
E. Takwa Ellakwa
Department of Physical Chemistry, Egyptian-Russian University, Egypt
American Journal of Drug Discovery and Development
Year: 2014 | Volume: 4 | Issue: 4 | Page No.: 248-252







ABSTRACT
The effect of nutrition on different host plants (cabbage, castor bean and cotton) on midgut redox potentional (Eh), midgut pH and digestive enzymes of the fourth larval instar of Agrotis ipsilon was studied. The results showed that all larvae had strongly alkaline midguts pH ranging from 9-9.75. There were no significant differences between midgut pH of larvae fed for 24 h or that fed for 7 days, effect different host plants, reflecting the rapidity by which the larvae adjust their midgut pH. On the contrary, plant species had a significant effect on mid gut Eh. Conditions in all guts were oxidizing ranging from +171 to +250 mV i.e., all tested midguts Eh were positive. The results also revealed that continuous feeding on host, plants for 7 days significantly lowered the oxidizing state of gut, except cabbage, as compared to those larvae fed for 24 h. Each host plant had own specific effect on protease, lipase and invertase. However lipase was the least enzyme that affected. The study clarifies the effect of host plants on midgut physicochemistry which mid gut affect digestion and gut physiology, ultimately affecting the normal growth of the pest.
PDF Abstract XML References Citation
Received: July 10, 2014;
Accepted: October 15, 2014;
Published: December 27, 2014
How to cite this article
E. Takwa Ellakwa, 2014. Effect of Host Plant on Gut Physicochemistry and Digestive Enzymes of the Cutworm (Agrotis ipsilon) Larvae. American Journal of Drug Discovery and Development, 4: 248-252.
DOI: 10.3923/ajdd.2014.248.252
URL: https://scialert.net/abstract/?doi=ajdd.2014.248.252
DOI: 10.3923/ajdd.2014.248.252
URL: https://scialert.net/abstract/?doi=ajdd.2014.248.252
INTRODUCTION
The chemical and enzymatic reactions associated with digestion, absorption and detoxification of food in the insect gut are sensitive to changes in pH, redox potential (Eh) and other physicochemical parameters. Foliage contains nutrients refractory to digestion and allelochemicals of potential toxicity so, that herbivores are frequently nutrient limited (Bernays and Barbehenn, 1987; Slansky, 1992).
Physicochemical conditions of the midgut in caterpillars are likely to have a major impact on nutrient digestion and allelochemical activity. The efficiency of extraction of nutrients from plant tissues has been shown to depend on pH (Felton and Duffey, 1991) and also depend on Eh (Appel and Martin, 1990).
The previous documentations illustrated that diet pH, buffer concentration and buffering capacity affect growth utilization by the larvae of Manduca sexta (Karowe and Martin, 1993) and Spodoptra littoralis (Amin et al., 2006). Impact of two host plants species on insect gut pH (Amin, 2007) was studied. However, information about the influence of host plant on Agrotis ipsilon larvae are lacking.
The purpose of the present study was: (1) To determine midgut pH ranges (2) To show that the pH of the midgut is independent of host plant or not to evaluate the effect of nutrition on different host plants (cotton, castor bean and cabbage) on midgut redox potential and pH of the fourth larval instar and (3) To detect the effect of the host plant on some digestive enzymes, since, gut physicochemistry may also strongly impact digestion and nutrient assimilation in herbivorous insects.
MATERIALS AND METHODS
Insects and host plants: The used strain of A. ipsilon was a laboratory strain, obtained as newly hatched larvae from a culture of Central Agricultural Pesticides Laboratory, Dokki, Egypt. Larvae were reared under constant conditions of 25°C and 70±5% R.H. They were divided into 6 groups (n = 100), 3 of which fed on host plants from hatching till the fourth larval instar, the others fed on host plants for only 24 h but from hatching till the experiment time they fed on laboratory artificial diet (pH = 6).
Insect gut pH and redox potential measurements: To determine the effect of host plants on insect gut conditions, the gut conditions of the 4th larval instar were measured. Older instar caterpillars were chosen to work with because their larger size facilitates gut measurements. Larvae were immobilized on ince and the gut exposed by a ventral longitudinal incision. The body was pinned away from the gut blotted dry to avoid contamination by blood. The main steps described by Johnson and Felton (1996) were followed. Midgut lumen pH and Eh were recorded by using a micro-needle pH electrode (Sam Agulion, CT), a 0.02 in platinum electrode (Microelectrodes, inc.) and liquid junction Ag-AgCl reference electrode (Mere-I, WpI) placed in contact with moistened filter paper underneath the insect. The microelectrodes inserted through the midgut epithelium into the lumen. Redox measures typically showed a precipitous drop 10-20 sec after insertion followed by a gradual leveling off, so, redox values were recorded at 5 sec intervals for 60 sec. Then, midgut pH was recorded at the same location. Redox potentials were standardizing relative to a hydrogen reference probe by adding 200 mv (the potential of the reference electrode) to each value recorded.
