
V. Pooja
Amity Institute of Biotechnology, Amity University, J-3 Block, Sector 125, Noida, India
LiveDNA: 91.3299
Hina Sanwal
Amity Institute of Biotechnology, Amity University, J-3 Block, Sector 125, Noida, India
Seema Bhatnagar
Amity Institute of Biotechnology, Amity University, J-3 Block, Sector 125, Noida, India
Ashwani K. Srivastava
Amity Institute of Biotechnology, Amity University, J-3 Block, Sector 125, Noida, India
American Journal of Drug Discovery and Development
Year: 2012 | Volume: 2 | Issue: 1 | Page No.: 32-39







ABSTRACT
Myristica fragrans HOUTT has been traditionally used by Asian Indians to treat stomach pains, dysentery, vomiting and the symptoms of malaria. It is also chewed to prevent foul breath. The present study was carried out to elucidate the antifungal effect of four extracts; hexane, chloroform, methanolic and ethanolic extract obtained from Mace. The extracts were concentrated and re-suspended in DMSO. MIC80 was calculated by micro broth dilution method. Results have revealed the extracts to have good antifungal activity against Aspergillus niger and methanol and hexane extracts have been found to be most effective. Methanol and hexane extracts were also associated with de-melanization of Aspergillus niger. Further analysis revealed difference in the structure of the hyphal cell wall of methane and hexane extract treated cells.
PDF Abstract XML References Citation
Received: November 04, 2011;
Accepted: January 16, 2012;
Published: February 13, 2012
How to cite this article
V. Pooja, Hina Sanwal, Seema Bhatnagar and Ashwani K. Srivastava, 2012. Targeting Virulence: Novel Effect of Myristica fragrans on Melanisation and Conidiation of Aspergillus niger. American Journal of Drug Discovery and Development, 2: 32-39.
DOI: 10.3923/ajdd.2012.32.39
URL: https://scialert.net/abstract/?doi=ajdd.2012.32.39
DOI: 10.3923/ajdd.2012.32.39
URL: https://scialert.net/abstract/?doi=ajdd.2012.32.39
INTRODUCTION
Several species of pathogenic bacteria, fungi and helminths produce melanin which has been associated with virulence in many microorganisms making it an important factor in the pathogenesis. The complete biochemical and structural analysis of melanin is difficult making its study a challenging task (Nosanchuk and Casadevall, 2003). Melanin generally contributes to virulence by reducing the susceptibility of melanised microbes to host defence mechanisms. The polyketide biosynthetic pathway in ascomycetes and related deuteromycetes produces 1,8-dihydroxynaphthalene (DHN) that is a precursor of most fungal melanins known as DHN-melanins. Few human pathogens which form melanin precursors by the polyketide pathway are Aspergillus nidulans, Aspergillus niger, Alternaria alternata, Cladosporium carrionii, Exophiala jeanselmei, Fonsecaea compacta, Hendersonula toruloidea, Phaeoannellomyces werneckii, Phialophora richardsiae, Wangiella dermatitidis and Xylohypha bantiana (Jacobson, 2000).
Uninucleate, highly pigmented spores called the conidia which are mitotically derived, are responsible for the propagation of Aspergillus species and this is a common method of asexual reproduction. The morphological transition of vegetative hyphae to highly specialised spore bearing structures (conidiophores) is under stringent genetic regulation. Most of the data available so far on conidiation are based on the studies conducted on A. nidulans.
Myristica fragrans belongs to family Myristicaceae is native to the Banda Islands in Eastern Indonesia (Moluccas) and is cultivated in the Banda Islands, Brazil Grenada, the Caribbean, Malaysia, South India, Sri Lanka and Sumatra. It is spicy and bitter with clove like and piney overtones. The phytochemical analysis of M. fragrans shows the presence of essential oil in the range of 7 to 14% and about 30% fixed oil. It also contains 87.5% monoterpenes, 5.5% monoterpene alcohols and 7% other aromatics. Like nutmeg essential oil, the main constituents of mace essential oil are α-pinene, 1,8-cineole, myrcene, myristicin, limonene, terpinen-4-ol, γ-terpinene, sabinene and safrole (Pooja et al., 2012).
In the present study, the effect of M. fragrans on conidiation and melanisation of A. niger was investigated.
MATERIALS AND METHODS
Spices collection and identification: The spices used in this study were procured from local Indian markets and were later authenticated in the Botany Department of Amity University, NOIDA.
