
Amal Fayyad
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600, Bangi Selangor, Malaysia
LiveDNA: 970.8274
Nazlina Ibrahim
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600, Bangi Selangor, Malaysia
Wan Ahmad Yaacob
School of Chemical Sciences and Food Technology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600, Bangi Selangor, Malaysia
American Journal of Drug Discovery and Development
Year: 2013 | Volume: 3 | Issue: 2 | Page No.: 84-94







ABSTRACT
Acute and recurrent Herpes simplex virus type 1 (HSV-1) infections remain an important problem due to the emergence of acyclovir (ACV) resistant virus that makes the search for novel antiviral substances imperative. The aim of this study was to evaluate the antiviral activity of hexane fraction from Marrubium vulgare against Herpes simplex virus type 1 and to determine the mechanism of its action. Cytotoxicity, antiviral assay, time of addition, time of removal, virucidal activity, cells pre-treatment, penetration, staining and thin sections by transmission electron microscope and quantitative PCR assays were conducted to determine the mode action of hexane fraction. Cytotoxicity was demonstrated using the MTT assay, antiviral activities were demonstrated using plaque reduction assay and analysis of data was performed by one way ANOVA and T-test. Hexane fraction was found to inhibit type 1 Herpes simplex virus (HSV-1) in a dose-dependent manner. Further studies revealed that hexane fraction inactivated HSV-1 particles directly as was verified by transmission electron microscopy. Pretreatment of cells and penetration assays did not affect the inhibition of the virus. Hexane fraction suppressed the intracellular growth of HSV-1 in post-infection in time of addition (from 2-8 h p.i.), in time of removal (from 12-48 h p.i) and inhibited HSV-1 IE, E and L gene expressions. In summary, our findings suggest that the hexane fraction can inactivate extracellular viral particles and inhibit viral biosynthesis in host cells. The results also showed that the hexane fraction from Marrubium vulgare could be used as a potential antiviral agent.
PDF Abstract XML References Citation
Received: September 15, 2013;
Accepted: November 19, 2013;
Published: March 10, 2014
How to cite this article
Amal Fayyad, Nazlina Ibrahim and Wan Ahmad Yaacob, 2013. In vitro Virucidal Activity of Hexane Fraction of Marrubium vulgare
Against Type 1 Herpes simplex Virus. American Journal of Drug Discovery and Development, 3: 84-94.
DOI: 10.3923/ajdd.2013.84.94
URL: https://scialert.net/abstract/?doi=ajdd.2013.84.94
DOI: 10.3923/ajdd.2013.84.94
URL: https://scialert.net/abstract/?doi=ajdd.2013.84.94
INTRODUCTION
Herpes simplex virus (HSV-1) is a DNA virus, which is a causative agent for orofacial Herpes infection. It causes HSV encephalitis, meningitis and blinding keratitis particularly in immunocompromised patients (Cheng et al., 2004; Likhitwitayawuid et al., 2006; Akkarawongsa et al., 2008; Goldwich et al., 2011). Given the increasing number of immune-suppressed patients and the emerging acyclovir resistant strains of HSV-1, there is the need to discover drugs which have an alternative mode of action (Grover et al., 2011). Medicinal plants supply a diversity of chemical compounds with potential inactivation abilities on viruses (Astani et al., 2011; Ojo et al., 2009). In this study, we selected Marrubium vulgare (M. v) as a therapy for HSV-1. Marrubium vulgare is widely distributed throughout the Mediterranean region (Kanyonga et al., 2011). The essential oil of M. v has been demonstrated to produce a relaxant effect (El Bardai et al., 2003) in addition to exhibit antioxidant (Kadri et al., 2011) and antimicrobial properties (Zawislak, 2012; Kanyonga et al., 2011). A recent study reported that M. v contains compounds which vary in chemical composition, these include polysaccharides, polyphenolics, flavonoids, phenolics, tannins, carvacrol, triterpenes and anthraquinones (Zawislak, 2012). This present study was carried out to assess the mode of action of the hexane fraction on viral infection.
