
Kossi M. Joppa
Laboratory of Physiology and Pharmacology, Faculty of Medicine and Pharmacy, University of Lome, P.O. Box 1515, Togo
LiveDNA: 228.31884
Kossi Metowogo
Laboratory of Physiology and Pharmacology, Faculty of Medicine and Pharmacy, University of Lome, P.O. Box 1515, Togo
Irenee M. Kueviakoe
Laboratory Service of CHU Campus, Faculty of Medicine and Pharmacy, University of Lome, P.O. Box 1515, Togo
Povi Lawson-Evi
Laboratory of Physiology and Pharmacology, Faculty of Medicine and Pharmacy, University of Lome, P.O. Box 1515, Togo
Ahoefa Vovor
Laboratory of Hematology of CHU Sylvanus Olympio, Faculty of Medicine and Pharmacy, University of Lome, P.O. Box 1515, Togo
Kodjo Aklikokou
Laboratory of Physiology and Pharmacology, Faculty of Medicine and Pharmacy, University of Lome, P.O. Box 1515, Togo
Messanvi Gbeassor
Laboratory of Physiology and Pharmacology, Faculty of Medicine and Pharmacy, University of Lome, P.O. Box 1515, Togo
American Journal of Drug Discovery and Development
Year: 2014 | Volume: 4 | Issue: 1 | Page No.: 58-65







ABSTRACT
Sickle cell anemia is a genetic disease in which the final step leading to morbidity is the polymerization of deoxyhemoglobin S. Therefore, one of the recommended treatments for sickle cell disease is its inhibition. For this reason, the aim of this study was to evaluate the inhibitory properties of Morinda lucida semi-ethanolic extract on the polymerization of deoxyhemoglobin S. The deoxyhemoglobin polymerization was induced by sodium dithionite in a phosphate buffer in the absence and presence of M. lucida extract. Then the absorption spectrum of each sample was determined and the absorbance at 578 and 630 nm, respectively for hemoglobin and methemoglobin was measured. The solution of deoxyhemoglobin with dithionite presents a very cloudy appearance unlike negative control (without dithionite) which has a clear appearance. The results showed a 58.6% increase in turbidity (absorbance) compared to the negative control. M. lucida extract (1.5 mg cm-3) reduces very significantly the polymerization (p<0.001). The absorbance is 0.636 versus 1.331 for the positive control and 0.839 for the negative control. The oxidation of hemoglobin to methemoglobin as revealed by the absorption peak at 630 nm is significantly inhibited by the extract (p<0.000). M. lucida induces 52.5% inhibition of oxidation. This antioxidant property was confirmed by the results of the 2, 2-diphenyl-1-picrylhydrazyl (DPPH) test. Results showed that the semi-ethanolic extract of M. lucida very significantly decreases the deoxyhemoglobin S polymerization. Moreover the extract has an antioxidant property which prevents hemoglobin oxidation in methemoglobin.
PDF Abstract XML References Citation
Received: September 06, 2013;
Accepted: October 31, 2013;
Published: March 10, 2014
How to cite this article
Kossi M. Joppa, Kossi Metowogo, Irenee M. Kueviakoe, Povi Lawson-Evi, Ahoefa Vovor, Kodjo Aklikokou and Messanvi Gbeassor, 2014. Study of the Inhibition of Deoxyhemoglobin S Polymerization by the Hydroalcoholic
Extract of Morinda lucida. American Journal of Drug Discovery and Development, 4: 58-65.
