosomiasis, understanding of the host/parasite association is necessary, since the host-parasite relationship is complex and question remains concerning the susceptibility of snails to infection by respective trematodes and their specificity and suitability as hosts for continued parasite development. Understanding the genetics involved in the complex host/parasite relationship may lead to select actively resistant snails and mass culture them to increase the proportion of alleles for insusceptibility as a possible mean for biological control of schistosomiasis in natural population.
REFERENCES
- Abath, F.G., A.L. Gomes, F.L. Melo, C.S. Barbosa and R.P. Werkhuser, 2006. Molecular approaches for the detection of Schistosoma mansoni: Possible applications in the detection of snail infection, monitoring of transmission sites, and diagnosis of human infection. Mem. Inst. Oswaldo Cruz, 101: 145-148.
PubMed - Abdel-Hamid, Z.A., S.M. Rawi and A.F. Arafa, 2006. Identification of a genetic marker associated with the resistance to Schistosoma mansoni infection using random amplified polymorphic DNA analysis. Mem. Inst. Oswaldo Cruz, 101: 863-868.
PubMed - Al-Mathal, E.M. and M.A. Fouad, 2006. Effect of Commiphora molmol on adults, egg masses and egg-deposition of Biomphalaria arabica under laboratory conditions. J. Egypt Soc. Parasitol., 36: 305-314.
PubMed - Aly, H.F. and S.A. Aly, 2006. Essential role of Citrus reticulata and Mirazid in treatment of Schistosoma mansoni infected mice: Biochemical and parasitological studies. Pol. J. Food Nutr. Sci., 15: 461-467.
Direct Link - Aly, S.A., H.F. Aly, N. Saba-el-Rigal and E.M. Sammour, 2004. Induced changes in biochemical parameters of the molluscan tissues non-infected using two potent plants molluscicides. J. Egypt Soc. Parasitol., 34: 527-542.
PubMed - Barbosa, L., R.L. Caldeira, O.S. Carvalho, T.H. Vidigal, L.K. Jannotti-Passos and P.M. Coelho, 2006. Resistance to Schistosoma mansoni by transplantation of APO Biomphalaria tenagophila. Parasite Immunol., 28: 209-212.
PubMed - Bender, R.C., E.J. Broderick, C.P. Goodall and C.J. Bayne, 2005. Respiratory burst of Biomphalaria glabrata hemocytes: Schistosoma mansoni-resistant snails produce more extracellular H2O2 than susceptible snails. J. Parasitol., 91: 275-279.
PubMed - Blair, L. and J.P. Webster, 2007. Dose-dependent schistosome-induced mortality and morbidity risk elevates host reproductive effort. J. Evol. Biol., 20: 54-61.
PubMed - Caldeira, R.L., P. Jannotti, M.P. Lira and O.S. Carvalho, 2004. Diagnostic of Biomphalaria snails and Schistosoma mansoni: DNA obtained from traces of shell organic materials. Mem. Inst. Oswaldo Cruz, 99: 499-502.
Direct Link - Chen, J.H., L.Y.Wen, X.Z. Zhang, J.F. Zhang, L.L. Yu and L.D. Hong, 2006. Development of a PCR assay for detecting Schistosoma japonicum-infected Oncomelania hupensis. Zhongguo Ji Sheng Chong Xue Yu Ji Sheng Chong Bing Za Zhi., 24: 204-207.
PubMed - Clark, T.E., C.C. Appleton and S.E. Drewes, 1997. A semi-quantitative approach to the selection of appropriate candidate plant molluscicide: A South African application. J. Ethnopharmacol., 56: 1-13.
PubMed - Coelho, J.R. and F.S. Bezerra, 2006. The effects of temperature change on the infection rate of Biomphalaria glabrata with Schistosoma mansoni Mem. Inst. Oswaldo Cruz, 101: 223-224.
PubMed - Coelho, P.M., O.S. Carvalho, Z.A. Andrade, R.L. Martins-Sousa and F.M. Rosa et al., 2004. Biomphalaria tenagophila/Schistosoma mansoni interaction: Premises for a new approach to biological control of schistosomiasis. Mem. Inst. Oswaldo Cruz, 99: 109-111.
