
Suaib Suaib
Department of Plant Breeding, Faculty of Agriculture, Halu Oleo University, Kendari, Southeast Sulawesi, Indonesia
LiveDNA: 62.23167
Darwis Suleman
Department of Soil Sciences, Faculty of Agriculture, Halu Oleo University, Kendari, Southeast Sulawesi, Indonesia
Asian Journal of Crop Science
Year: 2018 | Volume: 10 | Issue: 4 | Page No.: 207-213







ABSTRACT
Background and Objectives: Higher production of maize (Zea mays L.) could be obtained by breeding in a much faster method by using the xenia and metaxenia phenomena as the main observations. The objectives of this research were to study the possibility of maize hybrid varieties seeds as crossing parents in exploiting both xenia and metaxenia phenomena in creating the base population materials for breeding the new promising candidate of higher production variety of maize. Materials and Methods: Three seeds from hybrid varieties as crossing parents were Bonanza and Ganebo varieties (sweet maize) and Arumba variety (waxy maize). These varieties were separated and sown into the 500 cm long and 300 cm wide plot, 75 cm between rows and 30 cm within rows, with one seed per hill, so that there would be 56 individual plant per plot. The authors employed diallel crossing method and obtained three of each of regular F1 kernels, reciprocal F1 kernels and open pollinated F1 kernels of female parents as data sources. F1’s kernels colour, 100 kernels weights (g) and the number of kernels per ear were the xenia components and diameter (mm) and lengths of ears (cm) were the metaxenia components. One way analysis of variance according to completely randomized design was used to analyze the observed variables and the Tukey t-test was subjected in judging the difference between F1’s. Results: The research results showed that the 100 kernels weight, the number of kernels per ear and the diameter and the lengths of ears of some F1’s as xenia and metaxenia phenomena, respectively, resulted better than their maternal parents under open pollination. Conclusion: There was a possibility to use these both hybrid varieties and xenia and metaxenia phenomena in producing the seeds materials in a faster way as base population for the next selection step in creating a promising new higher production of maize variety.
PDF Abstract XML References Citation
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Suaib Suaib and Darwis Suleman, 2018. Fastest Base Population Formation of Maize Through Exploitation of Xenia and Metaxenia Phenomena Derived from Hybrid Maize Varieties as Parents. Asian Journal of Crop Science, 10: 207-213.
DOI: 10.3923/ajcs.2018.207.213
URL: https://scialert.net/abstract/?doi=ajcs.2018.207.213
DOI: 10.3923/ajcs.2018.207.213
URL: https://scialert.net/abstract/?doi=ajcs.2018.207.213
INTRODUCTION
The term “Xenia” has already been referred to many species in 1881 by Focke1 and “metaxenia” in corn by Kellerman and Swingle2 and Denney3. These terms were two of the most widely discussed topics among the scientists during the period of 1881-1900’s. Both phenomena were defined in disparities by some scientists in that the understanding in this aspect remain unclear, conflicted and overlapping from one to another. The term of “xenia” refers to the immediate effect of pollen on endosperm as a result of double fertilization (union of sperm with egg and two polar nuclei) in seed parent, while “metaxenia” refers to the direct effect of pollen on parts of seed and fruit outside embyosac4. According to Denney3, the term of xenia was defined as the phenomenon of direct differential effects produced by pollen from different sources on the discernible characteristics, especially of size, colour, shape, chemical composition and developmental timing of part of seeds and other fruits, including the embryo, endosperm and maternal tissues, in the period from fertilization until germination. Whereas, the term of metaxenia was defined as the effect of pollen on fruit shape and other characteristics5.
From the explanations above the xenia phenomenon could be simplified as some characteristics that related to the seeds and the metaxenia may be addressed as some fruit characteristics, so that the kernel and ear characteristics of maize plant are strongly related to the xenia and metaxenia, respectively. Some attributes of kernel and ear characteristicare determining the production of maize other than fibre production. The heaviest the kernel and the longest the ear of some maize cultivars, the higher production would be obtained. Another maize kernel and ear characteristics are also determining the maize production, for example, the number of kernels per ear and the diameter of ear. Chemical composition of maize kernel would also be a xenia phenomenon like.