DOI: 10.3923/ajdd.2014.58.65
URL: https://scialert.net/abstract/?doi=ajdd.2014.58.65
DOI: 10.3923/ajdd.2014.58.65
URL: https://scialert.net/abstract/?doi=ajdd.2014.58.65
INTRODUCTION
Sickle cell anemia is a genetic disorder that affects the physiology of red blood cells. It is caused by a mutation leading to the synthesis of abnormal hemoglobin (HbS). This hemoglobin acquires properties which alter the physiology, the structure and the rheology of red blood cells (Bunn, 1997; Hebbel, 1991). Of these characteristic properties of HbS involved in the pathophysiology of sickle cell anemia, the polymerization is the property intimately involved in the vicious circle of sickle cell disease. Unlike normal hemoglobin (HbA), when the oxygen saturation decreases, the deoxyhemoglobin S molecules polymerize to form fibers of varying length (Catonne, 2002; Ohnishi, 2001; Bunn, 1997). This polymerization reduces the affinity of HbS for oxygen and modifies or enhances the permeability of ion and water flux in the erythrocyte membrane (Bunn, 1997). The direct consequence is the reduction of oxygen to the body and the dehydration of RBCs leads to the sickling of red blood cells (Bunn, 1997; Hebbel, 1991). These sickled red cells induce vasocclusion which is implicated in the organic complications of sickle cell disease (Asakura et al., 1974) because of the rigidity of red blood cells who lose their flexibility (Noguchi and Schechter, 1985).
Several studies have shown that the inhibition of deoxyhemoglobin polymerization would be a basic treatment for sickle cell disease (Noguchi and Schechter, 1985; Moffat and Gibson, 1974; Chang et al., 1983; Dean and Schechter, 1978; Eaton et al., 1976; Sunshine et al., 1978) since it is at the heart of the pathophysiology of sickle cell disease.
In latest study, the anti-sickling effects of M. lucida extract have been highlighted (Joppa et al., 2008). The aim of this study is to investigate the polymerization of deoxyhemoglobin S molecules inhibitory property of M. lucida semi-ethanolic extract.
MATERIALS AND METHODS
Materials
Biological material: The leaves of Morinda lucida were collected in the rainy season at Adéticopé, town enjoying a tropical climate and situated at 20 km to the north of Lomé. The plant extract is prepared from the maceration of dried leaves under cooling in an ethanol-water mixture at 80% (80:20). The filtrate of this maceration was evaporated under vacuum in a bath of 45°C using a Buchi Rotavapor R-210.
Sickle cell blood samples were obtained from consenting volunteer donors and collected into the vacutainer containing anticoagulant.
Chemicals: For induction of the polymerization, the sodium chloride (NaCl) was used to wash the Red Blood Cells (RBC), potassium dihydrogen phosphate (K2HPO4) and potassium hydrogen phosphate (KH2PO4) for the preparation of a phosphate buffer and sodium dithionite (Na2S2O4) for the deoxygenation of the hemoglobin solution. 2, 2-diphenyl-1-picrylhydrazyl (DPPH) and quercetin are used for antioxidant test.
METHODS
Preparation of hemoglobin solution: Blood samples were drawn by venipuncture using a vacutainer containing EDTA and centrifuged at 3000 rpm for 15 min. The RBCs were obtained and washed three times with NaCl 0.9% at 5000 rpm for 5 min to remove albumin which exacerbates the polymerization (Noguchi and Schechter, 1985; Singer and Singer, 1953). The washed RBCs are hemolysed by mixing one volume of RBCs in six volumes of distilled water (2 cm3 of pellet in 12 cm3 of water) (Baledent, 2000). The mixing is placed at 4°C for 15 min. The hemolysate obtained was then centrifuged at 13,500 rpm for 10 min to remove the stroma (Adachi and Asakura, 1979).
Induction and inhibition of polymerization: The induction of polymerization is based on the principle of the insolubility of hemoglobin S in the absence of oxygen. A reducing solution was prepared by dissolving 25 mg of dithionite in 2 cm3 of phosphate buffer. The phosphate buffer (pH 7.4) is obtained by dissolving 28.2 g of K2HPO4 and 16.0 g KH2PO4 in 100 cm3 of distilled water (Baledent, 2000; Liao et al., 1996).
Deoxyhemoglobin polymerization is then induced by mixing 4 volumes of the reducing solution with 1 volume of the hemoglobin solution. In the negative control we replaced the dithionite solution with the phosphate buffer used to prepare the solution of dithionite.
To study the inhibitory effect of M. lucida we added 1 cm3 of the extract in the test tubes against 1 cm3 of distilled water to the control tubes.
Then using the Vernier spectrophotometer (Ocean Optics Inc.), the polymerization rate is determined by turbidimetry (Liao et al., 1996; Noguchi and Schechter, 1985). The absorption spectrum of each sample is collected and the absorbance at 578 nm for hemoglobin and 630 nm for methemoglobin are measured (Asakura et al., 1974).