MATERIALS AND METHODS
Preparation of hexane fraction: Five grams leaves of M. v were extracted with 90 mL of methanol and water (2:1) by incubating for two days in a shaker at 37°C. The solution was then filtered by Whatman filter paper No. 1 and the methanol was removed under reduced pressure. Subsequently, the aqueous part was washed thrice with hexane and filtered. The hexane solvent was thereafter evaporated by a rotary evaporator and the residue collected by acetone before being left to dry.
Cells and virus: Vero cells were propagated in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 5% Fetal Bovine Serum (FBS), penicillin G (100 U mL-1) and streptomycin (100 μg mL-1) and non essential amino acids. Cell culture was maintained at 37°C under a humidified 5% CO2 atmosphere. Both Vero cells and HSV-1 were obtained from laboratory stocks.
Cytotoxicity assay: For evaluation of cytotoxicity, cells were seeded into 96-well plates and serial dilutions of the hexane fraction (200-70 μg mL-1) were added onto confluent cells in four replicates for each concentration of the drug. Cells treated with pure medium were used as controls. After 72 h of incubation, MTT solution [3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyl tetrazolium bromide was added and the plates were incubated for 3 h. After the MTT solution was removed, 130 μL of DMSO was added to each well to dissolve the formazan crystals. After gently shaking the plates, the observances were read on a multi-well spectrophotometer (BioRad Model 680, USA) at 540 nm. The CC50 was defined as the cytotoxic concentration of the fraction that reduced the observance of treated cells to 50% when compared with control.
Antiviral assay: A 12 well plate was cultivated with Vero cells (105 cell mL-1) and incubated for 24 h at 37°C. Monolayers were then infected with 100 PFU wel-1 of HSV-1. After 1 h of adsorption at 37°C in a 5% CO2 humidified atmosphere, the inocula was aspirated and 0.5 mL of medium 1% methyl cellulose with 100, 90 and 80 μg mL-1 was overlaid on the cell sheet. Each concentration was tested in triplicates. After infected cultures were incubated for 48 h at 37°C in a 5% CO2 incubator, cells were stained and dried. Viral plaques were counted and the percentage of virus reduction was calculated.
Determination of the mechanism of antiviral activity
Mechanism of virus inhibition: Hexane fraction was selected to study the mechanism of antiviral action on HSV-1 at different steps of the viral infection cycle. In all assays described, after the final incubation period the residual virus titers of the treated and untreated cell cultures were determined and expressed as percentage inhibition.
Cell pretreatment: Vero cell monolayers were treated with various non-toxic concentrations of hexane fraction ranging from 100-80 μg mL-1 and incubated for 24 h at 37°C. Vero cell line without treatment was used as a control. After removing the media, the pretreated cells were infected with HSV-1 (100 PFU well-1) and incubated for 2 h at 37°C, then the inocula removed and overlay medium was added. After two days, the number of plaques was counted and compared with the untreated viral infected cell controls (Yucharoen et al., 2011; Harden et al., 2009).
Penetration assay: Penetration assay was conducted followed procedures that have been described previously (Saddi et al., 2007; Kratz et al., 2008; Cheng et al., 2006; Hayashi et al., 1993) with minor modifications. Briefly, Vero monolayers grown in 12-well plates were prechilled at 4°C for 1 h. The medium was aspirated and the cell monolayer was infected with 100 PFU well-1 of HSV-1 and incubated at 4°C for 3 h. The medium was replaced with fresh medium, in the absence or presence of 100, 90 and 80 μg mL-1 from hexane fraction and the temperature was abruptly increased to 37°C to maximize penetration of the virus. Penetration proceeded forvarious time periods (30, 90 and 120 min, 1, 1.5 and 2 h). Then monolayers were treated with PBS at pH 3 for 1 min to neutralize any non-penetrated virus. After three washes with serum-free medium, cells were overlaid with DMEM-1% methyl cellulose and incubated for a further 48 h before the cell monolayer was fixed and stained. Plaques were counted and the percentage of inhibition of penetration was calculated by the following equation:
Virucidal assay: This assay followed procedures that have been previously described (Cheng et al., 2004; Saddi et al., 2007; Kratz et al., 2008; Shogan et al., 2006). Mixtures of hexane fraction and 1.0x106 PFU of HSV-1 in serum-free DMEM were incubated for 1 h at 37°C in 5% CO2 prior to the dilution of the mixture. The monolayers of cells were then infected with different dilutions (1x103, 1x104 and 1x105) of the mixture and incubated for 2 h at 37°C. The inocula were aspirated and overlay was added on infected cells and re-incubated for two days. The residual infectivity was determined by a viral plaque reduction assay.
