
Yosep Seran Mau
Faculty of Agriculture, University of Nusa Cendana. Jl. Adisucipto, Penfui, 85001 Kupang, East Nusa Tenggara, Indonesia
LiveDNA: 62.22036
Antonius Simon Sebho Ndiwa
Faculty of Agriculture, University of Nusa Cendana. Jl. Adisucipto, Penfui, 85001 Kupang, East Nusa Tenggara, Indonesia
LiveDNA: 62.22691
Jenny Ema Rut Markus
Faculty of Agriculture, University of Nusa Cendana. Jl. Adisucipto, Penfui, 85001 Kupang, East Nusa Tenggara, Indonesia
LiveDNA: 62.22351
Shirly Seahan Oematan
Faculty of Agriculture, University of Nusa Cendana. Jl. Adisucipto, Penfui, 85001 Kupang, East Nusa Tenggara, Indonesia
LiveDNA: 62.22904
Anggiani Nasution
Indonesian Center for Rice Research. Jl. Raya No. 9 Sukamandi, 41256 Subang, West Java, Indonesia
LiveDNA: 62.22380
Dody Dwi Handoko
Indonesian Center for Rice Research. Jl. Raya No. 9 Sukamandi, 41256 Subang, West Java, Indonesia
LiveDNA: 62.22271
Kisman Makbul
Faculty of Agriculture, University of Mataram. Jl. Majapahit No. 42, 83115 Mataram, West Nusa Tenggara, Indonesia
LiveDNA: 62.22433
Asian Journal of Crop Science
Year: 2018 | Volume: 10 | Issue: 2 | Page No.: 53-65







ABSTRACT
Background and Objective: Blast is one of the most destructive diseases of rice worldwide. Use of blast resistant varieties is considered the most effective and economical way of controlling the disease. Local cultivars of upland rice are potential sources of blast resistance, especially for development of superior red and black rice varieties. The objectives of this study were: (1) To assess blast resistance levels of red and black upland rice local cultivars against four dominant races of Pyricularia grisea (P. grisea) in Indonesia and (2) To identify red and black upland rice cultivars having resistance to both single and multiple races of P. grisea. Materials and Methods: The study was conducted in two sites, i.e., in the Glasshouse of Faculty of Agriculture, University of Nusa Cendana and the Glasshouse of Muara Research Station of Indonesian Center for Rice Research during August-November, 2015. A completely randomized design was employed with two-factor treatments, i.e., rice genotype and P. grisea race. Forty two rice genotypes (39 local cultivars and three check varieties) were inoculated with four races of P. grisea, each was five replicates. The variables observed were infection period and disease severity, each was subjected to analysis of variance, followed by DMRT post hoc test. The disease severity was used for classification of blast resistance levels of tested rice genotypes. Results: There was a highly significant interaction effect of rice genotype by P. grisea race on either infection period or disease severity. The tested rice genotypes were classified into six resistance levels for races 033 and 073 inoculations and five resistance levels for races 133 and 173 inoculations. Conclusion: Twenty three local genotypes exhibited blast resistant reaction (highly resistant+resistant), of which 11 genotypes were resistant to single race and 12 genotypes were resistant to multiple races of P. grisea.
PDF Abstract XML References Citation
Received: December 03, 2017;
Accepted: March 12, 2018;
Published: May 31, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Yosep Seran Mau, Antonius Simon Sebho Ndiwa, Jenny Ema Rut Markus, Shirly Seahan Oematan, Anggiani Nasution, Dody Dwi Handoko and Kisman Makbul, 2018. Blast Resistance Levels of Red and Black Upland Rice Local Cultivars from Indonesia. Asian Journal of Crop Science, 10: 53-65.
DOI: 10.3923/ajcs.2018.53.65
URL: https://scialert.net/abstract/?doi=ajcs.2018.53.65
DOI: 10.3923/ajcs.2018.53.65
URL: https://scialert.net/abstract/?doi=ajcs.2018.53.65
INTRODUCTION
Rice (Oryza sativa L.) is one of the most important staple food crops in the world and the most important one in Indonesia. Efforts to increase rice production to meet the increasing demand for rice are currently challenged by both biotic and abiotic stresses. One of the most challenging biotic stresses for rice production worldwide is blast disease caused by Pyricularia grisea (Cooke) Sacc. (cyn: P. oryzae Cavara)1,2, teleomorph Magnaporthe oryzae3,4. Pyricularia grisea can infect all growth stages of rice crop such as leaves, culm and panicles, thus affecting photosynthetic efficiency and grain yield5,6. Substantial yield losses had been reported to be caused by blast disease. In susceptible varieties, yield losses due to blast ranged from 20-100% in Japan and India7. In Vietnam, Hai et al.8 reported grain yield losses of 38.21-64.57% due to neck blast on susceptible rice varieties.
Rice blast disease has been so far controlled using fungicides, resistant genotypes, good agronomic practices and bio-control agents5. Among these, the use of resistant genotypes is considered the most economical, efficient and environmentally friendly method to control rice blast9-12. Control of blast disease using resistant varieties, however, is not always long lasting due to the breakdown of the resistance. This is especially true for monogenic resistance due to the nature of high variability of the pathogen and continuous generation of novel pathogenic variation resulting in the appearance of new pathogenic races in a relatively short time after cultivation of the cultivar13.
Development of new rice varieties with resistance to multiple races of the blast fungus is considered an efficient way of delaying the breakdown of the resistance. This can be done by selecting new sources of resistance or by pyramiding resistance genes for development of durable blast resistance varieties. Wild species and local cultivars are likely sources for new and broad spectrum blast resistant genes. Upland rice local cultivars, in particular, are potential sources of new blast resistance genes since these genotypes have been paid less attention for as compared to the lowland paddy rice.