The same solutions of hemoglobin concentration for different batches was used because the concentration of deoxyhemoglobin is, in itself, a factor involving polymerization (Noguchi and Schechter, 1985).
Statistical analysis: The data are treated by Systat 11 and expressed as Mean±SEM (Standard Error of the Mean). The differences are carried out by the Student test and it been consider significant for p<0.05. The graphs are performed with GraphPad Prism® 5.00.
RESULTS AND DISCUSSION
Induction of polymerization: As opposed to the control tubes which kept a clear appearance, the incubation of the hemoglobin solution at room temperature with sodium dithionite induced a very cloudy appearance of the solution after 15 min. The measurement of the polymerization by turbidimetry (Noguchi and Schechter, 1985) revealed a 58.6% increasing of turbidity in the tube with dithionite compared to the control (p<0.001). Indeed, the absorbance at 578 nm of the control is 0.839 against 1.331 for the test tubes (Fig. 1). The turbidity in the presence of dithionite is due to the reducing property of dithionite which deprives hemoglobin molecules of oxygen. These deoxyhemoglobin S molecules polymerize in the absence of oxygen to form fibers of deoxyhemoglobin (Bunn, 1997; Eaton et al., 1976; Embury, 1986; Wang et al., 2000). According to studies by Noguchi et al. (1980) there is a strong negative correlation between oxygen saturation and the formation of these polymers of deoxyhemoglobin. However, dithionite is a reducing agent that induces 0% oxygen saturation. The total oxygen desaturation associated with the phosphate buffer at the high concentration used (Liao et al., 1996; Adachi and Asakura, 1979) led to a high rate of formation of deoxyhemoglobin S polymers giving a very cloudy aspect of hemoglobin S solutions (Baledent, 2000; Wang et al., 2000).
Moreover the absorption spectrum of hemoglobin S obtained before and after polymerization has the same characteristics as those established by Asakura et al. (1974). Indeed, three regular peaks at 421, 540 and 576 nm and an irregular peak at 630 nm were obtained (Fig. 2). These peaks show the conformity of our results with those obtained by Asakura et al. (1974) and validates our methods used to evaluate the inhibitory properties of deoxyhemoglobin polymerization.
In this study, turbidity was used to quantify the deoxyhemoglobin polymerization instead of the sedimentation method (Asakura et al., 1974) usually used to determine the rate of deoxyhemoglobin polymerization because the sedimentation method underestimates the polymerization.
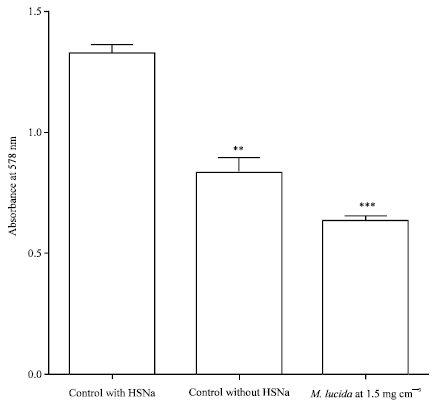 |
| Fig. 1: | Effect of M. lucida on polymerization of hemoglobin S in solution induced by dithionite. In presence of dithionite, the deoxyhemoglobin S molecule form a polymer leading to deoxyhemoglobin S fibers conferring a cloudy aspect to the solution. The polymerization rate was determinend by turbidimetry. This figure showed that the deoxyhemoglobin polymers were formed in presence of dithionite (**p<0.0001 for positive control vs. negative control) but M. lucida inhibit this polymerization and improv the deoxyhemoglobin S solubility comparatively to the negative control (***p<0.01 for M. lucida vs. negative control) |
Indeed, after polymerization we have polymers of different sizes in solution. Then, the fibers of hemoglobin of small size like hemoglobin tetramers remain in suspension at the interface of the pellet and the supernatant. Also, the pellet contains some trapped free hemoglobin molecules (Noguchi and Schechter, 1985).