Digestive enzyme assays: The fourth larval instar guts were dissected by immersing in iso osmotic saline (0.15 KCl, pH 7). With the aid of a sharp razor, the dorsal side of the body was longitudinally opened, exposing the alimentary canal, cutting it from anterior and posterior ends. Then the guts were rinsed twice with saline. The guts were homogenized (5 guts/1 mL distilled water) and the supernatant was analyzed for tryptic (protease), invertase and lipase activity.
The proteolytic activity was determined according to Birk et al. (1962) by the casein digestion method. Invertase was determined by the method of Ishaaya and Swirski (1976) which is based on the digestion of sucrose by invertase. The free oldhydic group of glucose formed after sucrose digestion was determined using 3,5-dinitrosalicylic acid reagent. Lipase activity was determined by a slight modification of the procedure of Tahoun and Abdel-Ghaffar (1986). The method was based on the determination of the decrease in ester content of triolein as substrate.
Statistical analysis: Results were analyzed by one way analysis of variance (ANOVA) using Costat statistical software. ANOVA F-statistics was significant at p<0.01. Means by the Duncan’s multiple range test were compared.
RESULTS AND DISCUSSION
Effect of host plant on midgut contents pH: Analysis of variance of the midgut contents pH of the 4th larval instar reared on 3 different host plants revealed that there was a significant decrease (Table 1) ranging from 9-9.75.
Larvae fed on castor bean leaves having a more alkaline midgut than other larvae (pH 9.75) while larvae feed on cotton leaves having the lowest (8.9). The results also showed that there is no significant difference between larvae fed for 24 h or that fed for 7 days on host plant with respect to their midgut pH.
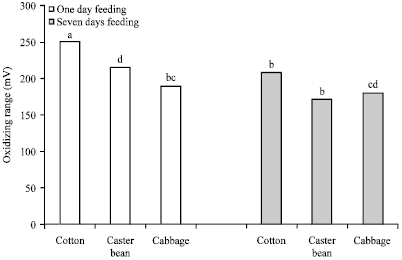 |
| Fig. 1: | Effect of host plant on midgut redox potential of Agrotis ipsilon fourth larval instar. Vertical lines give standard deviations. Bars with different letters are significantly different at p<0.01 |
| Table 1: | Effect of different host plants on midgut contents pH of Agrotis ipsilon fourth larval instar |
 | |
Means with different letters are different at p<0.01 | |
Effect of host plants on midgut redox potential: On the contrary, plant species had a significant effect on the midgut Eh (Fig. 1) conditions in all guts were oxidizing ranging from +171 to +250 mV. i.e., all tested midguts redox potential were positive. The results also revealed that continuous feeding on host plants for 7 days significantly lowered the oxidizing state of gut, except cabbage.
Effect of host plants on digestive enzymes: Feeding of larvae from egg hatching till the fourth larval instar (7 days) on different host plants significantly affect digestive enzymes. Continuous feeding for such period was selected to ensure the effect of plant chemistry on gut cells. Results in Table 2 demonstrates that each plant had own specific effect. However, lipase was the least affected enzyme. It was shown that larvae fed on cotton had the highest level of protease and the lowest level of invertase. On the contrary larvae fed on caster bean leaves had the highest level of invertase and the lowest level of protease. Larvae fed on cabbage had the lowest level of lipase.
Midgut pH was not affected by the host plant used in this study, remaining strong alkaline despite the ingestion of acidic foliage. Preliminary studies confirmed that most leaf tissues have low pH values (Smith and Raven, 1979). Schultz and Lechowicz (1986) have reported that the average pH of freshly homogenized leaf tissues from 23 hard wood tree species was 5 (range 4.1-6.2).
The independence of midgut pH reflects the action of the powerful cation pump in the midgut epithelium of caterpillars (Dow, 1987).
The alkalinity of midgut pH after feeding for 24 h on host plants reflects the rapidity by which larvae adjust their midgut pH. With respect to cotton plant, midgut pH was significantly decreased than feeding on castor bean and cabbage leaves.
| Table 2: | Digestive enzymes activity after feeding of Agrotis ipsilon larvae for 7 days on different host plants |
 | |
Protease activity was expressed as O.D units min-1 gut-1, Lipase activity was expressed as μM oleic acid min-1 gut-1, Invertase activity was expressed as μg glucose min-1 gut-1. Means, within columns, bearing different subscripts are significantly different at p<0.01 | |
Cotton plant might slightly affect the efficiency of the cation pump of the cation pump of midgut epithelium which has been estimated to consume 10% of cellular ATP (Dow, 1987). Adjusting of midgut pH might affect growth of larvae. The growth of Monduca sexta is reduced by 37% on acidic well buffered artificial diets (Karowe and Martin, 1993). Amin et al. (2006) concluded that larvae of S. litoralis larvae probably to overcome unsuitable physicochemical conditions of the introduced diet increased their demand on energy through consuming digested or assimilated food like carbohydrates instead of building new tissues which finally influencing larval performance parameters.