Indonesia is rich in local cultivars of upland rice, most particularly the pigmented (red and purple/black) upland rice that can be used as sources of blast resistance. These pigmented rice cultivars have so far obtained less attention for development as compared to the white rice. Pigmented rice, however, has greater nutritious properties, especially its health-embracing benefits, which causes the consumer’s demand for this rice is now significantly greater than before14,15.
Pigmented rice here refers to rice genotypes having colored pericarp such as red or purple/black rice16. Pigmented rice contains high levels of polyphenolic compounds15,17 that serve health-promoting properties such as anti-cancer18, antioxidants19 and other disease-curing properties20. The phenolic compounds in rice are also important for plant development and protect the plant from diseases and infections17.
Despite its health-promoting properties, the pigmented rice (red and black rice) is currently limitedly available, due partly to the limited number of existing superior pigmented rice varieties21,22. In Indonesia, only a few improved red rice varieties are available at present, of which only one is upland rice superior variety23. Improved black rice variety is even unavailable yet at present. The increasing demands for pigmented rice necessitates development of superior red and black upland rice varieties by employing the available genetic resources, including local cultivars. A number of red and black upland rice local cultivars collected from various locations in East Nusa Tenggara (ENT) province of Indonesia have been assessed for their genetic diversity24 and this collection is very genetically diverse in both morphological and agronomical characters. These red and black upland rice germplasm, however, have not been evaluated for blast resistance, which is an important trait for superior red and black rice varietal production. New sources of broad spectrum blast resistance genes are likely to be obtained from these local germplasm owing to their high genetic diversity24. Development of superior red and black rice varieties is a demanding task as the selection for superior traits includes both desirable agronomical traits and also qualitative traits such as pericarp color. It would be of a great benefit to obtain parental source from red and black rice genotypes which also possess broad spectrum blast resistance as it will accelerate the breeding program without the need to select for pericarp color. At present, there is no report on broad spectrum blast resistance in pigmented upland rice cultivars. Studies on blast resistance in pigmented (red and purple/black) upland rice had previously been carried out in Malaysia25 but only a singe race of P. grisea was employed in the study. The QTLs for blast resistance had also been identified in rice germplasm12,7 but mostly are on white rice. Thus, the present study offers a great opportunity to obtain broad spectrum blast resistance in red and black upland rice cultivars through evaluation of resistance to multiple races of P. grisea. The objectives of this study were: (1) To assess blast resistance levels of red and black upland rice local cultivars against four dominant races of P. grisea in Indonesia and (2) To identify red and black upland rice cultivars having resistance to both single and multiple races of P. grisea.
MATERIALS AND METHODS
Rice genotypes: A total of 42 rice genotypes were included in this experiment. The genotypes consisted of 39 local cultivars of red and black upland rice and three check varieties (Indonesian released superior varieties). The upland rice local cultivars were collected from farmers in various locations in East Nusa Tenggara province, Indonesia, comprised of red rice (ADN-04, ALR-01, BLA-01, BLU-01, BLU-02, HK-06, ISN-02, MANU-04, MGR-012, PAU-01, PJ-01, PKD-01, PKP-01, PMK-01, PRA-01, SBD-04, SBD-05, TLB-02, TLB-04, WTN-01) and purple/black rice (ADN-03, ADN-05, ALR-02, BLA-01, BLU-03, CBL-01, ISN-03, KMD-01, MGP-01, MKL-01, NGR-021, PMT-01, SBD-01, SBD-02, SBD-03, SBR-01, SLR-07, TLB-05, WTN-02). The three check varieties were Kencana Bali (blast susceptible check), Asahan (blast resistant check) and Aek Sibundong (Indonesian released red rice variety), all were kindly provided by Indonesian Center for Rice Research, Sukamandi, West Java, Indonesia.
P. grisea races: Four most dominant races of P. grisea in Indonesia, i.e., races 033, 073, 133 and 173 were used in the present study. The four races of the blast fungus were obtained from Rice Pathology Laboratory of Indonesian Center for Rice Research in Muara Research Station, Bogor.
Experimental site: This experiment was carried out in two locations, i.e., the Glasshouse of Faculty of Agriculture, University of Nusa Cendana and the Glasshouse of Muara Research Station of Indonesian Center for Rice Research. Environmental conditions of the two glasshouses were set to be nearly similar to allow replicated evaluation of blast resistance. The blast resistance evaluation was carried out during August-November, 2015.
Experimental design: The present experiment was laid out in a completely randomized design with a factorial treatment design of two factors, i.e., rice genotype (42 genotypes) and P. grisea race (four races). Each experimental unit consisted of five replicates, 3 replicates in the Glasshouse of University of Nusa Cendana and 2 replicates in the Glasshouse of Muara Research Station of Indonesian Center for Rice Research.
Plant cultivation: Tested rice genotypes were sown in plastic trays of 45 cm×30 cm×10 cm filled with soil. Five seeds of each genotype were planted in each of the nine rows contained in each plastic tray. The susceptible check variety Kencana Bali was also planted in every plastic tray used in the experiment. A basal fertilizer containing 5 g urea, 1.3 g (trisodium phosphate) TSP and 1.2 g KCl was applied to every 10 kg dry soil used for growing the tested rice genotypes’ seedlings.
Preparation of conidial suspension: Each of P. grisea races was propagated on the potato dextrose agar medium in a petri dish for 7 days. The pure culture of each fungus isolate was then transferred into oat meal agar media for 12 days. Ten days after the transfer of the pure culture, the fungus colony was rubbed using distilled water plus 0.01 g L–1 streptomycin and was then stored in a luminous incubator with a 20-watt fluorescent lamp for 48 h. On the 12th day, rubbing of the fungus colony was repeated by using an image brush No.10 and sterile water containing 0.02% Tween 20 to get spore solution.