Inhibition of polymerization induced by extract of M. lucida: Figure 1 and 3 show a very significant decrease in polymerization in the presence of M. lucida extract (p<0.000). The plant extract at 1.5 mg cm-3 induces a 52% decrease in turbidity. The absorbance in the presence of the plant extract is also lower than the negative control (without sodium dithionite). It is 0.636 in the presence of the extract against 0.839 for the negative control and 1.331 for the control with dithionite.
In these experimental conditions the degree of turbidity expresses the rate of polymerization of deoxyhemoglobin molecules (Baledent, 2000; Liao et al., 1996; Hofrichter et al., 1976; Moffat and Gibson, 1974) due to the reduction of their solubility in reduced solution (Danish et al., 1994). In the light of these results it state that the alcoholic extract of M. lucida inhibited the polymerization of deoxyhemoglobin molecules and therefore increases their solubility.
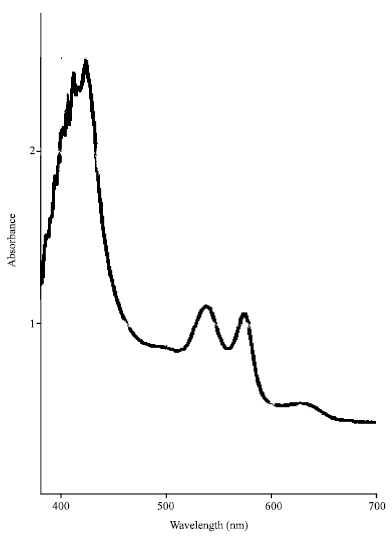 |
| Fig. 2: | Absorption spectrum of hemoglobin S in solution. The hemoglobin spectrum was obtained with Vernier spectrometer and it showed the four characteristic absorption pics. The first pic at 421 nm is not clear because a dilution to obtain low concentration of hemoglobin S solution may run over the pics of 540, 578 and 630 nm |
This property explains the antisickling effect of this plant as highlighted in our previous studies (Joppa et al., 2008) seeing that the direct mechanism inducing sickling is the deoxyhemoglobin S polymerization (Bunn, 1997). Indeed, in absence of oxygen or in case of the dehydration of sickle red blood cells, deoxyhemoglobin molecules become insoluble and polymerize to form fibers with a length exceeding the diameter of red blood cells, causing or accentuating dehydration. Red blood cells deform (sickling) and become more rigid (Bunn, 1997). These changes in red blood cells underlie the vicious cycle leading to the sickle cell crises.
So, to conclude, M. lucida used in the treatment of sickle cell crises intervenes upstream of the vicious cycle of the pathophysiology of sickle cell crises by enhancing the solubility of deoxyhemoglobin.
Inhibition of oxidation of hemoglobin to methemoglobin: Figure 3 shows an absorption peak at 630 nm corresponding to the absorption of methemoglobin (Asakura et al., 1974).
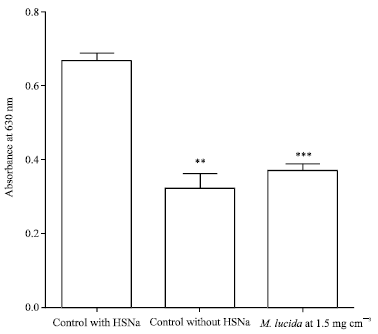 |
| Fig. 3: | Inhibition of hemoglobin oxidation in methemoglobin in presence of M. lucida. The sodium dithionite (HSNa) have induced very significant (**p<0.0001) conversion of hemoglobin in methemoglobin comparatively to the negative control. This formation of methemoglobin is signicatively inhibited by M. lucida at 1.5 mg cm-3 (p<0.0001), ***p>0.05 for M. lucida at 1.5 mg cm-3 vs. negative control (control without HSNa) |
This absorption is 0.668 for the control against 0.317 for the test showing a decreasing of methemoglobin formation in presence of M. lucida. Our results carry out M. lucida protect effect against oxidation of hemoglobin in methemoglobin induced by dithionite (p<0.000). Indeed, dithionite releases NO which reacts with iron of hemoglobin and convert it in methemoglobin (Bonaventura et al., 2002). This methemoglobin in which iron is in trivalent form is unfit for the binding of oxygen in vivo.