Time-of-addition and time-of-removal assays: In the time-of-addition assay, monolayers of Vero cells grown in 12 well plates were infected with 100 PFU well-1 of HSV-1 for 2 h at 37°C in 5% CO2. At 2, 4, 6, 8, 10 and 12 h after infection, inocula were removed and 100 μg mL-1 of hexane fraction with 1% methyl cellulose was overlaidthe cell sheet and re-incubated 48 h. The percentage inhibition was calculated as the reduction in virus titre observed in infections containing the fraction compared with that of infections containing media only as a control.
In the time-of-removal assay, Vero cells were incubated with 100 PFU well-1 of HSV-1 for 2 h and immediately treated with 100 μg mL-1 of hexane fraction for 2, 4, 6, 8, 10, 12, 14, 16, 20, 24, 26, 30, 34, 38, 40, 44 and 48 h. The fraction was then removed and the cells overlaid with1% MC, then the culture was incubated up to 48 h after infection. The percentage inhibition was calculated as above (Cheng et al., 2004; Souza et al., 2007).
Transmission electron microscopy (TEM): For negative staining of the virions, the pellet of virions was collected from the infected cell supernatant and diluted with 200 μL DMEM. Half of this quantity was used as a control (virions only) and the remaining part was treated with hexane fraction. Five microlitter of each sample was deposited onto a formvar-carbon-coated copper grid followed by addition of 5 μL of uranyl acetate stain for 3 min. Excess suspension was drained using filter paper. The grids were observed by TEM. For thin-section TEM, the virion was collected from the infected cell supernatant with titer 1x108. The sample was divided into control (virion only) and the other half which was treated with hexane fraction. Both groups were fixed in 4% gluteraldehyde for 24 h at room temperature, washed with 0.1 M PBS, post-fixed with 1% osmium tetroxide for 2 h at 4°C, washed with 0.1 M PBS and dehydrated in a graded series of ethanol (35-100%). Subsequently, the samples were infiltrated with resin and acetone mixture at 1:1 for 1 h and at 3:1 for 2 h. This was kept in 100% resin over night, embedded with resin in beam capsules andpolymerized in the oven at 60°C for 48 h. The sections were cut between 60-90 nm by cryo Ultramicrotome and collected on copper grids. The grids were viewed with TEM.
Extraction of total cellular RNA: The total RNA was extracted using RNeasy mini kit and QIAshredder spin column as described by the manufacturer. The 5x106 cells were infected with or without 0.02 m.o.i. Herpes Simples Virus1 in the presence or absence of 100 μg mL-1 hexane fraction. After infection, Vero cells were collected at various times (4, 8, 16, 24 h p.i) and stored at 80°C. The samples were lysed with AVE-AVL, homogenized, washed with AW1 and AW2, eluted in RNAse-free water and then frozen in aliquots at-80°C. The concentration of the extracted RNA was calculated by measuring the optical density at 260 nm by nanodrop.
DNAse-treatment: The samples of RNA were thawed on ice, then genomic DNA was removed by Dnase1 RNase-free kit (Thermo Scientific). The total RNA 1 μg was incubated with 10xreaction buffer 1 μL with MgCl2, DNAse1-RNAse-free 1 μL and Rnase-free water at 37°C for 30 min. Subsequently, 1 μL of EDTA was added and incubated 10 min at 65°C.
cDNA synthesis: Viral RNA was reverse-transcribed using Maxima H Minus First Strand CDNA Synthesis kit (Thermo Scientific). The complementary DNA (CDNA) synthesis was carried out in a Thermal Cycler. The CDNA reaction (20 μL) consisted of total RNA (1 μg), random Hexamer primer (1 μL), dNTP (1 μL), 5xRT Buffer (4 μL), Maxima H Minus Enzyme Mix (1 μL) and RNase-free water. The reaction mixture was incubated for 10 min at 25°C, followed by 30 min incubation at 50°C. The reaction was terminated by heating at 85°C for 5 min, aliquoted and then frozen at -80°C.