Inoculation and disease assessment: A spore concentration of 2×105 mL–1 conidia was used for inoculation of the rice plants. Counting of the conidia was performed by using a hemocytometer. Inoculation was done by spraying on 21 days old plants or 4-5 leaf stadia. The inoculated plants were incubated for 24 h in a humid chamber and then transferred to a glasshouse. High moisture condition (80-90%) in the glasshouse was maintained by using an in-house condensation.
Variables observed in this study included: (1) Infection period and (2) Disease severity (disease score). Infection period was observed from day 1 after inoculation while disease severity was observed at 7 days after inoculation (DAI). The disease severity was recorded based on visual estimation using a 0-9 scale of SES IRRI26 method, where, 0 = No lesions observed, 1 = Small brown specks of pin-point size or larger brown specks without sporulating, 2 = Small roundish to slightly elongated necrotic gray spots, about 1-2 mm in diameter, with a distinct brown margin, lesions are mostly found on lower leaves, 3 = Lesion type is the same as in scale 2, but significant number of lesions are on the upper leaves, 4 = Typical susceptible blast lesions 3 mm or longer, infecting less than 4% of the leaf area, 5 = Typical blast lesions infecting 4-10% of the leaf area, 6 = Typical blast lesions infecting 11-25% of the leaf area, 7 = Typical blast lesions infecting 26-50% of the leaf area, 8 = Typical blast lesions infecting 51-75% of the leaf area and many leaves are dead and 9 = More than 75% leaf area affected. Plants with blast scores of 0-1 were classified ‘highly resistant,' 2 were classified ‘resistant,' 3 were classified ‘moderately resistant", 4 were classified ‘moderately susceptible’, 5-7 were classified ‘susceptible’ and 8-9 were classified ‘highly susceptible’.
Statistical analysis: Pooled data of infection period and disease severity from the two experiments was each subjected to a two-way analysis of variance to determine the effect of treatments on the observed variables. Analysis of variance was performed using Genstat Version 1227. A post hoc test of Duncan’s multiple range test (DMRT) was performed to separate the treatment means (p<0.05). Mean disease severity was used to classify the blast resistance levels of tested rice genotypes.
RESULTS
Effect of treatments on infection period and disease severity: Results of analysis of variance revealed a highly significant effect (p<0.01) of interaction between rice genotype and P. grisea race for blast disease traits: Infection period and disease severity (Table 1). The highly significant interaction effect of rice genotype by P. grisea race on these two variables indicates different responses of the tested rice genotypes against P. grisea races and the responses varied considerably with the different treatment of the fungus races.
As with the interaction effect, each of the single factor treatments of rice genotype and P. grisea race also highly significantly affected (p<0.01) the blast infection period and disease severity indicating substantial differences in the two blast disease traits among the rice genotypes and P. grisea races.
Infection period: The study results revealed that infection period varied considerably among tested rice genotypes and this variation occurred over the four pathogenic races used for inoculation. Two-way table of blast disease infection period of rice genotypes inoculated with four aces of P. grisea is presented in Table 2. The infection period of blast disease within the tested rice genotypes ranged between 2.0 and 7.0 DAI was shown in Table 2.
The shortest/fasted infection period did occur in 2 days after inoculation, which was observed in the local genotypes SBD-02 and BLU-01 inoculated with race 033 (2.0 and 2.2 DAI, respectively), ALR-02, HK-06, SBD-01, SBD-03 and SBD-04 inoculated with race 073 (2.0 DAI), TLB-02 inoculated with race 133 (2.0 DAI) and ISN-03, Manu-04 and SBD-03 inoculated with race 173 (2.0 DAI). The local genotype SBD-03 showed the shortest infection period for three race inoculations, i.e., races 033, 073 and 173.
The longest infection period was observed at 7 DAI in SLR-07 and TLB-04 for inoculation with race 033, MGR-02 for inoculation with race 073, BLA-01, HK-06, SBD-01, SBD-03, WTN-02 for inoculation with Race 133 and ALR-02, PKD-01, SBD-01, SBD-02 for inoculation with race 173. Differences in rice genotype and P. grisea race pairwise combinations for the longest infection period probably indicates differences in host-pathogen gene combinations controlling this observed trait.
Over the four race inoculations, the longest infection period was observed in the blast-resistant check variety Asahan (5.20 DAI), but it was not significantly different from that of local genotypes BLA-01, PAU-01 and TLB-04. Meanwhile, the shortest infection period was observed in ISN-03, TLB-02 and PJ-01(2.0 DAI for the first two genotypes and 3.0 DAI for the last genotype). On the other hand, over all tested rice genotypes, race 133 caused the longest infection period on average (4.47 DAI) while race 073 caused the shortest infection period (3.29 DAI), which was not statistically different from that of race 033 (3.34 DAI).