Following Kellerman dan Swingle2, observation about xenia and metaxenia in maize plant had been done in much more intensive, either in frequencies or in researcher team. Some of these were Kiesselbach and others1,5-13. As a result, an existing in plant breeding application in form of “top-cross” procedure has been applied in this plant species14-16. Xenia and metaxenia studies in another plant species were date palm by Swingle and others17-19, Vicia faba (L.) by Duc et al.20, Mandarin by Weingatner et al.21-23, Hylocereus polyrhizus by Mizrahi et al.24, Selenicereus mealanthus by Dag and Mizrahi25, Gossypium hirsutum by Pahlavani and Abolhasani26 and tomato plant (Solanum lycopersicum L.) by Piotto et al.27.
Possessing the higher production of maize, more precise and faster method of its breeding have already been proposed and implemented. Forming inbreed line through selfing method for several generations, generally 7-8 generations selfing was the usual way28. The higher production of crossing pairing parents was also the most parents candidate pre-requisite29. Meanwhile, the plant hybrid vigour or heterosis achieved in their filials must be in higher level eventhough this was not the exact event. It could be in the mid parent heterosis or in the high parent heterosis or in the heterobeltiosis grades30,31. To this days, no reports had successfully been published on a more precise and faster way derived from hybrid variety as crossing pairing parents.
Exploration of both xenia and metaxenia phenomena in forming the base population of a candidate of superior maize variety as selection material was rarely published, except the scientific studies under genetical aspect of some crop plants. One of the usefulness result from similar study was the existence and the application of “top cross” method, while the materials used was from the inbreed line as the crossing pairing parents14-16. Eventhough this was a preliminary study in using a hybrid varieties as crossing parents, the speculation of their fruitfullness should be proved. Faster method to determine the promising basic population for selection materials was the primary reason in observing the xenia and metaxenia by crossing of parentals pairing as hybrid variety. From this aspect, the colour of kernel, 100 kernel weight (g) and the number of kernels as xenia component were choosen, whereas the diameter (mm) and the shoot length (cm) as metaxenia element were picked-out. All variables mentioned above could be considered in determining the maize production level in as early as of the time.
This article will reported and deeply discussed in relations to the application of both xenia and metaxenia phenomena in creating a new promising superior variety of maize in a short period of time. Exploitation of both phenomena as a way to form of basic population for material selection further was also discussed.
MATERIALS AND METHODS
Location and plant materials: The field experiment was conducted at Faculty Agriculture Experimental Station II, Halu Oleo Campus, Anduonohu Kendari, Southeast Sulawesi Indonesia, from March-July, 2018. A set of tools for land preparation, plant management and harvesting and variable measurements were used, while the seeds as a source of parental crossings of the three hybrid varieties, namely Bonanza, Ganebo and Arumba were used. The two first varieties were sweet maize and dent type kernels and the latter was waxy maize type with flint type kernels. Days to harvest of these varieties were varied according to the variety but they were 60-70 days based on variety identity.
Field experimental procedure and design: Parental crossing sources were separately sown each in the 500 cm length and 300 cm wide plot and 200 cm between plots. The length of 75 cm between rows and 30 cm within row were applied as plants space and one seed per hill, so that there were seven rows for the length side and eight holes for the wide side and a total of 56 plants per plot. To avoid natural crossing pollination with different varieties the planting time arrangement was applied as an isolation method and each variety was sown two times, four rows at the first time and the remaining four rows after 2 weeks. The plants were managed according to the standard of maize hybrid plantation. Crossing parent pairings was done by artificial hands pollination to get both regular and reciprocal kernel and ear of F1 and by natural open pollination within the variety to get the F1 kernel and ear of female parent as comparison data. An artificial hands crossing was conducted when the shoot of female parent were silking and bagged by shootbag and the pollen source of male parent was bagged by tasselbag as soon before anthesis. The diallel method of Griffing32 was used in parents crossing pattern.
Research variables: Those kernels and ears were harvested when the shoots achieved the physiological mature state in which the shootleaf was browning in colour. The kernel colour, 100 kernel weight (g) and the number of kernel per ear were xenia variables, whereas the ear diameter (mm) and ear length (cm) were metaxenia variables. Three times of each variable were subjected as replication.
Data analysis: The colour of kernels as a qualitative measurements was analyzed by qualitative method then qualitatively described and the quantitative measurement was analyzed by one way analysis of variance based on a completely randomized design (CRD). Mean separation between F1’s was judged through LSD (Least Significant Difference) test of Tukey33 at alpha 0.05 of significant level. All data analysis procedures was conducted by statistical package of SAS for Windows version 6.1334.