Quantitative PCR (qPCR): The cDNA was diluted (1: 4), mixed with 2x master mix, primers and RNase-free water to 25 μL total volume. The mixture was then subjected to qPCR amplification in Q-5 biorad with a three-step cycling protocol (ThermoScientific No. k0382). The following primers were used against HSV-1 immediate-early (ICP27), forward primer, 5'-GCCGCGACGACCTGGAATCG-3'; reverse primer, 5'-TGTGGGGCGCTGGTTGAGGATC-3', for early (TK), forward primer, 5'-AGGTATCGCGCGGCCGGGTAG-3'; reverse primer, 5'-ATGGCTTCG TACCCCTGCCA-3', for late (gD) forward primer, 5'-ATGGGAGGCAACTGTGCTATCC-3'; reverse primer, 5'-CTCGGTGCTCCAGGATAAAC-3', genes and for the cellular cyclophilin A forward primer, 5'-ATGGCACTGGTGGAAGTC-3', reverse primer, 5'-ATTCCTGGACCCAAAGCG-3' gene.
Data analysis: The 50% Cytotoxicity Concentration (CC50) was defined as the test hexane concentration required for a reduction of cell viability by 50%. CC50 values were calculated using regression analysis. The mean number of plaques was used to calculate the percentage of plaques inhibited. A curve relating percent plaque inhibition against extract concentrations was used to calculate the inhibitory concentration that elucidated a 50% reduction of plaque formation (IC50). The gene expression was calculated with one way ANOVA and T-test.
RESULTS
Toxicity study on the hexane fraction indicated no toxic effect on the cells at concentrations less than 140 μg mL-1 as shown in Fig. 1.
The CC50, IC50 and the therapeutic index TI (CC50/IC50) of the hexane fraction against HSV-1 were 140, 50 and 2.8, respectively.
Microscopic examination of cell monolayers after 3 days of incubation with hexane fraction at <140 μg mL-1 showed no changes in morphological shape compared with the control, therefore, hexane fraction was used at concentrations <140 μg mL-1 in successive studies.
The inhibitory effect of hexane fraction was examined after its addition at different time periods relative to virus infection. The results showed that various concentrations of hexane fraction added at 24 h prior to virus infection or post penetration did not have antiviral activity. However, the addition of hexane fraction out at 1 h post infection inhibited the virus in a dose-dependent manner as shown in Fig. 2.
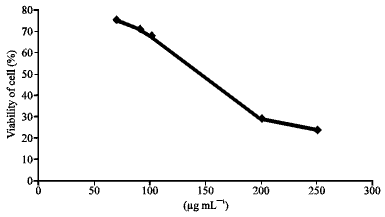 |
| Fig. 1: | Effect of hexane fraction on vero cells |
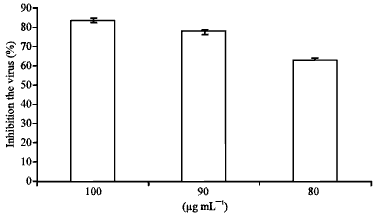 |
| Fig. 2: | Antiviral activity of hexane fraction on HSV-1 |
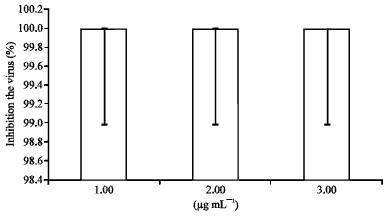 |
| Fig. 3: | Virucidal activity of hexane fraction on HSV-1 |
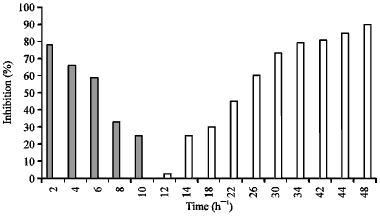 |
| Fig. 4: | Effect of hexane fraction on HSV-1 at time of addition and time of removal (2-10 h, the time of addition. 12-48 h time of removal) |
To verify that the hexane fraction had virucidal activity, the hexane fraction was incubated with the virus for 1 h. As shown in Fig. 3 the plaque formation was completely inhibited when the virion was treated with hexane fraction.