Disease severity: Researchers observed highly significant variation (p<0.01) of blast disease severity among tested rice genotypes and so did the effect of single factor rice genotype. Meanwhile, the single factor P. grisea race significantly affected (p<0.05) the trait. Overall, the observed disease severity (disease score) ranged between 1.0 and 8.2 (disease score of 0-9) (Table 3). The significant interaction effect indicates that the blast disease severity differed significantly among the tested rice genotypes and the degree of these differences did change with the change of P. grisea race.
| Table 1: | Analysis of variance of infection period and disease severity of 42 rice genotypes inoculated with four races of Pyricularia grisea |
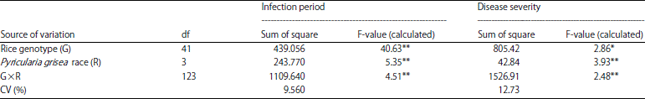 | |
df: Degree of freedom, **Highly significant (p<0.01), *Significant (p<0.05) | |
| Table 2: | Mean infection period (DAI) of 42 rice genotypes inoculated with four races of Pyricularia grisea |
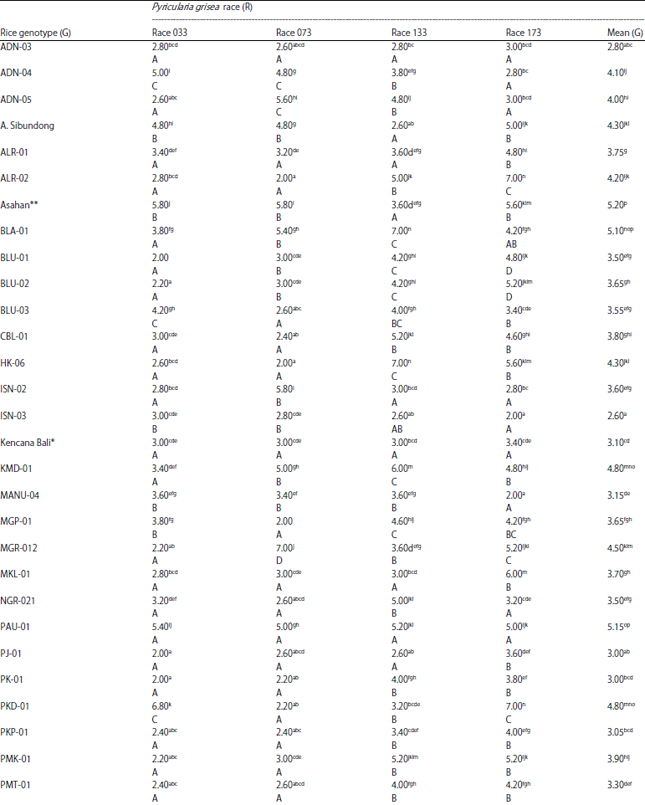  | |
Data is the average of five replicates. Values in the same column and row with the same letter(s) are not significantly different at 5% DMRT. Small letters indicate comparison within the same column while capital letters indicate comparison within the same row. *Susceptible check, **Resistant check | |
For all treatment combinations shown in Table 3, the highest disease mean score was 8.2 observed in SBD-02 when inoculated with race 033 while the lowest disease score (1.0) was observed in several rice genotypes, i.e., SLR-07 (race 033), SBD-05 (race 133) and SBD-02 (race 173). It was interesting to note from Table 3 that the local genotype SBD-02 suffered the most when inoculated with race 033, but it suffered the least when inoculated with race 173. The high disease score suffered by SBD-02 when inoculated with race 033 agrees well with its reaction for infection period, where this local genotype also produced the shortest infection period (2 DAI) when inoculated with the same race.
The highest mean disease score over all four race inoculations was observed in the blast susceptible check variety Kencana Bali (6.2) while the shortest disease score (1.0) was observed in the blast-resistant check variety Asahan and the local genotype SLR-07. The last two genotypes also showed the lowest disease score in race 033 inoculation and relatively lower disease scores than most of the tested rice genotypes in the other three race inoculations. Data in Table 3 shows that over the four race inoculations, all local rice genotypes produced lower disease score than the susceptible check Kencana Bali but these genotypes, except SLR-07, produced higher disease scores than the resistant check Asahan. The SLR-07 and Asahan showed lower mean disease scores over all four race inoculations, presumably, indicating their overall resistance to P. grisea races employed in this study.
Race 033 caused the highest disease score (4.05) over all tested rice genotypes, which then followed by race 073 that produced the second highest disease score (3.81) while races 073 and 133 caused statistically similar disease scores (3.57 and 3.46, respectively). In race 033 inoculation, only two local genotypes that produced similar or higher disease scores than the susceptible check Kencana Bali while the rest of the genotypes produced lower disease scores. Disease scores of these genotypes, except SLR-07, were higher than that of the resistant check variety Asahan.
In race 073 inoculation, five local genotypes produced similar or higher disease scores than the susceptible check Kencana Bali and the other local genotypes produced lower disease score than Kencana Bali. Except for ADN-03, ALR-01, ISN-02 and SBD-02, disease scores of local genotypes were higher than the resistant check Asahan.