PubMed - Dos Santos, A.F., D.P. de Azevedo, C. dos Santos Mata Rda, D.L. de Mendonca and A.E. SantAna, 2007. The lethality of Euphorbia conspicua to adults of Biomphalaria glabrata, cercaria of Schistosoma mansoni and larvae of Artemia salina. Bioresour. Technol., 98: 135-139.
CrossRef - El-Ansary, A., S. El-Bardicy, M.S. Soliman and N. Zayed, 2000. Sublethal concentration of Ambrosia maritime (Damsissa) affecting compatibility of Biomphalaria alexandrina snails to infection with Schistosoma mansoni through disturbing the glycolytic pathway. J. Egypt Soc. Parasitol., 30: 809-819.
Direct Link - El-Ansary, A., K. El-Shaikh and M. El-Sherbini, 2000. SDS-PAGE-Separated tissue proteins of Biomphalaria alexandrina snails in the presence and absence of Schistosoma mansoni. Egypt J. Parasitol., 30: 125-136.
PubMed - El-Ansary, A., E.M. Sammour, M.S. Soliman and F.A. Gawish, 2001. In vivo, attenuation of schistosome cercarial development and disturbance of egg laying capacity in Biomphalaria alexandrina using sublethal concentrations of plant molluscicides. J. Egypt. Soc. Parasitol., 31: 657-669.
Direct Link - El-Din, A.T., 2006. Molluscicidal effect of three monoterpenes oils on schistosomiasis and fascioliasis vector snails in Egypt. J. Egypt Soc. Parasitol., 36: 599-612.
PubMed - El-Emam, M.A. and H. Madsen, 1982. The effect of temperature, darkness, starvation and various food types on growth, survival and reproduction of Helisoma duryi, Biomphalaria alexandrina and Bulinus truncatus. Hydrobiologia, 88: 265-275.
CrossRef - El-Khoby, T., N. Galal and A. Fenwick, 1998. The USAID government of Egypt schistosomiasis research project (SRP). Parasitol. Today, 14: 92-96.
CrossRef - El-Sayed, K.A., 2006. Effect of the plant Cupressus macro-carpa (Cupressacea) on some haematolo-gical and biochemical parameters of Biomphalaria alexandrina snails. J. Egypt Soc. Parasitol., 36: 911-924.
PubMed - Engels, D., L. Chitsulo, A. Montresor and L. Savioli, 2002. The global epidemiological situation of schistosomiasis and new approaches to control and research. Acta Tropica, 82: 139-146.
CrossRef - Fathalla, O.A., M.E. Haiba and A.S. Maghraby, 2003. Synthesis of new uracil-5-sulfonamide derivatives and immunostimulatory effect of a chemically modified hemolymph of Biomphalaria alexandrina on Schistosoma mansoni infected mice. Arch. Pharm. Res., 26: 358-366.
PubMed - Ghandour, A., 1978. The development of Schistosoma haematobium in the hamaster. Trop. Med. Parasitol., 72: 219-225.
PubMed - Goodall, C.P., R.C. Bender, J.K. Brooks and C.J. Bayne, 2006. Biomphalaria glabrata cytosolic copper/zinc superoxide dismutase (SOD1) gene: Association of SOD1 alleles with resistance/susceptibility to Schistosoma mansoni. Mol. Biochem. Parasitol., 147: 207-210.
PubMed - Grassi, L., M. Torres Jorda, Z. Andrade and S.M. Gonzalez Cappa, 2001. Short report: Schistosoma mansoni miracidia are killed by the defense system of an Argentine strain of Biomphalaria straminea. Am. J. Trop. Med. Hyg., 65: 290-292.
Direct Link - Hahn, U.K., R.C. Bender and C.J. Bayne, 2001. Involvement of nitric oxide in killing of Schistosoma mansoni sporocysts by hemocytes from resistant Biomphalaria glabrata. J. Parasitol., 87: 778-785.