These results indicated that hexane fraction inactivated the virus directly with potent virucidal effect against HSV-1. These results is in agreement with the results obtained by TEM negative Staining and thin sections of the virion treated directly with hexane fraction (Fig. 5a-d). On the other hand, untreated control HSV-1 samples showed whole, undisrupted virion. The glycoprotein spikes were seen as completely intact with the complete envelope. In contrast, for the hexane fraction-treated samples, virion particles observed in the grid sections contained viruses which were partially disrupted.
The inhibition of plaque formation was observed when the hexane fraction was added at between 1 and 6 h post virus infection. Figure 4 shows that the inhibition was 80, 66, 59, 33 and 25% when the hexane fraction was added after 2, 4, 6, 8 and 10 h post-infection, respectively. In contrast, the addition of the drug after 12 h post-infection failed to show any inhibition of the virus. On the other hand, in the time of removal assay (Fig. 4), the inhibition of virus concentration started at 12 h and increased gradually. The inhibition obviously increased at 30 h and reached to a maximum at 48 h.
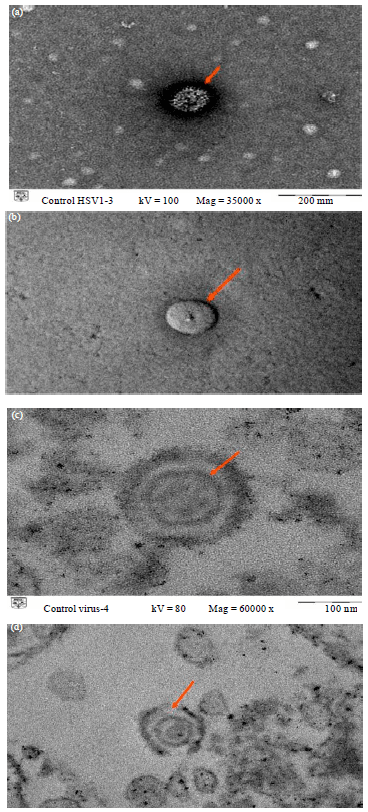 |
| Fig. 5(a-d): | (a) Control virion of HSV-1 by negative staining (red arrow), (b) Treated virion with hexane fraction by negative staining (red arrow), (c) Resin-embedded section of control HSV-1 (red arrow) and (d) Resin-embedded section of treated HSV-1 with hexane fraction (red arrow) |
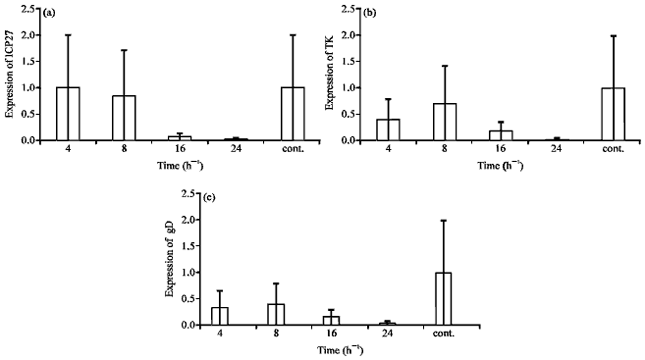 |
| Fig. 6(a-c): | (a) Effect of hexane fraction on ICP27 expression, (b) Effect of hexane fraction on TK expression and (c) Effect of hexane fraction on gD expression |
To further analyze the mechanisms of hexane fraction, the total cellular RNA was collected at various times after viral adsorption. Our results indicated that the levels of (ICP27), (TK) and (gD) m RNA expression gradually decreased as shown in (Fig. 6a-c) and the highest inhibition was at 24 h post infection. The hexane fraction significantly reduced the expression of UL54 (ICP27), UL23 (TK) and US6 (gD) (p<0.05).
DISCUSSION
Marrubium vulgare is a natural plant that grows in the Mediterranean and is used to treat infections of skin. The extract and leaves of M. v are rich in many compounds like terpenes and anthraquinone, which have anti-inflammatory, vasorelaxant (El Bardai et al., 2003), anti-diabetic (Boudjelal et al., 2012), antioxidant (Kadri et al., 2011), anti-inflammatory, analgesic, anti-bacterial (Masoodi et al., 2008; Kanyonga et al., 2011) and antioedematogenic (Stulzer et al., 2006) activities. Here we further extended studies on M. v hexane fraction by investigating its antiviral properties and mechanism of action.