| Table 3: | Mean blast disease severity (disease score) of 42 rice genotypes inoculated with four races of Pyricularia grisea |
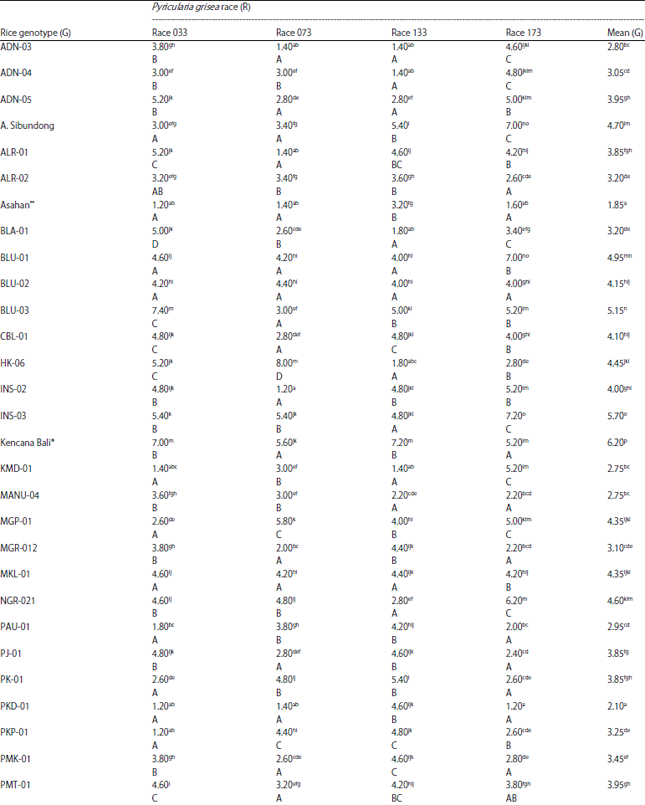 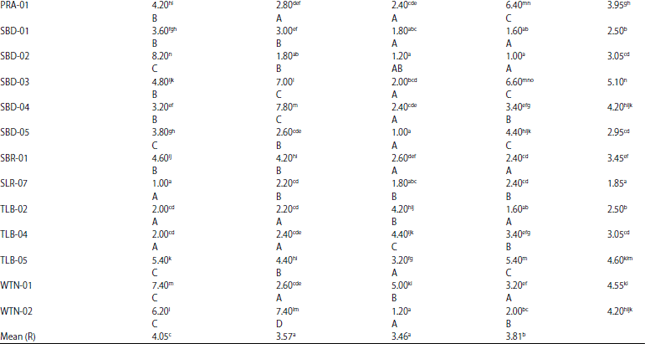 | |
Data is the average of five replicates. Values in the same column and row with the same letter(s) are not significantly different at 5% DMRT. Small letters indicate comparison within the same column while capital letters indicate comparison within the same row. *Susceptible check, **Resistant check. | |
All local genotypes exhibited lower disease scores than the susceptible check Kencana Bali when the genotypes were inoculated with race 133. Of these local genotypes, ten genotypes (SBD-05, WTN-02, SLR-07, SBD-02, SBD-01, KMD-01, HK-06, BLA-01, ADN-4 and ADN-03) produced significantly lower disease scores than the resistant check variety Asahan. Furthermore, in race 173 inoculation, five local genotypes produced disease score similar to that of the resistant check Asahan while ten local genotypes produced higher disease scores than the susceptible check Kencana Bali.
Classification of blast resistance level: Blast resistance levels of tested rice genotypes were assessed based on the blast disease score (Table 3). The blast resistance levels of tested rice genotypes were presented in Table 4.
It is clearly seen in Table 4 that blast resistance levels of tested rice genotypes segregated substantially across the four races employed. The 42 rice genotypes were classified into six resistance categories (highly resistant, resistant, moderately resistant, moderately susceptible, susceptible and highly susceptible) for race 033 and 073 inoculations and five resistance categories (highly resistant, resistant, moderately resistant, moderately susceptible, susceptible) for race 133 and 173 inoculations.
In race 033 inoculation, five rice genotypes were highly resistant and three genotypes were resistant while the rest of the genotypes fell into moderately resistant (six genotypes), moderately susceptible (eight genotypes), susceptible (19 genotypes) and highly susceptible (one genotype). When inoculated with race 073, the tested rice genotypes segregated into five highly resistant, five resistant, 16 moderate resistant, seven moderately susceptible, seven susceptible and two highly susceptible genotypes. A higher number of highly resistant (six) and resistant (eight) rice genotypes were observed when inoculation was done using race 133. In this inoculation, the number of rice genotypes showing moderately resistant, moderately susceptible and susceptible reaction was, respectively, 5, 10 and 13 genotypes. No highly susceptible resistance genotype was observed in race 133 inoculation. Inoculation with race 173 resulted in only two genotypes being classified highly resistant while ten genotypes were classified resistant, nine genotypes were moderately resistant, 6 genotypes were moderately susceptible and 15 genotypes were susceptible. As with race 133 inoculation, no highly susceptible genotype was observed in race 173 inoculation.
| Table 4: | Mean disease scores and blast resistance levels of 42 rice genotypes inoculated with four races of Pyricularia grisea |
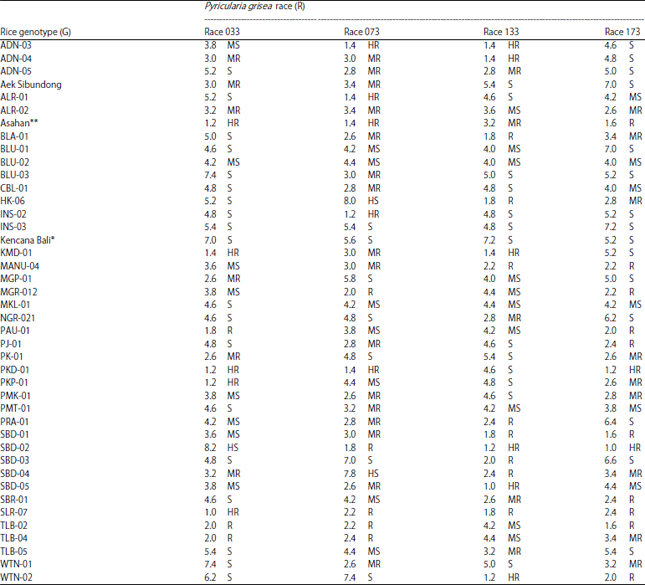 | |
HR: Highly resistant, R: Resistant, MR: Moderately resistant, MS: Moderately susceptible, S: Susceptibe, HS: Highly susceptible. *Susceptible check, **Resistant check | |
Results of blast resistance classification (Table 4) demonstrate that number of highly resistant, resistant, moderately resistant, moderately susceptible, susceptible and highly susceptible genotypes varied considerably according to the pathogenic races of P. grisea used for inoculation. This implied that different host resistance/susceptibility genes and pathogen virulence/avirulence genes might have involved in the host-pathogen interaction, which resulted in different blast resistance phenotypes observed in the present study.