Preparation of the plant extract: The spice was cleaned under running tap water and dried. In order to obtain the spice extract about 10 g of spice were crushed with mortar and pestle and sieved. The dried powder was then extracted with 40 mL n-Hexane, Chloroform, Methanol and Ethanol consecutively for 72 h/solvent under constant stirring. The extract was then filtered and dried under reduced pressure and resuspended in DMSO (Krishnan et al., 2010).
Organisms: A. niger (ATCC 16404) used in this study was obtained from Amity Institute of Microbial Technology. The A. niger culture was sub cultured on Potato Dextrose Agar medium. Working cultures were subsequently maintained on Potato dextrose agar slants at 4°C. Long-term preservation of the cultures was done in 15% glycerol as conidial suspension at -80°C.
Minimum inhibitory concentration: Determination of the Minimum Inhibitory Concentration (MIC80) was carried out for the extracts that showed inhibitory effect on the test micro-organism. MIC was conducted based on microdilution method with minor modification in a 96 well plate according to NCCLS guidelines. To the first well 180 μL of fresh YPD media was added with 20 μL of the extract. The well contents were mixed and serially diluted till the 10th well. One hundred microliter of fresh conidial culture were then added to all wells except 11th well which were kept as a media control. The final concentration of the conidial culture was kept at 104 cells/well. Two hundred microliter of the fresh media were added to the 11th well and 100 μL of Candida+100 μL of the media were added in the 12th well which were kept as (+) control. The plate was incubated for 7 days at 30°C. The plate was then read visually and spectroscopically by ELISA plate reader. The MIC80 was defined as the concentration of the drug that produced 80% growth inhibition (NCCLS-National Committee for Clinical Laboratory Standards, 1998).
Inhibition of conidiation: The inhibition of conidiation in A. niger by different extracts of M. fragrans by plating the germinated conidias of A. niger on PDA plates. The conidias in the concentration of 104 were plated on PDA plates containing extracts in sub MIC concentration. The concentration was kept 1 to 10 times less than the MIC80 concentration. The plates were incubated at 30°C for 7 days to obtain fungal colonies. From the centre of the 7-day old A. niger colonies agar blocks (1 cm in diameter) were excised using a sterile cork borer. The conidia from the agar blocks were suspended in sterile distilled water (4 mL per agar block) by vigorous agitation and the conidial density (number of conidia per colony) was determined by haemocytometry.
Microscopic analysis of the A. niger colony: Approximately 1x105 conidia were spread on potato dextrose agar plates in duplicate containing hexane extract (0.068 mg mL-1), chloroform extract (0.275 mg mL-1), methanol extract (0.054 mg mL-1) and ethanol extract (0.118 mg mL-1). The concentration of the extract in each case was 8 times less than its MIC concentration. Fungal cultures were allowed to grow for 7 days at 30°C and then examined visually for white colonies and microscopically for the presence of conidia using lactophenol cotton blue staining. The colony morphology and hyphal cell wall pigmentation were recorded photographically using a Nikon digital camera.
Inhibition of conidiation in liquid culture: The 1x105 germinated conidias were grown in 100 mL of potato dextrose broth at 30°C. The test culture broth was added with the extract (8 times less than MIC) and the control broth. The liquid culture was incubated at 30°C for 7 days. Results were noted.
Estimation of ergosterol content of the cell wall of A. niger: The A. niger culture in a concentration of 1x104 conidias mL-1 were inoculated in 100 mL potato dextrose broth in the presence of sub-minimum inhibitory concentration of the extract. The culture was allowed to grow at 30°C for 5 days under gentle agitation. After 5 days the mycelium were harvested. The mycelia were weighed and hydrolysed with 1% SDS (100 mL/10 g of mycelium). The mycelia were dehydrated with absolute Ethanol and dried. The dried cell wall was suspended in distilled water and an equal volume of CHCl3 and CH3OH (1:2) was added. The CHCl3 layer was separated. The cell wall was re-extracted with a mixture of CHCl3 and water (1:1) in a separating funnel. The Chloroform layer was then transferred in a watch glass and evaporated to dryness. The dry mass was measured and resuspended in Ethanol. The % ergosterol was calculated by Young et al. (2003).
Ergosterol % = [(A281.5/A290 x F)/pellet wt]-[(A230/A518 x F/pellet wt] |
where, F is ethanol dilution factor
Statistical analysis: Paired t-test values were calculated for all the extracts for assessment of fold reduction in ergosterol assay using SAS (version 9.2). The p-values of = 0.005 were considered highly significant.