Direct Link - Hamed, M.A. and M.A. Hetta, 2005. Efficacy of Citrus reticulata and Mirazid in treatment of Schistosoma mansoni. Mem. Inst. Oswaldo Cruz, 100: 771-778.
Direct Link - Humphries, J.E. and T.P. Yoshino, 2006. Schistosoma mansoni excretory-secretory products stimulate a p38 signalling pathway in Biomphalaria glabrata embryonic cells. Int. J. Parasitol., 36: 37-46.
CrossRef - Ittiprasert, W., R. Nene, A. Miller, N. Raghavan, F. Lewis, J. Hodgson and M. Knight, 2009. Schistosoma mansoni infection of juvenile Biomphalaria glabrata induces a differential stress response between resistant and susceptible snails. Exp. Parasitol., 123: 203-211.
PubMed - Ittiprasert, W., A. Miller, J. Myers, V. Nene, N. M. El-Sayed and M. Knight, 2010. Identification of immediate response genes dominantly expressed in juvenile resistant and susceptible Biomphalaria glabrata snails upon exposure to Schistosoma mansoni. Mol. Biochem. Parasitol., 169: 27-39.
Direct Link - Jannotti-Passos, L.K., K.G. Magalhaes, O.S. Carvalho and T.H. Vidigal, 2006. Multiplex PCR for both identification of Brazilian Biomphalaria species (Gastropoda: Planorbidae) and diagnosis of infection by Schistosoma mansoni (Trematoda: Schistosomatidae). J. Parasitol., 92: 401-403.
Direct Link - Joubert, P.H., S.J. Pretorius and F.J. krunger, 1991. Further studies on the susceptibility of Bulinus africans to infection with Schistosoma haematobium. Ann. Trop. Med. Parasitol., 85: 253-258.
PubMed - Jung, Y., T.S. Nowak, S.M. Zhang, L.A. Hertel, E.S. Loker and C.M. Adema, 2005. Manganese superoxide dismutase from Biomphalaria glabrata. J. Invertebr. Pathol., 90: 59-63.
CrossRef - Kabatereine, N.B., J. Kemijumbi, J.H. Ouma, R. F. Sturrock and A.E. Butterworth et al., 2003. Efficacy and side effect of praziquantel treatment in a highly endemic Schistosoma mansoni focus at Lack Albert, Uganda. Trans. R. Soc. Trop. Med. Hyg., 97: 599-603.
Direct Link - Khayath, N., G. Mithieux, C. Zitoun, C. Coustau, J. Vicogne, A.G. Tielens and C. Dissous, 2006. Glyceroneogenesis: An unexpected metabolic pathway for glutamine in Schistosoma mansoni sporocysts. Mol. Biochem. Parasitol., 147: 145-153.
CrossRef - Kloos, H., L.K. Passos, P. LoVerde, R.C. Oliveira and A. Gazzinelli, 2004. Distribution and Schistosoma mansoni infection of Biomphalaria glabrata in different habitats in a rural area in the Jequitinhonha Valley, Minas Gerais, Brazil: Environmental and epidemiological aspects. Mem. Inst. Oswaldo Cruz, 99: 673-681.
PubMed - Lehr, T., H. Geyer, K. Maass, M.J. Doenhoff and R. Geyer, 2007. Structural characterization of N-glycans from the freshwater snail Biomphalaria glabrata cross-reacting with Schistosoma mansoni glycoconjugates. Glycobiology, 17: 82-103.
Direct Link - Lima, N.M., A.F. dos Santos, Z. Porfirio , M.O. Goulart and A.E. Santana, 2002. Toxicity of lapachol and isolapachol and their potassium salts against Biomphalaria glabrata, Schistosoma mansoni cercariae, Artemia salina and Tilapia nilotica. Acta Tropica, 83: 43-47.
PubMed - Lockyer, A.E., L.R. Noble, D. Rollinson and C.S. Jones, 2004. Schistosoma mansoni: Resistant specific infection-induced gene expression in Biomphalaria glabrata identified by fluorescent-based differential display. Exp. Parasitol., 107: 97-104.