Rice genotypes exhibiting resistance to single and multiple races of P. grisea: The present study results revealed that a total of 24 rice genotypes (23 local cultivars and one check variety) showed blast resistance (highly resistant+resistant) to either single race or multiple races of P. grisea. Of these genotypes, 11 genotypes conferred resistance to a single race while 13 genotypes conferred resistance to multiple races of the blast fungus (Table 5). The single race resistant genotype included one genotype (PKP-01) that was resistant to race 033, two genotypes (ALR-01 and ISN-02) that were, respectively, highly resistant and resistant to race 073, one genotype that was highly resistant and six genotypes that were resistant to race 133 and two genotypes (PJ-01 and SBR-01) that were resistant race 173.
| Table 5: | Rice genotypes showing resistance to single and multiple races of Pyricularia grisea |
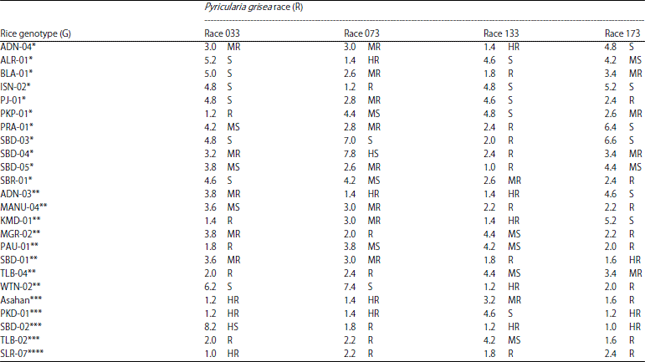 | |
HR: Highly resistant, R: Resistant, MR: Moderately resistant, MS: Moderately susceptible, S: Susceptibe, HS: Highly Susceptible. *Highly resistant/resistant to one race, **highly resistant/resistant to two races, ***Highly resistant/resistant to three races, ****Highly resistant/resistant to four races of P. grisea | |
The rice genotypes showing resistant to multiple races of P. grisea included eight rice genotypes that were resistant to two races, four genotypes that were resistant to three races and only one rice genotype that was resistant to all the four races employed in this study. The eight rice genotypes showing resistance (highly resistant and resistant) to two races included ADN-03 (races 073 and 133), KMD-01 (races 033 and 133), Manu-04 (races 133 and 173), MGR-02 (races 073 and 173), PAU-01 (races 033 and 173), SBD-01 (races 133 and 173), TLB-04 (races 033 and 073) and WTN-02 (races 133 and 173). Rice genotypes showing resistance to three races of P. oryza included the blast-resistant check variety Asahan and local cultivars PKD-01 (races 033, 073 and 173), SBD-02 (races 073, 133 and 173) and TLB-02 (races 033, 073 and 173). The only rice genotype showing resistance to all four races was the local cultivar SLR-07.
DISCUSSION
Present research results demonstrated significant variation in both blast infection period and disease severity of tested rice genotypes across the four races of P. grisea inoculations indicating that interaction of different host resistance - pathogen virulence genes might have taken place. Rice genotypes showing shorter infection period might demonstrate their compatible interaction with the pathogen for the onset of disease to take place faster while those with longer infection period might demonstrate their resistance toward the pathogen races in form of the delay of disease onset. Rice genotypes showing shorter infection period were expected to suffer higher disease severity as compared to those with longer infection period since the magnitude of disease severity is highly dependent on the length of time of compatible host-pathogen interaction under favorable environmental conditions28. As expected, present study analysis showed that most of the rice genotypes having shorter infection period tended to have higher disease severity at the time of disease assessment (7 DAI). However, we found a few of tested rice genotypes that deviated from this common expectation. These few rice genotypes exhibited shorter infection period (<3 DAI) but, on the contrary, showed lower disease severity at 7 DAI. This was observed in rice genotypes ADN-03, SLR-07, Aek Sibundong, PJ-01, PKP-01, SBD-02, TLB-02 and WTN-02, each was corresponding to a single to two races of P. oryzae.
These analysis of correlation between infection period and disease score revealed a moderate correlation coefficient of -0.53 for race 033, -0.45 for race 073, -0.64 for race 133 and -0.33 for race 173 inoculations. These results indicate that blast disease severity tended to show a negative correlation with infection period. The blast disease responses of rice genotypes that deviated from the observed negative correlation, presumably, show part of their resistance mechanism toward blast infection. Shorter disease infection period followed by a lower disease score may explain their resistance mechanisms against the pathogen upon the infection through induction of hypersensitive reaction (HR), which occurs via gene-for-gene interaction29. This hypersensitive response might have slowed down the progress of the disease, which was recorded as lower disease score at the time of disease assessment carried out at the end of the experiment (7 DAI).
Results showed that more than 50% of tested rice genotypes showed resistance (highly resistant and resistant reactions) to either single race or multiple races of P. grisea. Of these, 13 genotypes showed resistance to multiple races (two, three and four races) of the pathogen. This number is much higher than the reported blast resistance in upland rice in Indonesia30, where all six upland rice genotypes tested showed no ‘resistant reaction’ to the blast fungus field isolate. Blast resistance in pigmented upland rice had previously been reported from Malaysia25 but the resistance was only to a singe race of P. grisea. Resistance to multiple races of the blast fungus in the common white rice genotypes/germplasm had been reported by Wang et al.12, Fukuoka et al.13, Sharma et al.7, Haggag and Tawfik31 and Chaipanya et al.32 but none has been reported in red and black upland rice.