Assessment of cytotoxicity is obviously an important part of the estimation of the potential applicability of any antiviral agent. The goal is to have an antiviral agent with potent action on the selected target while demonstrating little or no effect on cellular function. In this study, it was observed that the vero cells treated with hexane fraction did not exhibit altered morphology or growth characteristics indicative of cytotoxic effects. In antiviral studies performed with M. v hexane fraction, we found that there was no inhibition even with pretreatment of the cells prior to infection. This indicates that the hexane fraction has no effect on the cell receptors. Also, following the penetration of HSV-1 into the cell, there was no inhibition in the plaque formation as compared with the control. The fraction takes a long time to penetrate the cell, so the virus enters before it is affected by the fraction. In the resting tests, complete inhibition (100%) of the virion of HSV-1 was observed when free virus was directly exposed to the hexane fraction in the virucidal assay. It was observed that treatment with hexane fraction at 2 h post-infection could inhibit HSV-1 infection, although the addition of the fraction at 8 h post-infection resulted in a relatively weaker inhibitory effect. The removal of fraction from 2-10 h p.i, revealed no inhibition to the virus but the reduction in plaque formation started from 12-48 h. A further effect of hexane fraction on the expression levels of HSV-1 IE, E and L genes was observed. As the results showed, the expression of IE, E and L genes were significantly reduced. It is possible that the fraction might affect the expression of UL54 (ICP27) gene thus blocking the subsequent expressions of UL23 (TK) and US6 (gD) genes. Consequently, the virucidal activity of hexane fraction is more likely to be attributed to its ability to interact with HSV-1 envelope. The hexane fraction disintegrated the HSV-1 envelope and this was verified by TEM suggesting that hexane fraction interferes with the virion and prevents it from entering the cell. This finding is supported by previous studies which reported on the mechanism of extracts on the virus as being by the solubilization of the virus envelope; or the chemical modification, degradation, or masking of some of the essential envelope proteins (Kratz et al., 2008; Carson et al., 2006). Also showed that terpenes derived from plants have been shown to have antiviral activity. Monoterpenes, sesquiterpenes and their associated alcohols are found to be virucidal against HSV-1. The antiviral activity of these substances is the greatest on free virus. Previous studies revealed that the highest concentration of 3% terpenes reduced the virus titer by 98.2%, an observation consistent with our findings. M. v is known to contain amounts of terpenes and anthraquanines, thus these substances in hexane fraction are likely individual or synergist contributors to the antiviral effects observed in the study. Another study reported that essential oils containing1% terpenes inhibited plaque form completely while 0.1% terpenes reduced plaque formation by approximately 10%. However, pretreatment the Vero cells with 0.1% terpenes before infection with the virus did not inhibit plaque formation. This is understandable considering the lipophilic nature of the oil which enables it to penetrate the virion envelope (Duschatzky et al., 2005; Jaime et al., 2013). In another study, the application of essential oils of Melissa officinalis from Lamiaceae for the treatment of HSV-2 has been reported recently (Allahverdiyev et al., 2004; Camargo Filho et al., 2008). Subsequent studies of anthraquanines effect on the HSV-1 envelope revealed that the anthraquinones acted directly on the envelope of HSV-1, thus the adsorption and replication of the virus rundown (Sydiskis et al., 1991; Cohen et al., 1996; Zandi et al., 2007). Hsiang and Ho (2008) investigated the action of emodin on HSV-1. It was observed in the study that emodin possesses antiviral activities which are elucidated in the form of inhibition of both UL12 and CK2 as well as in the disruption of the phospholipid bilayer of the envelope.
In summary, our findings suggest that the hexane fraction with its gene expression inhibitions and rapid virucidal activities could be an effective topical treatment for oral herpetic infections. These findings however emphasize the need for future studies that would isolate and identify the bioactive anti-HSV molecules in hexane fraction and determine whether the bioactive standard in hexane fraction exhibits antiviral effects against additional HSV strains.