CrossRef - Lockyer, A.E., L.R. Noble, D. Rollinson and C.S. Jones, 2005. Isolation and characterization of the full-length cDNA encoding a member of a novel cytochrome p450 family (CYP320A1) from the tropical freshwater snail, Biomphalaria glabrata, intermediate host for Schistosoma mansoni. Mem. Inst. Oswaldo Cruz, 100: 259-262.
CrossRef - Lockyer, A.E., J. Spinks, L.R. Noble, D. Rollinson and C.S. Jones, 2007. Identification of genes involved in interactions between Biomphalaria glabrata and Schistosoma mansoni by suppression subtractive hybridization. Mol. Biochem. Parasitol., 151: 18-27.
CrossRef - Lotfy, W.M., R.J. DeJong, B.S. Black and E.S. Loker, 2005. Specific identification of Egyptian Biomphalaria species and possible hybrids using the polymerase chain reaction based on nuclear and mitochondrial loci. Mol. Cell Prob., 19: 21-25.
CrossRefPubMedDirect Link - Madsen, H., 1990. The effect of sodium chloride concentration on growth and egg laying of Helisoma duryi, Biomphalaria alexandrina and Bulinus truncates. J. Moll. Stud., 56: 181-187.
Direct Link - Madsen, H., 1992. Food selection by freshwater snails in the Gezira irrigatiion canals, Sudan. Hydrobiology, 228: 203-217.
CrossRef - Mantawy, M.M., M.A. Hamed, E.M. Sammour and M. Sanad, 2004. Influence of Capparis spinosa and Acacia arabica on certain biochemical haemolymph parameters of Biomphalaria alexandrina. J. Egypt Soc. Parasitol., 34: 659-677.
PubMed - Mantawy, M.M. and A.H. Mahmoud, 2002. Effect of Allium cepa and Allium sativum feeding on glucose, glycogen, protein bands profile and phenol oxidase activity in Biomphalaria alexandrina. J. Egypt Soc. Parasitol., 32: 271-283.
PubMed - Marti, H.P., 1986. Field observations on the population dynamics of Bulinus globosus, the intermediate host of Schistosoma baematobium in the Ifakara area. Tanzania. J. Parasitol., 72: 119-124.
Direct Link - Massoud, A., D.M. Metwally, K.E. Khalifa and F.S. Habib, 2004. Compatibility of Biomphalaria alexandrina snails to infection with Schistosoma mansoni after exposure to sublethal concentrations of Myrrh. J. Egypt. Soc. Parasitol., 34: 995-1008.
PubMed - Mello-Silva, C.C., M.C. de Vasconcellos, J. Pinheiro and M.L.A. Rodrigues, 2006. Physiological changes in Biomphalaria glabrata say, 1818 (Pulmonata: Planorbidae) caused by sub-lethal concentrations of the latex of Euphorbia splendens var. hislopii N.E.B (Euphorbiaceae). MemOrias Instituto Oswaldo Cruz, 101: 3-8.
CrossRefDirect Link - Mostafa, B.B., 2006. Effect of three dormant oils on schistosomiasis and fascioliasis vector snails and its relation with some non-target snails. J. Egypt. Soc. Parasitol., 36: 809-826.
PubMed - Ndifon, G.T. and F.M.A. Ukoli, 1989. Ecology of freshwater snails in South-Western Nigeria. I: Distribution and habitat preferences. Hydrobiologia, 171: 231-253.
CrossRefDirect Link - Ndlela, B. and M.J. Chimbari, 2000. A preliminary assessment of the potential of the Muschovy duck (Carirna maschata) as a biocontrol agent of schistosomiasis intermediate host snails. Cent. Afr. J. Med., 46: 271-275.
PubMed - Njenga, M.N., I.O. Farah, G.K. Muchemi and M. Nyindo, 1998. Peri-portal fibrosis of the liver due to natural or experimental infection with Schistosoma mansoni occurs in the Kenyan baboon. Ann. Trop. Med. Parasitol., 92: 187-193.