This reduction of methemoglobin formed in the presence of the extract reveals an antioxidant property o f the extract of M. lucida. In order to test this hypothesis, we’ve studied the antioxidant properties of the plant by the DPPH test according to the method as described by Lawson-Evi et al. (2011). This DPPH test showed that the semi-ethanolic extract inhibit the oxidant power of quercetin with a correlation coefficient of -0.98 (Fig. 4a). Even 0.05 mg cm-3 of M. lucida extract inhibition rate is 6.74% while 0.5 and 1 mg cm-3 inhibition rate is, respectively 48.76 and 85.28% (Table 1). Then the IC50 (50% inhibition of DPPH oxidation) obtained for the extract is 0.52 mg cm-3. These antioxidant properties of the ethanol extract of M. lucida has been also highlighted by the work of Ogunlana et al. (2008). Therefore, this antioxidant property of the plant justifies the inhibition of oxidation of hemoglobin to methemoglobin in the presence of the plant extract. Moreover it should be noted that at the high concentrations (≥2.5 mg cm-3), the extract appears pro-oxidant (Fig. 4b). This pro-oxidant effect of the plant revealed by the DPPH test have not induced a significant increasing of methemoglobin formation at the concentrations of 1.5 and 3 mg cm-3 (p = 0.196).
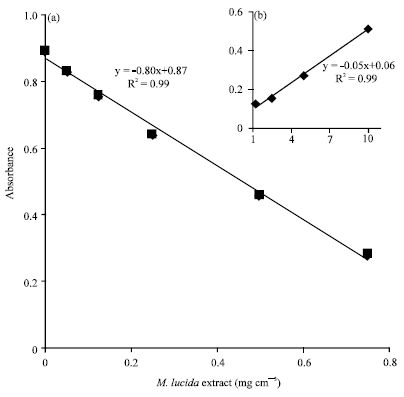 |
| Fig. 4(a-b): | Antioxidant activity of M. lucida on DPPH radicals, (a) Extract reduces significantly oxidant radicals with lows concentrations and (b) At the highest concentrations of the extract (≥2.5 mg cm-3) antioxidant activity is reduced |
| Table 1: | Inhibition rate of the DPPH oxidant power by M. lucida |
 | |
Inhibition rate: 100x(Absorbance control-Absorbance test)/Absorbance control | |
CONCLUSION
From the Itano test modified, we have induced and quantified deoxyhemoglobin S polymerization by turbidimetry. This method is a direct measurement of the deoxyhemoglobin polymers formed in suspension. This polymerization induced by sodium dithionite was very significantly reduced by the M. lucida extract. This extract also prevented the oxidation of hemoglobin to methemoglobin by its antioxidant properties. In sum, M. lucida increases the solubility of deoxyhemoglobin S and protects the latter against the free radicals oxidant power.
REFERENCES
- Adachi, K. and T. Asakura, 1979. The solubility of sickle and non-sickle hemoglobins in concentrated phosphate buffer. J. Biol. Chem., 254: 4079-4084.
PubMedDirect Link - Asakura, T., T. Ohnishi, S. Friedman and E. Schwartz, 1974. Abnormal precipitation of oxyhemoglobin S by mechanical shaking. Proc. Nat. Acad. Sci. USA, 71: 1594-1598.
Direct Link - Bonaventura, C., C.H. Taboy, P.S. Low, R.D. Stevens, C. Lafon and A.L. Crumbliss, 2002. Heme redox properties of S-nitrosated hemoglobin A0 and hemoglobin S Implications for interactions of nitric oxide with normal and sickle red blood cells. J. Biol. Chem., 277: 14557-14563.
CrossRefDirect Link - Bunn, H.F., 1997. Pathogenesis and treatment of sickle cell disease. N. Engl. J. Med., 337: 762-769.
CrossRefPubMedDirect Link - Catonne, Y., 2002. Aspects orthopediques de la drepanocytose. [Orthopedic aspects of sickle cell disease]. Proceedings of the International Conferences d'Enseignement de la Sofcot, Volume 79, December 12, 2002, France, pp: 245-262.
Direct Link - Chang, H., S.M. Ewert, R.M. Bookchin and R.L. Nagel, 1983. Comparative evaluation of fifteen anti-sickling agents. Blood, 61: 693-704.