ACKNOWLEDGMENTS
This study was supported by Universiti Kebangsaan Malaysia (BKBP-FST-K006401).
REFERENCES
- Akkarawongsa, R., T.B. Potocky, E.P. English, S.H. Gellman and C.R. Brandt, 2008. Inhibition of herpes simplex virus type 1 infection by cationic β-peptides. Antimicrob. Agents Chemother., 52: 2120-2129.
CrossRefDirect Link - Allahverdiyev, A., N. Duran, M. Ozguven and S. Koltas, 2004. Antiviral activity of the volatile oils of Melissa officinalis L. against Herpes simplex virus type-2. Phytomedicine, 11: 657-661.
CrossRefPubMedDirect Link - Astani, A., J. Reichling and P. Schnitzler, 2011. Screening for antiviral activities of isolated compounds from essential oils. Evid. Based Complement. Alternat. Med.
CrossRef - Boudjelal, A., C. Henchiri, L. Siracusa, M. Sari and G. Ruberto, 2012. Compositional analysis and in vivo anti-diabetic activity of wild Algerian Marrubium vulgare L. infusion. Fitoterapia, 83: 286-292.
CrossRefDirect Link - Camargo Filho, I., D.A.G. Cortez, T. Ueda-Nakamura, C.V. Nakamura and B.P. Dias Filho, 2008. Antiviral activity and mode of action of a peptide isolated from Sorghum bicolor. Phytomedicine, 15: 202-208.
CrossRefDirect Link - Carson, C.F., K.A. Hammer and T.V. Riley, 2006. Melaleuca alternifolia (tea tree) oil: A review of antimicrobial and other medicinal properties. Clin. Microbiol. Rev., 19: 50-62.
CrossRefDirect Link - Cheng, H.Y., T.C. Lin, C.M. Yang, K.C. Wang, L.T. Lin and C.C. Lin, 2004. Putranjivain A from Euphorbia jolkini inhibits both virus entry and late stage replication of herpes simplex virus type 2 in vitro. J. Antimicrob. Chemother., 53: 577-583.
CrossRef - Cheng, H.Y., C.M. Yang, T.C. Lin, D.E. Shieh and C.C. Lin, 2006. ent-Epiafzelechin-(4α→8)-epiafzelechin extracted from Cassia javanica inhibits herpes simplex virus type 2 replication. J. Med. Microbiol., 55: 201-206.
CrossRef - Cohen, P.A., J.B. Hudson and G.H.N. Towers, 1996. Antiviral activities of anthraquinones, bianthrones and hypericin derivatives from lichens. Experientia, 52: 180-183.
CrossRef - Duschatzky, C.B., M.L. Possetto, L.B. Talarico, C.C. Garcia and F. Michis et al., 2005. Evaluation of chemical and antiviral properties of essential oils from South American plants. Antiviral Chem. Chemother., 16: 247-251.
Direct Link - El Bardai, S., N. Morel, M. Wibo, N. Fabre, G. Llabres, B. Lyoussi and J. Quetin-Leclercq, 2003. The vasorelaxant activity of marrubenol and marrubiin from Marrubium vulgare. Planta Med., 69: 75-77.
Direct Link - Goldwich, A., A.T. Prechtel, P. Muhl-Zurbes, N.M. Pangratz and H. Stossel et al., 2011. Herpes simplex virus type I (HSV-1) replicates in mature dendritic cells but can only be transferred in a cell-cell contact-dependent manner. J. Leukoc. Biol., 89: 973-979.
CrossRef - Grover, A., V. Agrawal, A. Shandilya, V.S. Bisaria and D. Sundar, 2011. Non-nucleosidic inhibition of Herpes simplex virus DNA polymerase: Mechanistic insights into the anti-herpetic mode of action of herbal drug withaferin A. BMC Bioinform.
CrossRef - Harden, E.A., R. Falshaw, S.M. Carnachan, E.R. Kern and M.N. Prichard, 2009. Virucidal activity of polysaccharide extracts from four algal species against herpes simplex virus. Antiviral Res., 83: 282-289.
CrossRefDirect Link - Hayashi, K., T. Hayashi, N. Morita and I. Kojima, 1993. An extract from Spirulina platensis is a selective inhibitor of herpes simplex virus type 1 penetration into HeLa cells. Phytother. Res., 7: 76-80.