Red and black upland rice genotypes showing resistance to multiple races of P. grisea as we observed in the current study are considered having a broad-spectrum resistance. Broad spectrum resistance is important in breeding for durable blast resistance. Broad-spectrum resistance can ensure the durability of resistance and durable resistance can be obtained through strategies such as planting mixture varieties, pyramiding of major R genes and using partial resistance genes33.
It is not known how many resistance genes are controlling blast resistance in the red and black local upland rice genotypes showing resistance to single or multiple races of P. grisea employed in this study. However, a previous study employing the wild rice O. rufipogon and rice cultivar IR64 (white rice) by Utami et al.34 found that resistance to race 033 was controlled by QTL Pir9-2(t) and QTL Pir12-2(t) in duplicate epistatic interaction while resistance to race 173 was controlled by a race specific-complementary additive QTL Pir2-3(t). These findings imply that resistance to race 033 and race 173 was controlled by at least three genes. Furthermore, genes controlling resistance to other two races that were also employed in this study, i.e., 073 and 133 are still unknown at present. Thus, red and black local upland rice genotypes employed in this study that showed resistance to three and four races of P. grisea are most likely to harbor more than three blast resistance genes/loci. Many QTLs for blast resistance have been reported by Wang et al.12 and Sharma et al.7 but mostly are on the common white rice.
Of the 11 genotypes that showed resistance to two-four races of P. grisea, six of which are red rice (MGR-012, PAU-01, TLB-02, TLB-04, WTN-02, PKD-01 and TLB-02) while the rest five genotypes are purple/black rice (ADN-03, KMD-01, SBD-01, SBD-02 and SLR-07)24. These genotypes are invaluable genetic resources that can be employed for development of blast resistance in red and black upland rice varieties. The genotype SLR-07 in particular, which is resistant to four races of P. grisea, is a very invaluable genetic resource for the development of black rice variety with broad-spectrum and durable blast resistance. No such superior black rice variety has been released in Indonesia at present. Development of superior red and black upland rice varieties utilizing broad-spectrum blast resistant genotypes identified in this study will accelerate achievement of the breeding objectives as there will be no need for selection of pericarp color and only a few genotypes can be involved in pyramiding of the blast resistance genes thus saving a significant amount of time, resources and breeding efforts.
CONCLUSION
The study results showed that the red and black upland rice genotypes evaluated were classified into six resistance levels (highly resistant, resistant, moderately resistant, moderately susceptible, susceptible and highly susceptible) for races 033 and 073 inoculations and five resistance levels (highly resistant, resistant, moderately resistant, moderately susceptible, susceptible) for races 133 and 173 inoculations. Of 39 local cultivars of red and black upland rice tested, 23 exhibited blast resistant reaction (highly resistant+resistant), 11 of which were resistant to single race and the rest 12 were resistant to multiple races of P. grisea. Eight local rice genotypes (ADN-03, MANU-04, KMD-01, MGR-02, PAU-01, SBD-01, TLB-04 and WTN-02) showed blast resistant reaction to two races, four genotypes (PKD-01, SBD-02 and TLB-02) showed resistant reaction to three races and one genotype (SLR-07) showed resistant reaction to all four races of P. grisea.
SIGNIFICANCE STATEMENT
This study revealed resistance to multiple races of P. grisea in red and black upland rice local cultivars that can be useful in development of red and black superior rice varieties with broad spectrum resistance to blast disease. This study results will help researchers, most particularly plant breeders and plant pathologist to uncover the critical area of development of superior broad spectrum blast resistant red and black upland rice varieties that are unavailable yet at present.
ACKNOWLEDGMENT
This study was carried out with co-funding supports from Directorate of Research and Community Services (DRPM) of The Indonesian Ministry of Research, Technology and Higher Education through Fundamental Research scheme 2015 (Grant Number 85/UN15.19.1/LT/2015) and Indonesian Agency for Agriculture Research and Development (IAARD) of The Indonesian Ministry of Agriculture through KKP3N Research Scheme 2015 (Grant Number 44.38/HM.240/I.1/3/2015.K). We gratefully thank DRPM and IAARD for providing us with the funding supports. We also thank Aro Balol for the great technical assistance carried out in the Glasshouse.
REFERENCES
- Rossman, A.Y., R.J. Howard and B. Valent, 1990. Pyricularia grisea, the correct name for the rice blast disease fungus. Mycologia, 82: 509-512.
CrossRefDirect Link - Couch, B.C. and L.M. Kohn, 2002. A multilocus gene genealogy concordant with host preference indicates segregation of a new species, Magnaporthe oryzae, from M. grisea. Mycologia, 94: 683-693.
CrossRefDirect Link - Zhang, N., S. Zhao and Q. Shen, 2011. A six-gene phylogeny reveals the evolution of mode of infection in the rice blast fungus and allied species. Mycologia, 103: 1267-1276.
CrossRefDirect Link - Choi, J., S.Y. Park, B.R. Kim, J.H. Roh, I.S. Oh, S.S. Han and Y.H. Lee, 2013. Comparative analysis of pathogenicity and phylogenetic relationship in Magnaporthe grisea species complex. PLoS ONE, Vol. 8.
CrossRefDirect Link - Ribot, C., J. Hirsch, S. Balzergue, D. Tharreau, J.L. Notteghem, M.H. Lebrun and J.B. Morel, 2008. Susceptibility of rice to the blast fungus, Magnaporthe grisea. J. Plant Physiol., 165: 114-124.
CrossRefDirect Link - Vasudevan, K., C.M.V. Cruz, W. Gruissem and N.K. Bhullar, 2014. Large scale germplasm screening for identification of novel rice blast resistance sources. Front. Plant Sci., Vol. 5.