RESULT
In vitro susceptibility test: We investigated the in vitro susceptibility of A. niger ATCC 16404 to different extracts of M. fragrans by the broth microdilution method (Table 1). It was found that A. niger was susceptible to all four extracts. The microorganism was found to be highly susceptible to Methanolic extract (MIC 0.432 mg mL-1), Hexane extract showed an MIC of 0.547 mg mL-1. The microorganisms were moderately susceptible to Chloroform extract (MIC80 : 2.2 mg mL-1).
| Table 1: | Minimum inhibitory concentration of the extracts of Myristica fragrans against A. niger. the MIC was determined by Micro broth dilution assay in a 96 well plate according to the NCCLS guideline |
 | |
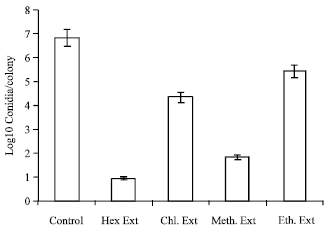 |
| Fig. 1: | Effect of various extracts of M. fragrans on conidiation of A. niger. The inhibition of conidiation was determined using the agar block conidiation assay. The results show the conidial count at an extract concentration of 8<MIC. Each histogram represents the mean of two independent determinations |
 |
| Fig. 2: | PDA plates showing A. niger colonies grown on PDA plates in the presence of extracts of M. fragrans. Plate 1: Control Plate (no extract), Plate 2: Hexane extract, Plate 3: Chloroform extract, Plate 4: Methanol extract, Plate 5: Ethanol extract |
Inhibition of conidiation: Figure 1 shows the effect of a sub-inhibitory concentration of the extracts on conidiation and production of melanin in A. niger. Cultures of A. niger were grown from germinated conidia on Potato dextrose agar in the presence of sub MIC concentration (8 times less) of each extract.
It was observed that the colonies grown in the presence of hexane and methanolic extracts were white in colour as compared with the control cultures grown in the absence of the extract. Microscopic examination of the white colonies showed almost complete absence of conidia in the case of A. niger in hexane extract and nearly 80% inhibition of conidiation in A. niger in the presence of methanolic extract (Fig. 2).
Result indicates reduced conidiation of the A. niger colonies when grown in the presence of hexane and methanol extract. Slightly reduced conidiation (25%) is seen in case of A. niger colonies when grown in the presence of chloroform and ethanolic extracts.
The conidial shape and size were also different in the presence of the test extract. The conidias were also grown in liquid cultures. The results were similar to the results achieved on PDA plates the Liquid culture also showed extreme reduction of conidiation in hexane extracts and substantial reduction in methanolic extracts as compared to the control. Ethanol and chloroform extracts showed reduction of only 25% (Fig. 3).
 |
| Fig. 3: | Liquid culture of 1 x 104 A. niger is grown in the presence of Sub-MIC concentration of hexane and methanolic extracts of M. fragrans. When compared with A. niger colonies of the control extreme reduction in the conidial growth can be seen in a 7 day old culture |
 |
| Fig. 4: | Hyphal cell wall analysis under 40X compound microscope was done for the A. niger colonies grown in hexane extracts (white colonies) and under control condition (black colonies). The cell wall from the hyphae of the A. niger colony grown on control plate is observed having a dark coloured outer membrane. The cell wall from the hyphae of the A. niger colony grown on test plate having hexane extract is observed having transparent outer membrane |
| Table 2: | The % fold reduction in the ergosterol content of hyphal cell wall of A. niger |
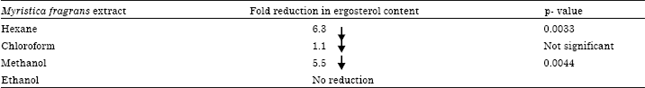 | |
The ergosterol content of the hyphal cell wall was estimated spectrophotometrically and % ergosterol was calculated by % ergosterol = [(A281.5/A290 x F)/pellet wt]-[(A230/A518 X F/pellet wt], where F is ethanol dilution factor. Result indicate fold reduction in the ergostrerol content in the hyphal cell wall grown in the presence of hexane>methanol>chloroform as compared to the control (no extract). No reduction was seen in the A. niger colonies grown in ethanolic extract. The p-value was found to significant in case of hexane extract (0.0033) and methanol extract (0.0044) | |
The hyphal cell wall analysis showed difference in the structure of the cell wall. The control cell wall showed a black region though out the wall were as the hyphae of the A. niger grown in the presence of the extract is white in colour. This may be due to decrease in the melanin content of the hyphal cell wall (Fig. 4).