CrossRef - Jaime, M.F.V., F. Redko, L.V. Muschietti, R.H. Campos, V.S. Martino and L.V. Cavallaro, 2013. In vitro antiviral activity of plant extracts from Asteraceae medicinal plants. Virol. J., Vol. 10.
CrossRef - Kadri, A., Z. Zarai, A. Békir, N. Gharsallah, M. Damak and R. Gdoura, 2011. Chemical composition and antioxidant activity of Marrubium vulgare L. essential oil from Tunisia. Afr. J. Biotechnol., 10: 3908-3914.
CrossRefDirect Link - Kanyonga, P.M., M.A. Faouzi, B. Meddah, M. Mponal, E.M. Essassi and Y. Cherrah, 2011. Assessment of methanolic extract of Marrubium vulgare for antiinflammatory, analgesic and anti-microbiologic activities. J. Chem. Pharm. Res., 3: 199-204.
Direct Link - Kratz, J.M., C.R. Andrighetti-Frohner, D.J. Kolling, P.C. Leal and C.C. Cirne-Santos et al., 2008. Anti-HSV-1 and anti-HIV-1 activity of gallic acid and pentyl gallate. Mem. Inst. Oswaldo. Cruz., 103: 437-442.
CrossRef - Likhitwitayawuid, K., S. Chaiwiriya, B. Sritularak and V. Lipipun, 2006. Antiherpetic flavones from the heartwood of Artocarpus gomezianus. Chem. Biodiversity, 3: 1138-1143.
CrossRef - Masoodi, M.H., B. Ahmed, I.M. Zargar, S.A. Khan, S. Khan and P. Singh, 2008. Antibacterial activity of whole plant extract of Marrubium vulgare. Afr. J. Biotechnol., 7: 86-87.
Direct Link - Ojo, O.O., J.O. Oluyege and O. Famurewa, 2009. Antiviral properties of two Nigerian plants. Afr. J. Plant Sci., 3: 157-159.
Direct Link - Saddi, M., A. Sanna, F. Cottiglia, L. Chisu, L. Casu, L. Bonsignore and A. de Logu, 2007. Antiherpevirus activity of Artemisia arborescens essential oil and inhibition of lateral diffusion in Vero cells. Ann. Clin. Microbiol. Antimicrob.,.
CrossRef - Shogan, B., L. Kruse, G.B. Mulamba, A. Hu and D.M. Coen, 2006. Virucidal Activity of a GT-Rich Oligonucleotide against Herpes Simplex Virus Mediated by Glycoprotein B. J. Virol., 80: 4740-4747.
PubMed - Souza, T.M.L., J.L. Abrantes, R.A. Epifanio, C.F.L. Fontes and I.C.P.P. Frugulhetti, 2007. The alkaloid 4-methylaaptamine isolated from the sponge Aaptos aaptos impairs Herpes simplex virus type 1 penetration and immediate-early protein synthesis. Planta Med., 73: 200-205.
CrossRef - Stulzer, H.K., M.P. Tagliari, J.A. Zampirolo, V. Cechinel-Filho and V. Schlemper, 2006. Antioedematogenic effect of marrubiin obtained from Marrubium vulgare. J. Ethnopharmacol., 108: 379-384.
CrossRefDirect Link - Sydiskis, R.J., D.G. Owen, J.L. Lohr, K.H. Rosler and R.N. Blomster, 1991. Inactivation of enveloped viruses by anthraquinones extracted from plants. Antimicrob. Agents Chemother., 35: 2463-2466.
PubMedDirect Link - Yucharoen, R., S. Anuchapreeda and Y. Tragoolpua, 2011. Anti-herpes simplex virus activity of extracts from the culinary herbs Ocimum sanctum L., Ocimum basilicum L. and Ocimum americanum L. Afr. J. Biotechnol., 10: 860-866.
Direct Link - Zandi, K., M.A. Zadeh, K. Sartavi and Z. Rastian, 2007. Antiviral activity of Aloe vera against herpes simplex virus type 2: An in vitro study. Afr. J. Biotechnol., 6: 1770-1773.
Direct Link