CrossRefDirect Link - Sharma, T.R., A.K. Rai, S.K. Gupta, J. Vijayan, B.N. Devanna and S. Ray, 2012. Rice blast management through host-plant resistance: Retrospect and prospects. Agric. Res., 1: 37-52.
CrossRefDirect Link - Hai, L.H., P.V. Kim, P.V. Du, T.T.T. Thuy and D.N.Thanh, 2007. Grain yield and grain-milling quality as affected by rice blast disease (Pyricularia grisea), at my Thanh Nam, Cai Lay, Tien Giang. Omonrice, 15: 102-107.
Direct Link - Ghazanfar, M.U., A. Habib and S.T. Sahi, 2009. Screening of rice germplasm against Pyricularia oryzae the cause of rice blast disease. Pak. J. Phytopathol., 21: 41-44.
Direct Link - Idowu, O.O., A.O. Salami, S.A. Ajayi, R.O. Akinwale and Y. Sere, 2013. Varietal resistance of rice to blast fungus Magnaporthe oryzae at two sites in Southwestern Nigeria. Afr. J. Biotechnol., 12: 5173-5182.
Direct Link - Wang, C., Y. Yang, X. Yuan, Q. Xu, Y. Feng, H. Yu and Y. Wang, 2014. Genome-wide association study of blast resistance in indica rice. BMC Plant Biol., Vol. 14.
CrossRefDirect Link - Fukuoka, S., N. Saka, H. Koga, K. Ono and T. Shimizu et al., 2009. Loss of function of a proline-containing protein confers durable disease resistance in rice. Science, 325: 998-1001.
CrossRefDirect Link - Ahmad, F., M.M. Hanafi, M.A. Hakim, M.Y. Rafii, I.W. Arolu and S.N.A. Abdullah, 2015. Genetic divergence and heritability of 42 coloured upland rice genotypes (Oryza sativa) as revealed by microsatellites marker and agro-morphological traits. PLoS ONE, Vol. 10.
CrossRefDirect Link - Rerkasem, B., S. Jumrus, N. Yimyam and C. Prom-u-Thai, 2015. Variation of grain nutritional quality among Thai purple rice genotypes grown at two different altitudes. ScienceAsia, 41: 377-385.
CrossRefDirect Link - Abd Latiff, N., S.A.Z. Alam, S.Z. Hanapi, N. Supari, M.A. Javed, L.C. Tin and M.R. Sarmidi, 2017. Evaluation of antioxidant activity and total polyphenols content on upland rice. J. Nat. Prod. Plant Resour., 7: 1-6.
Direct Link - Shao, Y., G. Zhang and J. Bao, 2011. Total phenolic content and antioxidant capacity of rice grains with extremely small size. Afr. J. Agric. Res., 6: 2289-2293.
Direct Link - Huang, Y.P. and H.M. Lai, 2016. Bioactive compounds and antioxidative activity of colored rice bran. J. Food Drug Anal., 24: 564-574.
CrossRefDirect Link - Deng, G.F., X.R. Xu, Y. Zhang, D. Li, R.Y. Gan and H.B. Li, 2013. Phenolic compounds and bioactivities of pigmented rice. Crit. Rev. Food Sci. Nutr., 53: 296-306.
CrossRefDirect Link - Hermanto, 2012. [Newly released superior rice varieties]. Warta Penelitian Pengembangan Pertanian, 34: 8-9, (In Indonesian).
Direct Link - Mau, Y.S., J.E. Markus, Shirly, S. Oematan and A. Ndiwa et al., 2017. Genetic diversity of red and black upland rice accessions from East Nusa Tenggara, Indonesia as revealed by agro-morphological characters. Biodiversitas, 18: 197-211.
CrossRefDirect Link - Tuhina-Khatun, M., M.M. Hanafi, M.Y. Wong and M.Y. Rafii, 2015. Reactions and diversity analysis of upland rice genotypes against blast disease of rice (Oryza sativa L.). Australas. Plant Pathol., 44: 405-412.
CrossRefDirect Link - IRRI., 2014. Standard Evaluation System for Rice. 5th Edn., International Rice Research Institute (IRRI), Los Banos, Philippines, ISBN-13: 9789712203046, Pages: 57.
Direct Link - Gururani, M.A., J. Venkatesh, C.P. Upadhyaya, A. Nookaraju, S.K. Pandey and S.W. Park, 2012. Plant disease resistance genes: Current status and future directions. Physiol. Mol. Plant Pathol., 78: 51-65.
CrossRefDirect Link - Taufik, M., T. Wijayanto and A. Wahab, 2012. The characterization and evaluation of local upland rice cultivars to blast disease (Pyricularia oryzae) resistance in South Sulawesi. Proceedings of the International Conferenceon on Sustainable Agriculture and Food Security: Challenges and Opportunities, September 27-28, 2011, Bandung, Indonesia, pp: 204-211.
- Haggag, W.M. and M.M. Tawfik, 2014. Identification of some rice genotypes resistant to blast disease in Egypt. Br. Biotechnol. J., 4: 894-903.
CrossRefDirect Link - Chaipanya, C., M.J. Telebanco-Yanoria, B. Quime, A. Longya and S. Korinsak et al., 2017. Dissection of broad-spectrum resistance of the Thai rice variety Jao Hom Nin conferred by two resistance genes against rice blast. Rice, Vol. 10.
CrossRefDirect Link - Utami, D.W., S. Moeljopawiro, H. Aswidinnoor, A. Setiawan and I. Hanarida, 2008. Blast resistance genes in wild rice Oryza rufipogon and rice cultivar IR64. Indonesian J. Agric., 1: 71-76.
Direct Link