Estimation of ergosterol: The ergosterol content of the hyphal cell wall was estimated by spectrophotometric analysis and calculating the % ergosterol content of the A. niger grown in the presence of the extracts of M. fragrans. The control was taken as the A. niger grown in the absence of the any extract. The difference was given as % fold reduction in the ergosterol content of the test as compared to the control tubes (Table 2).
DISCUSSION
Medicinal plants have been used since time immemorial to treat various infectious diseases. These studies have been carried out in various ethnic groups, with plants from various regions (Dulger and Gonuz, 2004; Bonjar et al., 2004; Saadabi, 2006; Motamedi et al., 2010; Anam et al., 2010; Bahrami and Ali, 2010). In the present study we have demonstrated the antifungal activity of M. fragrans Houtt. We also further demonstrated that this effect is mainly due to the reduction in melanisation of A. niger conidia and hyphae by M. fragrans extracts (Pooja et al., 2012). Hexane and methanol extracts had very low MIC values compared to the chloroform and ethanol extracts. This result corroborated with present finding that hexane and methanol extracts reduce conidiation maximally. When the treated hyphae were observed under the microscope the cell wall of the treated hyphae were deficient in melanin content leading us to the conclusion that the antifungal activity of M. fragrans is due to reduction of conidia formation and also due to reduction in the melanin content of the cells.
Over the past few years, the incidence of life threatening fungal infections have dramatically increased particularly in immune-compromised patients. The major risk factors in most of the cases of fungal disease include HIV infections, Administration of broad spectrum antibiotics, intravenous catheters and other implants etc. Unfortunately, there has not been any major development in anti fungal therapeutics and amphotericin B remains the drug of choice in most of the cases. The major drawbacks associated with the use of polenes include poor solubility at biological pH, increased toxicity and low bioavailability (Lokhande et al., 2006; Sarwar et al., 2011).
Melanin synthesis has been associated with virulence for a variety of pathogenic microbes. Melanin and melanin synthesis pathways are potential targets for antimicrobial drug discovery since melanin is believed to contribute to microbial virulence by reducing a pathogen's susceptibility to killing by host antimicrobial mechanisms and by influencing the host immune response to infection. Interestingly, the drug-binding properties of both host and pathogen melanins could influence the efficacy of an antimicrobial agent (Nosanchuk et al., 1999).
Melanin deficient C. neoformans mutant strains which were generated by UV- irradiation when injection into murine model were found to be a virulent. Reversion to virulence was associated with recovery of melanin production. It was seen in a study that low melanin producing strain induced TNF-α production in vitro in contrast high melanin containing strain inhibited TNF-α production and lymphoproliferation. The study demonstrated that melanin can inhibit the recognition of the organisms by host defences, thereby down regulating the afferent phase of T-cell mediated immunity (Huffnagle et al., 1995).
The localization of melanin in the fungal cell wall is supported by three studies. First, removal of the cell wall removes most of the dark colour. Second, cell walls of albino mutants of W. dermatitidis and C. neoformans appear hyaline in electron micrographs, whereas those of the parental wild types have an electron-dense outer layer. Feeding of scytalone to the albino culture of W. dermatitidis allowed the mutant to become melanized and re-established the electron-dense layer (Polak and Dixon, 1989).
It is possible to find examples of indifference, apparent protection and apparent potentiation when melanized and nonmelanized cells are exposed to various drugs. Melanin did not protect W. dermatitidis against antifungal drugs, since Mel¯ mutants were no more susceptible to a variety of antifungals than was the Mel+ wild type. However, the opposite conclusion was drawn with C. neoformans. When catechols were withheld, thereby preventing melanization, cryptococcal cultures survived a 1-h treatment with amphotericin B less well (Polak and Dixon, 1989).
The development of drugs that interfere with melanin polymerization or rearrangement may be useful therapeutic compounds for the treatment of these melanotic fungi and other pathogens that produce melanin. Also, it is possible that the use of agents that inhibit melanization may render melanotic fungi susceptible to drugs that bind to melanin. An interesting finding is the fact that voriconazole at 0.125 to 0.5 mg L-1 can inhibit conidiation in diverse Aspergillus spp., resulting in white colonies. Ravuconazole which is structurally similar to voriconazole, had similar effects only against Aspergillus fumigatus and Aspergillus flavus. It is possible that the inhibition of melanin formation in vivo may contribute to the therapeutic potencies of these triazoles by increasing the susceptibility to host defence mechanisms. The possibility that certain antifungal agents are less effective against melanotic molds should especially be considered when clinicians make choices for empirical therapy in patients with presumed mycotic diseases (Nosanchuk and Casadevall, 2006). Further prospects include combinatorial studies of M. fragrans extracts with drugs such as Voriconazole which are known antifungals which affect the melanisation of Aspergillus species. The present study paves the way for development of an effective antifungal which will not elicit any side effect since M. fragrans is routinely used as a dietary spice.