Earlier studies assumed that insects actively maintain specific gut conditions (Appel and Martin, 1990). In contrary, the present results indicate that midgut redox potential is strongly affected by host plant. We found significant variation in midgut redox potential when larvae fed different host plants. Johnson and Felton (1996) found that redox potential was influenced by both plant and lepidopteran species. Midgut is closely matched the redox potential of foliage homogenized under alkaline but not neutral pH, suggesting that alkalinity has a strong influence on the reducing strength of foliage in the gut lumen continous feeding from egg hatching till the fourth larval instar of A. ipsilon, in the present study, lowered Eh of the midgut. The lower Eh of the midgut reflects electron availability which may be produced by the alkaline pH, higher oxygen availability and for plant cell lysis (Appel and Maines, 1995). Plant leaves contain phenolics which are good reducing agents at or above pH 8.5, the pKa of henoxyl hydrogen but poor electron donors at acid pH 5 (Appel, 1993). Takwa (2007) reported that the observed reduction in S. littoralis weight gain could be attributed at least in part, to the effect on digestion after exposure to phenolic acids but not excluding the presence of other additional mechanisms. The results suggest that one of the these mechanisms is the effect on redox potential which might change the optimal conditions of the enzyme activity. Another mechanism is the effect of phenolics on midgut cell as reported by Ahmad (1992) due to direct oxidative challenge to the digestive system of the activity feeding insect. Amin (2007) related the different susceptibilies to insecticides via altering detoxification enzyme activity after rearing of larvae on different host plants to the difference in plant chemistry of host plants fed by larvae.
CONCLUSION
The study clarify the effect of host plants on physicochemistry of midgut which might affect digestion and gut physiology, ultimately affecting growth and vital activities of the pest.
REFERENCES
- Appel, H.M. and M.M. Martin, 1990. Gut redox conditions in herbivorous lepidopteran larvae. J. Chem. Ecol., 16: 3277-3290.
CrossRefDirect Link - Amin, T.R., S. Abo El-Enin and T.H. Ellakwa, 2006. Physicochemistry of diet influences nutritional indices and performance of the cotton leafworm, Spodoptera littoralis (Boisd.). Pak. J. Biol. Sci., 9: 2802-2807.
CrossRefDirect Link - Appel, H.M. and L.W. Maines, 1995. The influence of host plant on gut conditions of gypsy moth (Lymantria dispar) caterpillars. J. Insect Physiol., 41: 241-246.
CrossRefDirect Link - Appel, H.M., 1993. Phenolics in ecological interactions: The importance of oxidation. J. Chem. Ecol., 19: 1521-1552.
CrossRefDirect Link - Ahmad, S., 1992. Biochemical defence of pro-oxidant plant allelochemicals by herbivorous insects. Biochem. Syst. Ecol., 20: 269-296.
CrossRefDirect Link - Birk, Y., I. Harpaz, I. Ishaaya and A. Bondi, 1962. Studies on the proteolytic activity of the beetles Tenebrio and Tribolium. J. Insect Physiol., 8: 417-429.
CrossRefDirect Link - Felton, G.W. and S.S. Duffey, 1991. Reassessment of the role of gut alkalinity and detergency in insect herbivory. J. Chem. Ecol., 17: 1821-1836.
CrossRefDirect Link - Ishaaya, I. and E. Swirski, 1976. Trehalase, invertase and amylase activities in the black scale, Saissetia oleae and their relation to host adaptability. J. Insect Physiol., 22: 1025-1029.
CrossRefDirect Link - Johnson, K.S. and G.W. Felton, 1996. Physiological and dietary influences on midgut redox conditions in generalist lepidopteran larvae. J. Insect Physiol., 42: 191-198.
CrossRefDirect Link - Karowe, D.N. and M.M. Martin, 1993. Determinants of diet quality: The effects of diet pH, buffer concentration and buffering capacity on growth and food utilization by larvae of Manduca sexta (Lepidoptera: Sphingidae). J. Insect Physiol., 39: 47-52.
CrossRefDirect Link - Schultz, J.C. and M.J. Lechowicz, 1986. Hostplant, larval age and feeding behavior influence midgut pH in the gypsy moth (Lymantria dispar). Oecologia, 71: 133-137.
CrossRefDirect Link - Smith, F.A. and J.A. Raven, 1979. Intracellular pH and its regulation. Annu. Rev. Plant Physiol., 30: 289-311.
CrossRefDirect Link