PubMed - Dean, J. and A.N. Schechter, 1978. Sickle-cell anemia: Molecular and cellular bases of therapeutic approaches. N. Engl. J. Med., 299: 752-763.
CrossRefDirect Link - Eaton, W.A., J. Hofrichter and P.D. Ross, 1976. Delay time of gelation: A possible determinant of clinical severity in sickle cell disease. Blood, 47: 621-627.
Direct Link - Embury, S.H., 1986. The clinical pathophysiology of sickle cell disease. Ann. Rev. Med., 37: 361-376.
CrossRefDirect Link - Hebbel, R.P., 1991. Beyond hemoglobin polymerization: The red blood cell membrane and sickle disease pathophysiology. Blood, 77: 214-237.
Direct Link - Hofrichter, J., P.D. Ross and W.A. Eaton, 1976. Supersaturation in sickle cell hemoglobin solutions. Proc. Natl. Acad. Sci. USA, 73: 3035-3039.
Direct Link - Joppa, K.M., A. Vovor, K. Eklu-Gadegbeku, A. Agbonon, K. Aklikokou and M. Gbeassor, 2008. Effect of Morinda lucida Benth. (Rubiaceae) and Newbouldia leavis P. Beauv. (Bignoniaceae) on sickling of red blood cells. Med. Trop., 68: 251-256.
PubMed - Lawson-Evi, P., K. Eklu-Gadegbeku, A. Agbonon, K. Aklikokou, E. Creppy and M. Gbeassor, 2011. Antidiabetic activity of Phyllanthus amarus schum and thonn (Euphorbiaceae) on alloxan induced diabetes in male wistar rats. J. Applied Sci., 11: 2968-2973.
CrossRef - Liao, D., J.J. Martin de llano, J.P. Himanen, M.J. Manning and F.A. Ferrone, 1996. Solubility of sickle hemoglobin measured by a kinetic micromethod. Biophys. J., 70: 2442-2447.
CrossRefDirect Link - Moffat, K. and Q.H. Gibson, 1974. The rates of polymerization and depolymerization of sickle cell hemoglobin. Biochem. Biophys. Res. Commun., 61: 237-342.
CrossRefDirect Link - Noguchi, C.T., D.A. Torchia and A.N. Schechter, 1980. Determination of deoxyhemoglobin S polymer in sickle erythrocytes upon deoxygenation. Proc. Natl. Acad. Sci. USA, 77: 5487-5491.
Direct Link - Noguchi, C.T. and A.N. Schechter, 1985. Sickle hemoglobin polymerization in solution and in cells. Annu. Rev. Biophys., 14: 239-263.
CrossRefDirect Link - Ogunlana, O.E., O.O. Ogunlana and E.O. Farombi, 2008. Morinda lucida: Antioxidant and reducing activities of crude methanolic stem bark extract. Adv. Nat. Applied Sci., 2: 49-54.
Direct Link - Ohnishi, S.T. and T. Ohnishi, 2001. In vitro effects of aged garlic extract and other nutritional supplements on sickle erythrocytes. J. Nutr., 131: 1085S-1092S.
CrossRefDirect Link - Singer, K. and L. Singer, 1953. Studies on abnormal hemoglobins: VIII. The gelling phenomenon of sickle cell hemoglobin, its biologic and diagnostic significance. Blood, 8: 1008-1023.
Direct Link - Sunshine, H.R., J. Hofrichter and W.A. Eaton, 1978. Requirements for therapeutic inhibition of sickle haemoglobin gelation. Nature, 275: 238-240.
CrossRefDirect Link - Wang, Z., G. Kishchenko, Y. Chen and R. Josephs, 2000. Polymerization of deoxy-sickle cell hemoglobin in high-phosphate buffer. J. Struct. Biol., 131: 197-209.
CrossRefDirect Link - Danish, E.H., D.W. Lundgren and J.W. Harris, 1994. Inhibition of hemoglobin S polymerization by N-terminal band 3 peptides: New class of inhibitors: Solubility studies. Am. J. Hematol., 47: 106-112.
CrossRefPubMedDirect Link