RESULTS
By using complete dialelle crossing scheme of the three parents, those three F1 kernels and ears, three F1 reciprocals kernels and ears and three F1 open pollinated kernels and ears families were obtained. All these families were used as data analyzed in studying the xenia and metaxenia phenomena.
Xenia observation: Both Bonanza and Paragon parents were having the yellow kernel colour expression and the Arumba parent was white kernel colour. The kernel colour of qualitative character as xenia component which immediately expressed in female parent plant (F1 kernel) was observed as a result of kernel colour inheritance from male parent plant. Results of the research indicated that the yellow kernel colour of the male parent crossing with the yellow kernel colour of female parent generated the only yellow of kernel colour of F1. Meanwhile, when the male and female parents had different kernel colour like white-yellow or yellow-white, respectively, the kernel colour of female parent plant ultimately expressed with both white and yellow colour kernels (Table 1).
The 100 kernels weight (g) and the number of kernels per ear of F1’s resulted from crossing between parents were generally expressed the similar ear performances of female parent obtained to open pollination. One way analysis of variance of the effect of parental pollen on100 kernel weight and number of kernels per ear of male parents showed the highly significance of both xenia component characters. Mean squared between kernels of male and female parents and their F1’s 100 kernels weight and the number of kernel per ear were significantly differed. Ganebo and Arumba varieties as male parents and Bonanza variety as female parent were significantly differed from 100 kernel weight (Table 2).
| Table 1: | Male parent effect on F1 kernel colour of female parent |
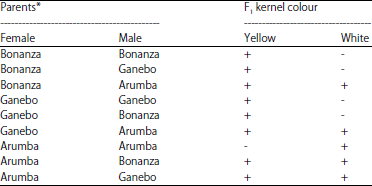 | |
| *Note of kernel colour, Bonanza: Yellow, Ganebo: Yellow and Arumba: White | |
| -: Not exist, +: Exist | |
| Table 2: | Variance analysis of the effect of m ale parent on the 100 kernel weight and the number of kernel per ear of Bonanza, Ganebo and Arumba female parents |
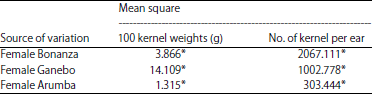 | |
| *: Statistically significant differences at the 0.05 probability level | |
| Table 3: | Effect of male parents on the 100 kernel weight (g) and the number of kernels per ear of Bonanza, Ganebo and Arumba female parents |
 | |
| Different letters in a column within the same female indicate the significant differences at alpha 0,05 level according to the LSD-test | |
Mean separation of 100 kernel weight of F1’s between parents pairing and their open pollination of female parents was significantly differed only the crossing between Bonanza as female parent and Ganebo and Arumba varieties as male parents. While, crossing pairings on both Ganebo and Arumba varieties as female parents were not significantly differed in 100 kernels weight. Another crossing pairings between Bonanza and Ganebo varieties as male parents and Arumba variety as female parent was also the only pairings that indicated the significantly differed in kernel number per ear. Whereas, crossing pairings on Bonanza and Ganebo varieties as female parent, statistically was not significantly differed in producing kernels number of F1’s compared to their open pollinated of female parents (Table 3).
Metaxenia observation: One way analysis of variance on both ear diameter (mm) and length (cm) were significantly differed in all crossing pairings (Table 4). Comparison analysis in ear diameter and length between the open pollinated male parents and their F1’s resulted from crossing parents were significantly differed at alpha 0.05 level according to Tukey’s test. Crossing pairings on Bonanza variety as female parent and Ganebo variety as male parent and between Arumba variety as female parent and Bonanza and Arumba varieties as male parent were significantly differed on ear diameter. Ganebo variety as female parent was not affected by Bonanza and Arumba varieties as pollen parents in expressing the ear diameter. Crossing on Ganebo variety as female parents did not produce higher values of ear diameter and significantly differed when crossed with Bonanza and Arumba varieties as male parents, respectively. Another crossing pairing between Ganebo variety as female parent and Bonanza variety as male parent and between Arumba variety as female parent and Ganebo variety as male parent were significantly affected by their ear length compared to their open pollinated female parents.
| Table 4: | Variance analysis of the effect of male parent on ear diameter (mm) and ear length (cm) of Bonanza, Ganebo and Arumba female parents |
 | |
| *: Statistically significant differences at the 0.05 probability level | |
Whereas, crossing to the Bonanza variety as female parent was not significantly differed in yielding the ear length of their F1’s compared to the ear length of open pollinated female (Table 5).