CrossRef - Rosa, F.M., A.L. Godard, V. Azevedo, P.M. Coelho and L. Barbosa, 2005. Biomphalaria tenagophila: Dominant character of the resistance to Schistosoma mansoni in descendants of crossbreedings between resistant (Taim, RS) and susceptible (Joinville, SC) strains. Mem. Inst. Oswaldo Cruz, 100: 19-23.
CrossRef - Saad, A. and N. Sayed, 2000. Discriminant analysis of free fatty acids of some Egyptian snails as a step for schistosomiasis control: New trends. J. Egypt Public Health Assoc., 75: 93-106.
PubMed - Smit, A.B., M. De Jong-Brink, K.W. Li, M.M. Sassen and S. Spijker et al., 2004. Granularin, a novel molluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation. FASEB J., 18: 845-847.
PubMed - Takougang, I., J. Meli and F. Angwafo, 2006. Field trials of low dose Bayluscide on snail hosts of schistosome and selected non-target organisms in sahelian Cameroon. Mem. Inst. Oswaldo Cruz., 101: 355-358.
PubMed - Tantawy, A.A., 2006. Molluscicidal effect of fenitrothion and anilofos on Lymnaea natalensis and Biomphalaria alexandrina snails and on the free larval stages of Schistosoma mansoni. J. Egypt Soc. Parasitol., 36: 629-642.
PubMed - Theron, A. and C. Coustau, 2005. Are Biomphalaria snails resistant to Schistosoma Mansoni?. J. Helminthol., 79: 187-191.
PubMed - Thomas, J.D., 1987. An evaluation of the interaction between freshwater pulmonate snail hosts of human schistosomes and macrophytes. Phil. Trans. R. Soc., 315: 75-125.
PubMed - Thomas, J.D. and A.I. Tait, 1984. Control of the snail hosts of schistosomiasis by environmental manipulation a field and laboratory appraisal in the Ibadan area, Nigeria. Phil. Trans. R. Soc. Trop. Med. Hyg., 305: 201-253.
CrossRefDirect Link - Truiti, M.C., I.C. Ferreira, M.L. Zamuner, C.V. Nakamura, M.H. Sarragiotto and M.C. Souza, 2005. Antiprotozoal and molluscicidal activities of five Brazilian plants. Braz. J. Med. Biol. Res., 38: 1873-1878.
PubMed - Utzinger, J., C. Mayombana, K. Mez and M. Tanner, 1997. Evaluation of chemical and physical-morphological factors as potential determinants of Biomphalaria pfeifferi. Mem. Inst. Oswaldo Cruz., 92: 323-328.
PubMed - Utzinger, J., S.H. Xiao, J. Keiser, Z.J. Chen and M. Tanner, 2001. Current progress in development and use of artemether for chemoprophylaxis of major human Schistosoma parasites. Curr. Med. Chem., 8: 1841-1860.
PubMed - Velasquez, L.E., R.L. Caldeira, V. Estrada and O.S. Carvalho, 2002. Morphological and polymerase chain reaction-restriction fragment lenght polymorphism characterization of Biomphalaria kuhniana and Biomphalaria amazonica from Colombia. Mem. Inst. Oswaldo Cruz., 97: 997-1004.
CrossRef - Bashtar, A., S.A. Ahmed, A.M. Soliman and M.A. Hamed, 2006. Biochemical studies on hepatocytes after immunization of mice with schistosomal worm and egg antigens. Asian J. Biochem., 1: 224-235.
CrossRefDirect Link - Ali, S.A. and M.A. Hamed, 2006. Effect of Ailanthus altissima and Zizyphus spina-christi on bilharzial infestation in mice: Histological and histopathological studies. J. Applied Sci., 6: 1437-1446.
CrossRefDirect Link - Flisser, A. and D.J. McLaren, 1989. Effect of praziquantel treatment on lung-stage larvae of Schistosoma mansoni in vivo. Parasitology, 98: 203-211.
CrossRefDirect Link - Hamed, M.A., 2006. Excretory secretory product of Fasciola hepatica worm protects against Schistosoma mansoni infection in mice. Indian J. Exp. Biol., 44: 554-561.
PubMedDirect Link