ACKNOWLEDGMENTS
The authors would like to thank Dr. Rajni Singh of Amity Institute of Microbial Biotechnology for providing ATCC 16404 A. niger used in this study.
REFERENCES
- Anam, K., A.G. Suganda, E.Y. Sukandar and L.B.S. Kardono, 2010. Antibacterial effect of component of Terminalia muelleri Benth. against Staphylococcus aureus. Int. J. Pharmacol., 6: 407-412.
CrossRef - Bahrami, A.M. and V. Ali, 2010. Effects of Scrophularia striata ethanolic leaves extracts on Staphylococcus aureus Int. J. Pharmacol., 6: 431-434.
CrossRefDirect Link - Dulger, B. and A. Gonuz, 2004. Antimicrobial activity of certain plants used in turkish traditional medicine. Asian J. Plant Sci., 3: 104-107.
CrossRefDirect Link - Jacobson, E.S., 2000. Pathogenic roles of fungal melanins. Clin. Microbiol. Rev., 13: 708-717.
Direct Link - Huffnagle, G.B., G.H. Chen, J.L. Curtis, R.A. McDonald, R.M. Strieter and G.B. Toews, 1995. Down-regulation of the afferent phase of T cell mediated pulmonary inflammation and immunity by a high melanin producing strain of Cryptococcus neoformans. J. Immunol., 155: 3507-3516.
Direct Link - Nosanchuk, J.D. and A. Casadevall, 2003. The contribution of melanin to microbial pathogenesis. Cell Microbiol., 5: 203-223.
PubMed - Krishnan, N., S. Ramanathan, S. Sasidharan, V. Murugaiyah and S.M. Mansor, 2010. Antimicrobial activity evaluation of Cassia spectabilis leaf extracts. Int. J. Pharmacol., 6: 510-514.
CrossRefDirect Link - Lokhande, P.D., K.R. Gawai, K.M. Kodam, B.Y. Waghmare, A.R. Chabukswar and S.C. Jagdale, 2006. Water soluble amide derivatives of polyene antibiotic and their antifungal activity. Trends Applied Sci. Res., 1: 529-533.
CrossRefDirect Link - Sarwar, M., I.H. Attitalla and M. Abdollahi, 2011. A review on the recent advances in pharmacological studies on medicinal plants: Animal studies are done but clinical studies needs completing. Asian J. Anim. Vet. Adv., 6: 867-883.
CrossRef - Motamedi, H., E. Darabpour, M. Gholipour and S.M.S. Nejad, 2010. Antibacterial effect of ethanolic and methanolic extracts of Plantago ovata and Oliveria decumbens endemic in iran against some pathogenic bacteria. Int. J. Pharmacol., 6: 117-122.
CrossRefDirect Link - Nosanchuk, J.D. and A. Casadevall, 2006. Impact of melanin on microbial virulence and clinical resistance to antimicrobial compounds. Antimicrob. Agents Chemother, 50: 3519-3528.
Direct Link - Nosanchuk, J.D., P. Valadon, H. Feldmesser and A. Casadevall, 1999. Melanization of Cryptococcus neoformans in murine infection. Mol. Cell Biol., 19: 745-750.
Direct Link - Polak, A. and D.M. Dixon, 1989. Loss of melanin in Wangiella dermatitidis does not result in greater susceptibility to antifungal agents. Antimicrob. Agents Chemother, 33: 1639-1640.
Direct Link - Pooja, V., H. Sanwal, A. Goyal, S. Bhatnagar and A.K. Srivastava, 2012. Activity of Myristica fragrans and its effect against filamentous and non-filamentous fungus. Int. J. Pharm. Pharm. Sci., Vol. 4.
Direct Link - Saadabi, A.M.A., 2006. Antifungal activity of some saudi plants used in traditional medicine. Asian J. Plant Sci., 5: 907-909.
CrossRefDirect Link - Bonjar, G.H.S., S. Aghighi and A.K. Nik, 2004. Antibacterial and antifungal survey in plants used in indigenous herbal-medicine of South East regions of Iran. J. Biol. Sci., 4: 405-412.
CrossRefDirect Link - Young, L.Y., C.M. Hull and J. Heitman, 2003. Disruption of ergosterol biosynthesis confers resistance to amphotericin B in Candida lusitaniae. Antimicrob. Agents Chemother, 47: 2717-2724.
CrossRef