DISCUSSION
Basically, there were four layers from outer to inner layer that composed the maize kernel, i.e., (1) Pericarp or outer layer, (2)Aleuron layer, (3)Endosperm layer and (4)Embryo. The kernel colour expression was due to the aleuron layer pigmentation, while the non-colour kernel expression was resulted from the lack of pigments produced by aleuron. Commonly, there are four genes that are responsible for the kernel colour expression asso called colour genes. Symbolicly, they are Pr1 gene that expressed the aleuron red colour, Y1 gene for white colour, C1 gene for coloured aleuron1 and R1 gene for coloured1. These two genes are similar but their name and symbols are different and encoded transcription activators that interact to influence the regulation of several structural genes within the anthocyanin pathway35.
In this field experiment, the kernels colour expression indicated the existency of xenia phenomen on pattern as listed in Table 1, particularly in crossing between yellow and white kernels in which the F1 ear or kernel of female parents produced the yellow and white colours in a proportionally comparable comparison.
| Table 5: | Effect of male parents on ear diameter (mm) and ear length of Bonanza-F1, Ganebo and Arumba female parents |
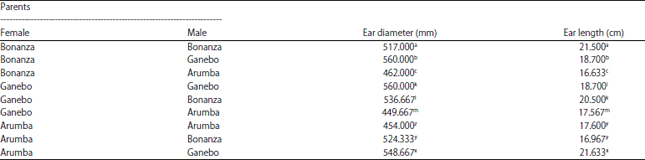 | |
| Different letters in a column within the same female indicate the significant differences at alpha 0,05 level according to the LSD-test | |
Unfortunately, the crossing between those parents that had the same colour kernels will be resulting an F1 which had the same kernels colour with their male and female parents which are naturally open pollinated. This findings was in line with Vancetovic et al.16 observation on top cross scheme of five commercial hybrid varieties (ZP-1, ZP-2, ZP-3, ZP-4 and ZP-5) with yellow colours as female parents and one open pollination variety with dark blue kernels colour as male parent. Vancetovic et al.16 reported that all the F1 ears (female ears, other than open pollinated) were similar to the male ears colour namely the dark blue colour. Another identical observation reported that the crossing between IHO variety with low carotene content and yellow colour kernels as female parent and PR variety with high anthocyanin content and dark purple colour kernels would result in the F1 kernels with which is similar to the female kernel colour i.e., dark purple colour kernels13.
Kernel colour expression in F1 female parent was the indication of xenia existency as a result of male parent pollen effect. By this pattern, another xenia elements like carbohydrates, proteins, lipids and other derivatives kernel contents, 100 kernel weight and number of kernels per ear, might easily be expected. Sprague36reported that the kernel colour inheritance pattern of maize showed that the red, yellow and purple colours were the result of four pairing factors, namely the respective symbols S1, S1, S2, s2, S3, S3, S4 and S4. Later, Ford35 explained that the fourth colours of maize kernel above were determined as alleles C1, C1-1, c1, Pr1, pr1, R1, r1 and Y1 coded by red, yellow and purple colours, respectively, whereas the y1 was the white colour kernel. The expressions of each maize kernel colour in F1 ear female was combination between their crossing pairing parents and the alleles number dependent35,36.
The weight of 100 kernels and the number of kernels per ear as another xenia elements generally indicated the decleaning on both variables of F1 female parent resulting from crossing between parents compared to the open pollination female parent. It means that the female parents plays an important role in the increasing the weight of 100 kernels and the number of kernel per ears. Only some crossing pairings which could be increased in these variables so that the compatibility of xenia phenomenon of both parents or the expression of xenia phenomenon in this observation was female parents dependent. From all pairing of parental crossings, there were two pairs of crossing which pointed the increasing of 100 kernels weight that are crossing between Bonanza female parent and Ganebo and Arumba varieties as male parents (Table 4). Meanwhile, the increase of kernels number per ear was expressed in the crossing between Arumba variety as female parent and Bonanza and Ganebo varieties as male parents. This expression showing the existence of paternal pollen effect of male plant in partial and the absence of maternal effect of female plant in remaining.
The important finding in this observation have shown that the parallele reports of another researchers12,13,16,37-39 in which either the 100 kernels weight or the kernels number per ear were reported the same pattern. This means that the inheritance of the 100 kernel weight and the kernel number per ear depended upon the heritability traits of crossing pairings between male and female parents. In this connection, the inheritance of some xenia element is strongly depended by pollen parent effect or male parent so called paternal effect. In contrast, the unexpressed xenia phenomenon may caused by the dominance female effect or maternal effect. So, the increasing of yield was indicated by the higher production of F1 female kernels derived from crossing between parents compared with the F1 female kernels by open pollination.
The ear diameter and length were metaxenia components that are closely related to the maize variety production. Ear length and diameter are two of some characters that are tightly associated in maize production and both of these characters are metaxenia elements. The bigger the ear diameter and the longer the ear length, the most probability of reaching the higher production of a maize variety as a result of the higher number of kernel per ear. As figured out at xenia phenomenon which showed only some crossing pairs that heritable, the metaxenia phenomenon in this findings was followed the above pattern. These could be seen in the F1 ear length and diameter obtained at the female parents in crossing pairing between male and female parents compared to the F1 open pollinated female parents (Table 5). Observation results from this field experiment showed the similar indication to the xenia effect that had been reported by several researchers from different regions12,13,16,37.
Both xenia and metaxenia in hesitancies are not always followed the increasing value of some characters but the decreasing value of some measures would be existed depending on the male parent potency of that character. In the case of the rising yield of some crop plant variety, the higher potential yield of male parent will be needed but in the case of lowering yield such as decreasing unsaturated lipid content of kernels, may substantially be needed the lower content of it component to the male parent. Then, the F1 kernels content obtained of female parent by crossing the male parent may be lower than the F1 kernels of female parent resulted from natural open pollination. Thus, the formation of the higher production of new superior maize variety through xenia and metaxenia exploitation, the higher production of male parent must be provided the higher values of 100 kernel weight, kernel number per ear and ear length and diameter.
CONCLUSION
The kernel colour as xenia element was fully inherited at crossing pairings between white and yellow colour kernels. Another xenia elements i.e. ,100 kernels weight was expressed when Ganebo and Arumba varieties were as male parents and the kernels number per ear was indicated when Bonanza and Ganebo varieties were as male parents. The ear diameter as metaxenia element was observed when Ganebo and Bonanza varieties as male parents. Whilst, the ear lengths as another metaxenia element was pointed out when Bonanza and Ganebo varieties were as male parents.
SIGNIFICANCE STATEMENT
This study reports results of a study on xenia and metaxenia on kernel and ear characteristics as production variables derived from hybrid maize varieties as parents. Results generated here will eventually help maize breeders use the xenia and metaxenia without lengthy procedure in forming the base population of maize as well as producing the new promising higher production of maize variety. The results presented will also give indication of appropriate use of both phenomena in generating the new superior varieties of maize.
ACKNOWLEDGMENT
We would like to extend our gratitude to the Ministry of Research, Technology and Higher Education of Republic of Indonesia for the financial support provided for this research project Contract No. 056/SP2H/LT/DRPM/2018. Also to Miss Nur Rizky Alfiany Suaib, English Language and Applied Linguistics, Faculty of Arts and Humanity, Coventry University, Priory Street, CV1 5FB, Coventry, United Kingdom, who provided proofreading for the English expressions.
REFERENCES
- Olfati, J.A., Z. Sheykhtaher, R. Qamgosar, A. Khasmakhi-Sabet, G. Peyvast, H. Samizadeh and B. Rabiei, 2009. Xenia and metaxenia on cucumber fruit and seed characteristics. Int. J. Vegetable Sci., 16: 243-252.
CrossRefDirect Link - Pixley, K.V. and M.S. Bjarnason, 1994. Pollen-parent effects on protein quality and endosperm modification of quality protein maize. Crop Sci., 34: 404-409.
CrossRefDirect Link - Letchworth, M.B. and R.J. Lambert, 1998. Pollen parent effects on oil, protein and starch concentration in maize kernels. Crop Sci., 38: 363-367.
CrossRefDirect Link - Bulant, C., A. Gallais, E. Matthys-Rochon and J.L. Priul, 2000. Xenia effects in maize with normal endosperm: II. Kernel growth and enzyme activities during grain filling. Crop Sci., 40: 182-189.
Direct Link - Ahuja, M. and N.S. Malhi, 2008. Genetic analysis of xenia effects in high oil maize lines. Indian J. Genet. Plant Breed., 68: 391-397.
Direct Link - Kahriman, F., C.O. Egesel and E. Sorlu, 2015. Effects of open- and self-pollination treatments on genetic estimations in maize diallel experiment. Spanish J. Agric. Sci., 13: 1-11.
CrossRefDirect Link - Kahriman, F., M. Serment, M. Haslak and M.S. Kang, 2017. Pollen effect (Xenia) for evaluating breeding materials in maize. J. Genetika, 49: 217-234.
CrossRefDirect Link - Thompson, P.R., A.B. Geyer, L.D. Lotz, H.J. Siegrist and T.L. Dobbels, 2002. Top cross high-oil corn production: Agronomic performance. Agron. J., 94: 290-299.
Direct Link - Thomison, P.R., A.B. Geyer, L.D. Lotz, H.J. Siegrist and T.L. Dobbels, 2003. Top cross high oil corn production: Select grain quality attributes. Agron. J., 95: 147-154.
Direct Link - Vancetovic, J., S. Zilic, S. Bozinovic and D. Ignjatovic-Micic, 2014. Simulating of top-cross system for enhancement of antioxidants in maize grain. Spanish J. Agric. Res., 2: 467-476.
CrossRefDirect Link - Swingle, W.T., 1928. Metaxenia in the date palm: Possibly a hormone action by the embryo or endosperm. J. Hered., 19: 257-268.
CrossRefDirect Link - Nixon, R.W., 1928. Immediate influence of pollen: In determining the size and time of ripening of the fruit of the date palm. J. Heredity, 19: 241-255.
CrossRefDirect Link - Duc, G., A. Moessner, F. Moussy and C. Mousset-Declas, 2001. A xenia effect on number and volume of cotyledon cells and on seed weight in faba bean (Vicia faba L.). Euphytica, 117: 169-174.
CrossRefDirect Link - Weingartner, U., O. Kaeser, M. Long and P. Stamp, 2002. Combining cytoplasmic male sterility and xenia increases grain yield of maize hybrids. Crop Sci., 42: 1848-1856.
Direct Link - Weingartner, U., T.J. Prest, K.H. Camp and P. Stamp, 2002. The plus-hybrid system: A method to increase grain yield by combined cytoplasmic male sterility and xenia. Maydica, 47: 127-134.
Direct Link - Weingartner, U., K.H. Camp and P. Stamp, 2004. Impact of male sterility and xenia on grain quality traits of maize. Eur. J. Agron., 21: 239-247.
CrossRefDirect Link - Mizrahi, Y., J. Mouyal, A. Nerd and Y. Sitrit, 2004. Metaxenia in the Vine Cacti Hylocereus polyrhizus and Selenicereus spp. Ann. Bot., 93: 469-472.
CrossRefDirect Link - Dag, A. and Y. Mizrahi, 2005. Effect of pollination method on fruit set and fruit characteristics in the vine cactus Selenicereus megalanthus ("yellow pitaya"). J. Hortic. Sci. Biotechnol., 80: 618-622.
CrossRefDirect Link - Pahlavani, M.H. and K. Abolhasani, 2006. Xenia effect on seed and embryo size in cotton (Gossypium hirsutum L.). J. Applied Genet., 47: 331-335.
CrossRefDirect Link - Piotto, F.A., K.D. Batagin-Piotto, M.D. Almeida and G.C.X. Oliveira, 2013. Interspecific xenia and metaxenia in seeds and fruits of tomato. Scient. Agricola, 70: 102-107.
CrossRefDirect Link - Allard, R.W., 1960. Principles of Plant Breeding. 1st Edn., John Wiley and Sons Inc., New York pp: 372-372.
CrossRefDirect Link - Griffing, B., 1956. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci., 9: 463-493.
Direct Link - Ford, R.H., 2000. Inheritance of kernel color in corn: Explanations & investigations. Am. Biol. Teach., 62: 181-188.
CrossRefDirect Link - Sprague, G.F., 1932. The inheritance of colored scutellums in maize. Techn. Bull., 292: 1-44.
Direct Link - Weiland, R.T., 1992. Cross-pollination effects on maize (Zea mays L.) hybrid yields. Can. J. Plant Sci., 72: 27-33.
CrossRefDirect Link - Alvarez-Prado, S., C.G. Lopez, M.L. Senior and L. Borras, 2014. The genetic architecture of maize (Zea mays L.) kernel weight determination. G3: Genes Genet. Genomes, 4: 1611-1621.
CrossRefPubMedDirect Link - Liu, Z., A. Garcia, M.D. McMullen and S.A. Flint-Garcia, 2016. Genetic analysis of kernel traits in maize-teosinte introgression populations. G3: Genes Genomes Genet., 6: 2523-2530.
CrossRefDirect Link